|
|
 |
|
Calanoida ( Ordre ) |
|
|
|
Arietelloidea ( Superfamille ) |
|
|
| |
| | | |
| Heterorhabdidae Sars, 1902 ( Arietelloidea ) | | Syn.: | Heterochätina Giesbrecht, 1892 (p.63);
Heterochaetidae Sars,1900 (p.79);
Heterorhabdinae : Esterly, 1905 (p.181) | | Ref.: | Sars, 1902 (1903) (p.73, 117); Gurney, 1931 a (p.84); Rose, 1933 a (p.198); Brodsky, 1950 (1967) (p.82, 341); Mazza, 1967 (p.190); Heptner, 1971 (p.146); 1972 a (p.54, Rev., Genera Key.); Grice, 1973 (p.943, Rem.); Andronov, 1974 a (p.1005); Razouls, 1982 (p.460); Bowman & Abele, 1982 (p.9); Brodsky & al., 1983 (p.143, 145); Zheng Zhong & al., 1984 (1989) (p.248); Mauchline, 1988 (p.708: cuticular pores); Huys & Boxshall, 1991 (p.463); Razouls, 1993 (p.307); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.816); Ohtsuka & al., 1997 (p.577, Phylogeny); Bradford-Grieve & al., 1999 (p.883, 902, 903, 942, 943: Key of Genera); Bradford-Grieve,1999 b (p.70, Def., Rem.); Park, 2000 (p.5, Rem., p.9: Genera & Sug-Genera Keys); Ohtsuka & Huys, 2001 (p.461); Boxshall & Halsey, 2004 (p.14; 49; 124: Def.; p.126: Geneta Key); Vives & Shmeleva, 2007 (p.292, part. Genera Key); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig. 2, 3, 4, molecular biology, phylogeny); Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | La famille des Heterorhabdidae est revue par Heptner (1972) qui estime artificiel les caractères utilisés, notamment pour les genres Hemirhabdus et Disseta. Les espèces sont regroupées sur la base de la structure des appendices buccaux liée au type de régime alimentaire qui évolue du mode filtreur (Disseta) au mode carnivore suceur (Heterorhabdus). L'auteur crée alors les deux genres nouveaux: Neorhabdus et Microdisseta. Un examen comparé des Md, Mx1, Mx2 et Mxp conduit l'auteur à envisager une évolution dans la famille. Le genre Heterorhabdus est divisé en deux sous-genres par Brodsky (1950): Paraheterorhabdus (type: H. robustus) et Euheterorhabdus (types: H. papilliger et H. norvegicus).
Grice (1973) ajoute à cette famille le nouveau genre Alrhabdus à partir de la découverte d'une femelle (A. johrdae), qui présente des affinités à la fois avec les Heterorhabdidae et les Augaptilidae, mais l'auteur, faute de la connaissance du mâle, ne crée pas de famille particulière. Il suggère que diverses espèces de calanoïdes hyperbenthiques sont dans une situation semblable. Park (2000, p.1, 8, 141) exclut ce genre de la famille des Heterorhabdidae de même que Microdisseta, maintenus ici pour mémoire, et confirme le genre Paraheterorhabdus de Brodsky (1950)
8 G: Microdisseta, Disseta, Hemirhabdus, Heterorhabdus, Heterostylites, Mesorhabdus, Neorhabdus, Paraheterorhabdus.
Les tailles sont les plus grandes dans les trois genres Hemirhabdus, Neorhabdus et Disseta, respectivement, suivi parMesorhabdus, Heterostylites, Paraheterorhabdus (Paraheterorhabdus), Heterorghabdus, respectivement, enfin avec les dimensions qui les distinguent nettement des genres précédents, Paraheterorahbdus (Antirhabdus) et surtout Microdisseta dont les longueurs corporelles sont inférieures au millimètre (± 0,740).
After Madhupratap & al. (1996), la famille des Heterorhabdidae représente de 0,1 à 0,3 % des copepodes selon la saison dans la couche de mélange des eaux océaniques de la région ouest de l'Inde (Mer Arabe), en usant un filet type Multiple Plankton Closing Net à 200 µm de vide de maille (mesh aperture). | 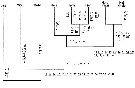 issued from : S. Ohtsuka, H.Y. Soh & S. Nishida in J. Crustacean Biol., 1997, 17 (4). [p.589, Fig.8]. Cladogramm showing phylogenetic relationships between genera of the family Heterorhabdidae generated with PAUP when all characters are set unordered. The numbers corresopnd to characters in Table 3 (following figure). Underline = multistate character; * = reversal; cross = convergence. Diss = Disseta; Micr = Microdisseta; Meso = Mesorhabdus; Hets = Heterostylites; Hemi = Hemirhabdus; Neo = Neorhabdus; Hete (Para) = Heterorhabdus (Paraheterorhabdus); Hete (Hete) = Heterorhabdus (Heterorhabdus). |
 issued from : S. Ohtsuka, H.Y. Soh & S. Nishida in J. Crustacean Biol., 1997, 17 (4). [p.587]. Characters used in the cladistic analysis (in the matrix) for genera on the family Heterorhabdidae. Codes 0-4 refer to transformation series of multistate characters. o = plesiomorphic state, 1 = advanced (apomorphic) state, 2-4 = further advanced states, 9 = missing data. |
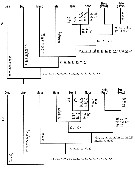 issued from : S. Ohtsuka, H.Y. Soh & S. Nishida in J. Crustacean Biol., 1997, 17 (4). [p.590, Fig.9]. Two cladograms showing phylogenic relationships between genera of the family Heterorhabdidae generated with PAUP when all characters are set-ordered. The numbers, symbols, and abbreviated generic names as in Fig. 8 and Table 3 (above). For the authors gut content analyses partly support Heptner's (1972) conclusion on the feeding habits of the family. The phylogenetic relationships among the genera of the family generated in the present study are almost identical with the noncladistic evolutionary tree of Hepner (1972), which was not based on detailed observations of the mandibular ventralmost teeth and the posterior surface of the labrum. The particle-feeder Disseta is the first to diverge from the main lineage, while the carnivores Hemirhabdus-Neorhabdus-Heterorhabdus are the terminal branch. Microdisseta is the second off-shoot in the evolution of the family. Mesorhabdus and Heterostylites are intermediate between these suspension feeders and the carnivores. The relative position of Heterostylites in the cladograms is equivocal. The specialization of the mandibular ventralmost tooth and the supposed paralytic-substance-releasing gland openings on the labrum are shared by Heterorhabdus and the Hemirhabdus-Neorhabdus clade. The evolutionnary transformation from particle feeders to pure carnivores has been accomplished by a combination of reductions or losses and specializations of several elements on the mouthpart appendages and the conversion of a lateral pair of labral secretory glands into a venom- or anaesthetic-realising system. Only in Heterorhabdus the relative elongation of the maxillary (Mx2) praecoxa is added to these transformations (length of Mx2 praecoxa in relation to body length female = 19.7). The elongation supposedly guarantees effective capture of prey animals (see in Ohtsuka, 1991). The remarkable elongation of the Mx2 praecoxa is found separately in Heterorhabdus and Heterostylites (ratio length of Mx2 praecoxa in relation to body length female = 14.4 in the latter) is not involved in the cladistic analysis. The lateral pair of large gland openings (type-1) on the posterior surface of labrum of Hemirhabdus-Neorhabdus-Heterorhabdus seem not be novel structures, but a conversion of two outer pores of two pairs of openings of ancestral type-3 glands into type-1 gland openings. In Disseta, Mesorhabdus, and Heterostylites, which lack type-1 glands, there are two pairs of openings of type-3 glands of almost equal size, while in Hemirhabdus, Neorhabdus, and Heterorhabdus, which possess type-1 glands, there is only one pair of openings of type-3 glands. This interpretation is supported by the fact that both type-1 and type-3 glands have one-paired gland cells and a common opening (see in Nishida & Ohtsuka, 1996); the conversion may be accompanied by an increase in the size of the gland system and the lateral movement of the position of the openings. In the Md, the reductions or losses of dorsal teeth and of daggerlike spinules have occurred throughout the evolution of the family. Heterostylites and Heterorhabdus may have independently developped the most advanced condition, the complete absence of these elements. The specialization of the ventralmost tooth and its isolation from the remaining teeth are evidently associated with the acquisition of the type-1 glands, and facilitate its protrusion from between the labrum and paragnath to inject a paralytic substance into prey (see in Nishida & Ohtsuka, 1996). The complete tubular structure of the ventralmost tooth of Heterorhabdus seems to be much effective in injecting a paralytic substance into prey than the semitube of Hemirhabdus and the groove of Neorhabdus. To avoid leaking the labral paralytic substance, the postrior side of the labrum presumably functions as an operculum to close the groove of Neorhabdus and the semitube of Hemirhabdus during injection of the substance into prey via the ventralmost mandibular tooth. The central teeth also appear to have become relatively elongate and slender in an evolutionary spep from Disseta to Mesorhabdus. In the Mx2, the transformations in the family can be explained by both developments and reductions of elements. Two setae on the basis and the first endopod segment are highly specialized into a grasping organ in the carnivores, with heavy sclerotization of the setae with hooked tips and an inner row of stout spinules. Reductions of setae on the second to fourth endopod segments are exhibited by all carnivores. Similar specializations and reductions of the Mx2 elements have convergently occurred also in the deep-sea predatory family Hyperbionychidae (see in Ohtsuka & al., 1993). |
 Issued from : E.T. Park & F.D. Ferrari in A selection from Smithsonian at the Poles Contributions to International Polar year. I. Krupnik, M.A. Lang and S.E. Miller, eds., Publs. by Smithsonian Institution Scholarly Press, Washington DC., 2009. [p.163, Table 8] Endemic species of several species of Heterorhabdidae found in four different areas of interest. A''+'' indicates presence. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.124]. Armature formula of swimming legs P1 to P5 Nota: Female P5 relatively unmodified, with 3-segmented rami; inner seta on 2nd exopodal segment strongly developed and directed medially. - Male P5 asymmetrical; biramous, typically with serrate coxa and basis, and 3-segmented rami; right leg typically with large medial process on basis. 1st exopodal segment with outer spine, 2nd with rounded or spinous inner process plus distal spine, 3rd typically forming curved claw armed with inner seta proximally and long subapical seta. Left leg with or without small medial process on basis; endopod formula 0-0; 0-1; 2,2,2, or with seta on middle segment absent. 1st and 2nd exopodal segmets each with outer spine, 3rd segment drawn out into apical claw, armed with inner and outer spines proximal to base of claw. - Eggs release into water. |
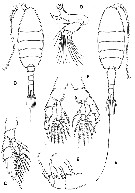 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.125, Fig. 25]. Heterorhabdidae. A, heterorhabdus robustus habitus female; B, Md: C, P5; D, habitus male; E, Hemirhabdus latus (as Macrorhabdus latus) female Mx2; F, Mesorhabdus gracilis male P5. [Sars, 1924]. |
 Issued from J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.71]. Spine and seta formula of swimming legs P1 to P5. - P5 female natatory, symmetrical, similar to other swimming legs but smaller; inner edge seta on exopod segment 2 ismodified into a long slender spine articulated with its segment; exopod segment 2 posterior distal border expanded into a toothed flange. - P5 male with a 3-segmented endopod and exopod on left and right; right basipod 2 usually with inner extensions of variable shape and usually fringed by hairs or spinules, sometimes with an inner extension on the left; right exopod segment 2 with an inner expansion of variable shape but usually in the form of a thick spine; left exopod segment 3 often but not always termunated by an elongate spine. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9]. Key of genera and subgenera of the family Heterorhabdidae. |
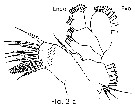 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 154, Fig.2 a]. Mx1 of Disseta palumbii female. |
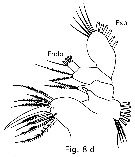 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 160, Fig.8 d]. Mx1 of Mesorhabdus angustus female. |
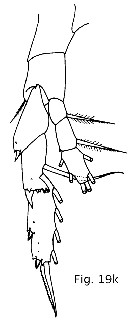 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 &171, Fig.19 k]. P5 of Heterostylites major female. |
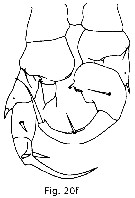 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 172, Fig. 20 f]. P5 (with endopods omitted, posterior view) of Heterostylites major male. Nota: female as 19 k. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 183, Fig.31 b]. Mx2 of Hemirhabdus grimaldii female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 187, Fig.35 b]. Left Mx2 (asp: anterior spine; psp = posterior spine; tsp = terminal spine) of Neorhabdus latus female. |
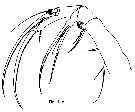 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 196, Fig.44 d]. Mx2 of Paraheterorhabdus (Paraheterorhabdus) robustus female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 196, Fig. 44 a, b]. Md of Paraheterorhabdus (Paraheterorhabdus) robustus female. a: left mandible (posterior view); b: masticatory edge of right mandible (posterior view). |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 209, Fig.57 a, b]. Fig. 57 a, b: Md of Paraheterorhabdus (Antirhabdus) compactus female. a: left mandible (posterior view); b, masticatory edge of right mandible (posterior view). |
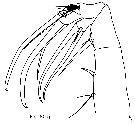 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 212, Fig.60 g]. Left Mx2 (posterior view) of Heterorhabdus spinifrons female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.268, Fig.116]. Morphological characters states in the genera Heterorhabdidae. |
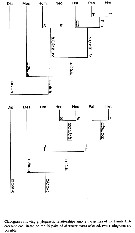 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.269, Fig.116 ]. Phylogenetic relationships among the genera of Heterorhabdidae. | | | | | Alrhabdus Grice, 1973 | | Ref.: | Grice, 1973 (p.943, 946); Razouls, 1982 (p.476); 1993 (p.307); Bradford-Grieve,1999 b (p.71, Déf.); Park, 2000 (p.1, 8, 141) | | Rem.: | Boxshall & Halsey (2004 (p.68, 126) transfèrent ce genre parmi les Augaptilidae, cependant avec quelque doute tant que le mâle n'ait pas été trouvé. | | | | (1) Disseta Giesbrecht, 1889 | |
| | Ref.: | Giesbrecht, 1892 (p.63, 369); Giesbrecht & Schmeil, 1898 (p.112); Esterly, 1906 a (p.71); A. Scott, 1909 (p.133); Wolfenden, 1911 (p.313); Sars, 1925 (p.221); Rose, 1929 (p.34, Rem.); Sewell, 1932 (p.308); Rose, 1933 a (p.198); Farran, 1948 d (n°15, p.3); Brodsky, 1950 (1967) (p.342, clé spp.); Tanaka, 1964 (p.32); Heptner, 1972 a (p.56, spp. Key); Razouls, 1982 (p.461); Mauchline, 1988 (p.706); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.816); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve,1999 b (p.71, Def.); Park, 2000 (p.11, Rem., spp. key); Boxshall & Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.293) | | Rem.: | type: Disseta palumbii. 3 spp.: | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 8.295 mm (n = 6; SD = 1.7147) and the mean male size is 7.698 mm (n = 6; SD = 1.7321). The size ratio (Male : Female) is 0.929 (n = 3; SD = 0.0617). The sex ratio (Female : Male) is 1. |  Diagnosis of Disseta: 1 - Ventralmost blade on mandibular gnathobase only slightly thickened, separated from adjacent teeth by narrow gap, no wider than base of blade 2 - Distal coxal endite of Mx2 with 3 unmodified setae. 3 - Basal and 1st endopodal endite of Mx2 without large, claw-like elements. Coxal endite of Mx1 with at least 2 setae. 4 - Basal endite of Mx2 without claw-like element, armed with 3 setae. |
 Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.75, Fig.42]. Disseta palumbii. Female (from Bradford-Grieve, 1999): A, urosome (dorsal view); B, same (lateral view); C, Md; D, P5, E, spine from inner distal corner of exopod segment 2 of P5. Male: F, P5. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.155, Fig.3 d-k]. Disseta palumbii male: d, right leg of P5 with 2 distal exopodal segments omitted (anterior view); e, left leg of P5 with2 distal exopodal segments omitted (anterior view); f, basipod and 1st exopodal segment of left leg of P5 (posterior view); g, P5 (posterior view)h, exopod of left leg of P5 (posterior view); i, same (anterior view); j, exopod of right leg of P5 (posterior, tilted clockwise); k, same (posterior view). mpr = medial projection. |
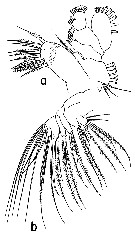 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.154, Fig.2 a, b]. Disseta palumbii female: a, left Mx1 (posterior view); b, left Mx2 (posterior view). Nota: - Mx1 with on t1st inner lobe (= arthrite) 3 small anterior submarginal, 9 marginal and 2 posterior submarginal spiniform setae; 2nd inner lobe (= coxal endite) with a single seta; 3rd lobe (= basal endite) with 3 setae of similar size; basis well developed with 4 setae; in endopod, 1st and 2nd fused segments with 3 and 4 setae, respectively, 3rd segment with 5 setae; exopod relatively large, extending to distal end of endopod, with 6 large setae along outer margin and 5 small setae distally. - Mx2 with 1st lobe with 2 proximal, 1 posterior subterminal, and 3 terminal setae in addition to a short spiniform process; 2nd to 4th lobes each with 1 posterior subterminal and 2 terminal setae; 5th lobe with 1 posterior subterminal, 2 terminal setae and 1 saberlike spine; 6th lobe with 1 posterior subterminal and 1 terminal setae and 1 saberlike spine. Saberlike spines of 5th and 6th lobes are finely serrated, only a little thicker but much shorter than principal terminal setae. Endopod with 4 separate segments.Of 7 endopodal setae, a posterior seta of 1st and 2nd sdegments much shorter than the remaining 5; only posterior setae of first 2 endopodal segments conspicuously armed with setules. | | | | | (2) Hemirhabdus Wolfenden, 1911 | |
| | Syn.: | Macrorhabdus Sars, 1920 c (p.11) | | Ref.: | Wolfenden, 1911 (p.308, Def.); Sars, 1925 (p.230); Sewell, 1932 (p.302, Rem.); Rose, 1933 a (p.205); Farran, 1948 d (n°15, p.3); Tanaka, 1964 a (p.26); Heptner, 1972 a (p.58, Rem.); Razouls, 1982 (p.477); Mauchline, 1988 (p.708); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.820); Mauchline, 1998 (p.70 : F; p.74: M); Bradford-Grieve,1999 b (p.76, Def.); Park, 2000 (p.51, 52: Key spp., Rem.: p.143); Boxshall & Halsey, 2004 (p.126); Vives & Shmeleva, 2007 (p.295) | | Rem.: | type: Heterochaeta grimaldii Richard, 1893 = Hemirhabdus grimaldii. Total: 2 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 10.233 mm (n = 4; SD = 2.0230) and the mean male size is 10.585 mm (n = 4; SD = 1.6406). The size ratio (Male : Female) is about 1 . Le sex ratio (Female : Male) is 1. | 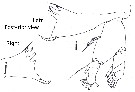 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal., San Diego, 2000, 31 [p.182, Fig. 30 i-j] Hemirhabdus grimaldii female: i, left Md (posterior view); j, masticatory edge of right Md (posterior view). |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal., San Diego, 2000, 31 [p.184, Fig. 32 d-j] Hemirhabdus grimaldii Male: d, P5 (anterior view); e, same (posterior view; right endopod omitted); f, exopod of right leg of P5 (posterior view); g, distal end of exopod of left leg of P5 (anterior view); h, 2nd exopodal segment of right leg of P5 (posterior view); i, same (anterior view); j, same (tilted clockwise). | | | | | (3) Heterorhabdus Giesbrecht, 1898 | |
| | Syn.: | Heterochaeta Claus, 1863 (p.180); Dahl, 1894 (p.77, Key spp.); Sars, 1900 (p.79); Heterochäta : Giesbrecht, 1892 (p.64, 372); Alloiorhabdus (part.) Wolfenden, 1911 (p.303); Vervoort, 1951 (p.140, Rem.)
Nota: Heterochaeta preoccuped name, therefore renamed Heterorhabdus by Giesbrecht & Schmeil (1898). | | Ref.: | Giesbrecht & Schmeil, 1898 (p.113, spp. Key); Sars, 1902 (1903) (p.117); Esterly, 1905 (p.182, clé spp.); van Breemen, 1908 a (p.117, spp. Key); A. Scott, 1909 (p.130); Wolfenden, 1911 (p.302); Sewell, 1932 (p.298); Wilson, 1932 a (p.131, spp. Key); Rose, 1933 a (p.199, spp. Key); Mori, 1937 (1964) (p.73); Sewell, 1947 (p.178, Rem.); Farran, 1948 e (n°16, p.3); Davis, 1949 (p.56); Brodsky, 1950 (1967) (p.344, spp. Key); Tanaka, 1964 a (p.1); Bradford, 1971 a (p.120, 135, spp. Key); Heptner, 1972 a (p.56, 59, spp. Key); Razouls,1982 (p.464); Gardner & Szabo, 1982 (p.363); Mauchline, 1988 (p.706); Zheng Zhong & al., 1984 (1989) (p.248); Chihara & Murano, 1997 (p.817); Mauchline, 1998 (p.70: F; p.74: M); Barthélémy, 1999 a (p.29); Bradford-Grieve & al., 1999 (p.943, 944: spp. Key); Bradford-Grieve,1999 b (p.76, Def.); Park, 2000 (p.89, Redef., Key of 4 Groups, Rem.: p.144); Boxshall & Halsey, 2004 (p.126); Vives & Shmeleva, 2007 (p.297, spp. key) | | Rem.: | type: Heterorhabdus spinifrons (Claus, 1863).
Brodsky , 1950 (1967, p.345) établit les 2 sous-genres : Heterorhabdus (type: H. robustus) et Euheterorhabdus (types : H. papilliger & H. norvegicus.
Park (2000, p.90) établit 4 groupes ("spinifrons", "papilliger", "fistulosus", "abyssalis").
Total: 31 spp. (+ 4 douteuses): | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 3.249 mm (n = 62; SD = 2.6382) and the mean male size is 2.792 mm (n = 62; SD = 0.6757). The size ratio (Male : Female) is 0.859 or if we consider the mean size for each species, we obtain 0.959 (n = 31; SD = 0.0512). The sex ratio est 1.
For the first group (''spinifrons''): the mean female size is 2.881 mm (n = 14; SD = 6860) and the mean male size is 2.719 mm (n = 14; SD = 0.6035). The size ratio (male : female) is 0.944. The sex ratio is 1.
For the second group (''papilliger''): the mean female size is 2.084 mm (n = 10; SD = 0.3832) and the mean male size is 2.047 mm (n = 10; SD = 0.3970). The size ratio (male : female) is 0.982. The sex ratio is 1.
For the third group (''fistulosus''): the mean female size is 3.278 mm (n = 4; SD = 0.6592) and the mean male size is 3.308 mm (n = 4; SD = 0.7100). The size ratio (male : female) is 1.009. The sex ratio is 1.
For the fourth group (''abyssalis''): the mean female size is 3.152 mm (n = 34; SD = 0.7383) and the mean male size is 2.980 mm (n = 34; SD = 0.6095).The size ratio (male : female) is 0.946. The sex ratio is 1.
| Diagnosis of Heterorhabdus: 1 - Ventralmost blade on mandibular gnathobase thickened and curved, separated from remaining distal teeth by marked diastema. 2 - Mx2 with 2 setal elements on basis and on distal endite of coxa; proximal praecoxal endite with 2 setae. Middle endite of syncoxa of Mxp typically represented by long, distally-curved seta. 3 - Mx2 with 2 large, claw-like setal elements on basis. |
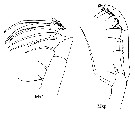
 1 a - Mxp: Middle coxal spine of Mxp smoothly curved (cf. H. spinifrons Fig.60-h). Mx2: None of the spines on 5th lobe conspicuously serrated with long spinules (cf. H. spinifrons Fig.60-g)....... spinifrons species Group.
1 b - Mxp: Middle coxa spine distinctly bent ( H. guineanensis Fig.73-a). Mx2: 1 of the spines on 5th lobe conspicuously serrated with long spinules (cf. H. guineanensis Fig.72-i)...... 2.
2 a - Mx2: Unserrated spine on 5th lobe much shorter than serrated spine (cf. H. H. hadrosomus Fig.84-i). Basal lobe of male right P5 relatively long and arising from middle of segment (cf. H. hadrosomus Fig.85-g)...... abyssalis species Group.
2 b - Mx2: Unserrated spine on 5th lobe nearly as long as or only a little shorter than serrated spine (cf. H. guineanensis Fig.72-i. Basal lobe of male right P5 relatively short and arising close to distal end segment. (cf. H. guineanensis Fig.73-c).......3 a, b.
3 a: Dorsally midanterior tubercular process of forehead somewhat truncate (cf.
H. egregius Fig.82-d). Mxp: Basis with large elevated pore on medial margin close to distal end (cf.H. egregius Fig.82-h)...... fistulosus species Group.
3 b: Dorsally, midanterior tubercular process of forehead pointed (cf. H. guineanensis Fig.72-d). Mxp: Basis without elevated pores on medial margin close to distal end......papilliger species Group. |
 Issued from T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31. [p.212, Fig.60, g-h]. Appendage types to identification for spinifrons species Group. |
 Issued from T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31. [p.224, Fig.72-i; p.225, Fig.-73-a]. Appendage types to identification for other species Group ( abyssalis'', ''papilliger'', ''fistulosus''). |
 Issued from T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31. [p.212, Fig.84-i, p.237; Fig.85-g]. Appendage types to identification for abyssalis species Group. |
 Issued from T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31. [p.224, Fig.72-i; p.225, Fig.73-g ]. Appendage types to identification for ofher species Group ( ''papilliger" and ''fistulosus"). |
 Issued from T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31. [p.224, Fig.72-d; p.225, Fig.73-a]. Appendage types to identification for papilliger species Group. |
 Issued from T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31. [p.234, Fig.82-d, h-k]. Appendage types to identification for fistulosus species Group. |
 Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.87, Fig.55]. Heterorhabdus spinifrons. Female (from Bradford-Grieve, 1999): A, genital segment (dorsal view); B, same (lateral view); C, forehead (dorsal view); D, same (lateral view); E, Mx2 lobe 4; F, P5. Male: G, forehead (dorsal view); H, same (lateral view); I, P5. |
 - - Midanterior tubercular process of forehead angular or producee into a spiniform process. - Small posterior subterminal spine of 4th lobe of Mx2 about 1/4 length of long, saberlike spines; large falciform spine of 5th lobe not conspicuously serrated with spinules and small saberlike spine far short of reaching distal end of falciform spine; endopodal setae relatively poorly developed. - In Mxp, middle marginal spine saberlike and smoothly curved, reaching distal end of basis at most. - In male P5, inner lobe of right basis arising from distal part of medial margin of segment, relatively short and pointing in a distomedial direction; medial projection of 2nd exopodal segment of right P5 large, arising along proximal margin of segment, terminated with a bilobed structure consisting of a rounded plumose proximal lobe and a naked or partially plumose distal lobe, and a small spiniform process in between. - 7 species are referred to this group: H. spinifronsH. subspinifronsH. caribbeanensisH. heterolobus H. quadrilobusH. ankylocolus H. insukae. After Park (2000, p.144), H. caribbeanensis, H. insukae are epipelagic and the rest of the species group are epi-mesopelagic. H. spinifrons, H. subspinifrons, H. caribbeanensis, H. insukae are all common circumglobal species occurring widely in the low and midlatitudes of the world's oceans. The most common of these 4 species was H. spinifrons followed by H. insukae. They accounted for 28 % and 26 %, respectively of the specimens found for the species group, and H. subspinifrons and H. caribbeanensis for 11 % and 6 %, respectively. H. ankylocolus representing only 6 % of the species group, is an Indo-Pacific equatorial species. H. heterolobus representing 7 % of the species group, was found exclusively in the equatorial waters of the whole Pacific. H. quadrilobus is endemic to the eastern Pacific, where it is distributed mainly along the coasts and extends westward for some distance in equatorial waters, which account for 16 % of the species group. |
 - - Midanterior tubercular process of forehead angular or producede into a spiniform process. - Small subterminal posteriorb spine of 4th lobe of Mx2 longer than 1/2 length of large, saberlike spines of same lobe; large falcate spine of 5th lobe conspicuously serrated with spinules and small saberlike spine reaching close to distal end of falcate spine; endopodal setae relatively poorly developed. - In Mxp, middle marginal spine falcate, distinctly bent at a point 2/7 length of spine from proximal end and extending beyond distal end of basis. - In male P5, inner lobe of right basis arising from distal end of medial margin, shorter than segment and extending in a distomedial direction; medial projection of 2nd exopodal segment of right P5 large, arising along proximal margin of segment, terminated with a bilobed structure consisting of a rounded plumose proximal lobe, a naked or partially plumose distal lobe, and a small terminal spiniform process in between. 5 species are referred to this species group: H. papilligerH. spiniferH. lobatusH. guineaenensisH. prolatus. For Park (2000, p.144), the papilliger species group seems to be epi-mesopelagic. H. papilliger and H. spinifer are circumglobal species occurring throughout the low and midlatitudes of the world's oceans. H. prolatus is an Indo-Pacific equatorial species, and H. guineanensis found exclusively in warm waters of the eastern Atlantic. H. lobatus has a rather unusual distribution, occurring along the southern coast of South Africa, around New Zealand, and along the west coast of the Americas from 40° S to 40° N. The most common species was H. papilliger, representing 51 % of specimens found for the species group, followed by H. lobatus accounting for 29 %. |
 - - Midanterior tubercular process of forehead low and somewhat truncated. - 3rd marginal seta of left caudal ramus and 3rd and 4th marginal setae of right ramus armed with spines in addition to normal setules. - Subterminal posterior spine of 4th lobe of Mx2 longer than 1/2 length of large, saberlike spines of same lobe; falcate spine of 5th lobe conspicuously serrated with spinules and saberlike spine reaching close to distal end of falcate spine; endopodal setae relatively well developed. - In Mxp, middle marginal spine of coxa falcate, strongly elbowed 1/4 length of spine from proximal end and extending beyond distal end of basis; basis with 1 or 2 large, elevated pores distal to 3rd middle marginal seta. - In male P5, inner lobe of right basis arising from distomedial corner of segment, shorter than segment and extending in a distomedial direction; medial projection of 2nd exopodal segment of right P5 about equal to or shorter than 1/2 length of segment, nearly conical, its proximal margin some distance from proximal end of segment. 2 species are referred to this species group: H. fistulosusH. egregius. Remarks: This species group is intermediate between the papilliger species and abyssalis species groups. Another character is that Mx2 is intermediate with regard to the length of the saberlike spine of its 5th lobe. For Park (2000, p.144), H. egregius is a circumglobal species occurring widely in the low and midlatitudes of the world's oceans and H. fistulosus seems to be endemic to the northern Pacific. |
 - - Midanterior ubercular process of forehead low and more or less truncated. - 3rd marginal seta of left caudal ramus and 3rd and 4th marginal setae of right ramus armed with spines in addition to normal setules. - Posterior subterminal spine of 4th lobe of Mx2 longer than 1/2 length of large, saberlike spines of same lobe; falcate spine of 5th lobe conspicuously serrated with spinules, saberlike spine about equal to or less than 3/4 length falcate spine; endopodal setae relatively well developed. - In Mxp, middle margibal spine of coxa falcate, distinctly bent 1/4 length of spine from proximal end and extending far beyond distal end of basis. - In male P5, inner lobe of right basis arising from middle or proximal half of segment, longer than segment and extending in a distolateral direction; medial projection of 2nd exopodal segment of right P5 relatively small, about equal to or shorter than 1/3 length of segment. 17 species are referred to this species group. H. norvegicusH. abyssalisH. clausiiH. tanneriH. austrinusH. pustuliferH. pacificusH. spinosus H. hadrosomusH. prokixusH. tuberculusH. oikoumenikisH. longisegmentusH. americanusH. cohibilisH. paraspinosusH. confusibilis. Remarks: The females of this species group are similar to those of the fistulosus species group but distinguished from them by a relatively short saberlike spine of the 5th lobe of Mx2 and the absence of extraordinarily large elevated pores on the basis of Mxp. The males are different from those of all other species groups of the genus in having a greatly elongated inner basal lobe of the right P5. For Park (2000, p.144) The species in this species group seems to be meso-bathypelagic. H. hadrosomus, H. oikoumenikis, H. pacificus, H. confusibilis are circumglobal species occurring throughout the low and midlatitudes of the world's oceans; of these, H. pacificus was the most common, accounting for 14 % of the specimens found for the group, followed by H. oikoumenikis accounting for 10 %. H. longisegmentus is also circumglobal but occurred only in the low and midlatitudes of the Southern Hemisphere and it was represented in the samples by 5 % of the group.
Only 1 species, H. tuberculus, occurred widely in the low and midlatitudes of the whole Indo-Pacific, and it was represented by 11 % of the group.
6 species: H. cohibilis, H. clausii, H. tanneri, H. abyssalis, H. americanus, H. prolixus were found only in the Pacific. H. cohibilis, H. clausii are Pacific equatorial species, the former occurring in the whole equatorial waters and the latter only in the central and eastern equatorial waters. H. tanneri is endemic to the coastal waters of the northern Pacific. H. abyssalis, H. americanus, H. prolixus are confined in the eastern Pacific, where they occur mainly in the coastal waters of the low latitudes. The most common of the Pacific species was H. tanneri, representing 15 % of the group, followed in order by H. abyssalis, H. prolixus, which were represented approximately by the same number of specimens (301 and 265, respectively).
H. norvegicus is distributed in the northern Atlantic and Arctic Ocean (Sars, 1902), in the study in Park (2000) the species was found in the Norwegian Sea and northwestern Atlantic down to 38° N.
H. austrinus, H. pustiulifer, H. spinosus, H. paraspinosus are confined to the Southern Ocean. H. austrinus , H. pustulifer are distributed mainly in the Antarctic waters, while H. spinosus, H. paraspinosus seem to be subantarctic species, whoch occurred in author's samples around South Africa, the southern end of South Africa and New Zealand. |
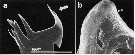 Issued from : S. Ohtsuka & S. Nishida in Copepod biodiversity in Japan: Recent Advances in Japanese Copepodology. In Matakawa M. & Kajihara H. eds. Springer-Verlag, 2017, Chapter 22. [p.582, Fig. 22.5, a, b]. a, Mandibular cutting edge of specialized carnivorous genus Heterorhabdus, hypodermic needle-like structure (arrowed); b, openings of the venom gland (1) and other secretory glands (2, 3) on posterior surface of the labrum. Nota: Heterorhabdus and related genera have a pair of fangs on the mandibular gnathobases (Nishida & Ohtsuka, 1996). Venom or anesthetic is released from two plandular cells via paired pores on the posterior surface of the labrum. These glands may be modified salivary glands homologous with those of particle-feeding heterorhabdids such as Disseta and Mesorhabdus (Ohtsuka & al., 1997). | | | | | (4) Heterostylites Sars, 1920 | |
| | Ref.: | Sars, 1920 c (p.11); 1925 (p.237); Sewell, 1932 (p.300); Rose, 1933 a (p.207); Farran, 1948 d (n°15, p.3); Davis, 1949 (p.60); Brodsky, 1950 (1967) (p.357); Tanaka, 1964 a (p.23); Heptner, 1972 b (p.58); Razouls, 1982 (p.473); Gardner & Szabo, 1982 (p.377); Mauchline, 1988 (p.707); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.816); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve & al., 1999 (p.943, 945: spp. Key); Bradford-Grieve,1999 b (p.88, Déf.); Park, 2000 (p.37, 39: spp. Key, Rem.: p.143); Boxshall Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.312, spp. Key) | | Rem.: | Type: Heterostylites longicornis (Giesbrecht, 1889). Total: 6 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 4.133 mm (n = 12; SD = 1.0460) and the mean male size is 3.948 mm (n = 12; SD = 0.8596). The size ratio (Male : Female) is 0.955. The sex ratio (Female : Male) is 1. | Diagnosis of Heterostylites: 1 - Ventralmost blade on mandibular gnathobase only slightly thickened, separated from adjacent teeth by narrow gap, no wider than base of blade 2 - Mx2 with large, claw-like seta on distal coxal endite, plus short seta. |
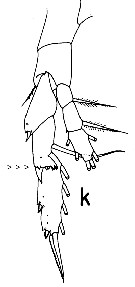 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal., San Diego, 2000, 31. [p.171, Fig. 19 K]. Heterostylites major female: P5 (anterior view). Note the characteristic of the 2nd exopodal segment with distal margin into a serrated lappet. Character considered as unique in the calanoid copepods. |
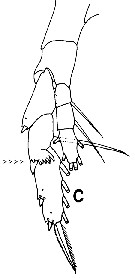 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal., San Diego, 2000, 31. [p.179, Fig. 27 c]. Heterostylites longioperculis female: P5 (anterior view). Note the characteristic of the 2nd exopodal segment with distal margin into a serrated lappet. |
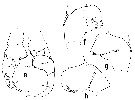 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal., San Diego, 2000, 31. [p.179, Fig. 27 e-h]. Heterostylites longioperculis male: e, P5 (posterior view; endopods omitted); f, 2nd exopodal segment of right leg (anterior view); g, same (posterior view); h, same (tilted clockwise). Note the greatly enlarged 2nd exopodal segment of the right P5 , characteristic of the genus. |
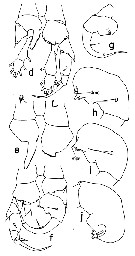 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal., San Diego, 2000, 31. [p.172, Fig. 20 d-j]. Heterostylites major male P5: d, legs with distal segments of right exopod omitted (anterior view); e, basipods of legs (anterior view; R = right, L = left); f, legs with endopods omitted (posterior view); g, exopod of right leg §anterior view); h, 2nd exopodal segment of right leg (posterior view); i, same (tilted counterclockwise); j, same (anterior view). dmc = distomedial corner; mpr = medial projection. | | | | | (5) Mesorhabdus Sars, 1905 | |
| | Ref.: | Sars, 1905 c (p.9); A. Scott, 1909 (p.132); van Breemen, 1908 a (p.126); Wolfenden, 1911 (p.312); Sars, 1925 (p.233); Sewell, 1932 (p.308); Rose, 1933 a (p.206); Farran, 1948 d (n°15, p.3); Tanaka, 1964 a (p.29); Heptner, 1972 a (p.57, Key spp.); Razouls, 1982 (p.474); Mauchline, 1988 (p.707); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.817); Mauchline, 1998 (p.70: F; p.74: M); Bradford-Grieve,1999 b (p.90, Rem.); Park, 2000 (p.23, Key spp., Rem.: p.143); Boxshall & Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.317, Key spp.) | | Rem.: | type: Mesorhabdus annectens Sars, 1905 (= Heterorhabdus brevicaudatus) Wolfenden, 1905. Total: 5 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 5.287 mm (n = 10; SD = 1.4503) and the mean male size is 5.326 mm (n = 9; SD = 1.5488). The size ratio (Male : Female) is about 1 (0.948 to 1.038). The sex ratio (Female : Male) is 1. | Diagnosis of Mesorhabdus: 1 - Ventralmost blade on mandibular gnathobase only slightly thickened, separated from adjacent teeth by narrow gap, no wider than base of blade 2 - Distal coxal endite of Mx2 with 3 unmodified setae. 3 - Basal and 1st endopodal endite of Mx2 each with 1 large, claw-like seta. Coxal endite of Mx1 with 1 seta. |
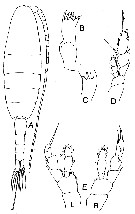 Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.91, Fig.58]. Mesorhabdus gracilis. Female A, habitus (dorsal view); B, right Md blade; C, left Md blade); D, P5. Male: E, P5 (R = right leg; L = left leg) | | | | | (6) Microdisseta Heptner, 1972 | |
| | Ref.: | Heptner, 1972 a (p.60); Razouls, 1982 (p.463); 1993 (p.307); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve,1999 b (p.91, Def. Rem.); Park, 2000 (p.1, 7,141); Boxshall & Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.320) | | Rem.: | Boxshall & Halsey (2004, p.126) ne suivent pas l'opinion de Park sur l'incertitude de la place de ce genre dans cette famille. Total: 1 espèce: | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 0.740 mm (0.68 to 0.80 mm) and the mean male size is 0.74 mm (0.73 to 0.75 mm). The size ratio (Male : Female) is 1. The sex ratio (Female : Male) is 1. | Diagnosis of Microdisseta: 1 - Ventralmost blade on mandibular gnathobase only slightly thickened, separated from adjacent teeth by narrow gap, no wider than base of blade 2 - Distal coxal endite of Mx2 with 3 unmodified setae. 3 - Basal and 1st endopodal endite of Mx2 without large, claw-like elements. Coxal endite of Mx1 with at least 2 setae. 4 - Basal endite of Mx2 with claw-like element plus 1 seta. |
 Issued from : G.D. Grice & K. Hulsemann in J. Zool., 1965, 146. [p.246, Fig.17, f-J, l]. As Disseta minuta. Female : f, Md palpus; g, gnathal lobe of Md; h, Mx1; i, Mx2; j, Mxp. Male: l, anal somite and caudal rami. | | | | | (7) Neorhabdus Heptner, 1972 | |
| | Ref.: | Heptner, 1972 a (p.58, Key spp.); Razouls, 1982 (p.476); 1993 (p.307); Bradford-Grieve,1999 b (p.92, Déf., Rem.); Park, 2000 (p.57, 59: Key spp., Rem.: p.143); Boxshall & Halsey, 2004 (p.126); Vives & Shmeleva, 2007 (p.320) | | Rem.: | type: Neorhabdus latus (Sars, 1905). Total: 5 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 9.140 mm (n = 10; SD = 2.3632) and the mean male size is 8.483 mm (n = 6; SD = 0.005). The size ratio (Male : Female) is 0,972 (n = 4; SD = 0.0662). The sex-ratio (Female : Male) is 1,25. (Probably equal in the genus) | Diagnosis of Neorhabdus: 1 - Ventralmost blade on mandibular gnathobase thickened and curved, separated from remaining distal teeth by marked diastema. 2 - Mx2 with at most 1 strong claw-like element on basis and on distal endite of coxa; proximal coxal endite with 3 or more setae. Middle endite of syncoxa of Mxp represented by 1 to 3 short setae. 3 - Distal praecoxal endite of Mx2 represented by 2 setae, proximal endite lobate, with up to 3 setae. |
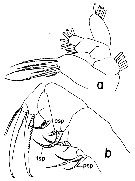 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.187, Fig.35 a, b]. Neorhabdus latus female: a, left Mx1 (posterior view); b, left Mx2 (posterior view; asp = anterior spine; psp = posterior spine; tsp = terminal spine. Mx1 with 6 setae on 1st inner lobe (= arthrite on praecoxa); no setae on 2nd or 3rd inner lobe; 3 or 4 setae on outer lobe; 2 or 3 setae on basis; 6 setae on endopod and 4 setae on exopod. |
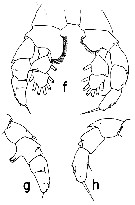 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.188, Fig.36 f-h]. Neorhabdus latus male: f, P5 (anterior view); g, exopod of right leg of P5 (posterior view); h, exopod of left leg of P5 (posterior view). | | | | | (8) Paraheterorhabdus Brodsky, 1950 | |
| | Ref.: | Brodsky, 1950 (1967) (p.345); Park, 2000 (p.71, 2 S-G: Antirhabdus & Paraheterorhabdus, spp. Key, Rem.: p.143); Boxshall & Halsey, 2004 (p.126) | | Rem.: | Total: 8 spp. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 3.516 mm (n = 16; SD = 0.9922) and the mean male size is 3.329 mm (n = 16; SD = 0.9397). The size ratio (Male : Female) is 0.952 (n = 8; SD = 0.0688) The sex ratio (Female : Male) is 1. | Diagnosis of Paraheterorhabdus: 1 - Ventralmost blade on mandibular gnathobase thickened and curved, separated from remaining distal teeth by marked diastema. 2 - Mx2 with 2 setal elements on basis and on distal endite of coxa; proximal praecoxal endite with 2 setae. Middle endite of syncoxa of Mxp typically represented by long, distally-curved seta. 3 - Mx2 with 1 claw-like setal elements + 1 small seta on basis. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.196, Fig.44 a-e]. Paraheterorhabdus (Paraheterorhabdus) robustus female: a, left Md (posterior view); b, masticatory edge of right Md (posterior view); c, left Mx1 (posterior view); d, left Mx2 (posterior view); e, right Mxp (anterior view; asp = anterior spine; psp = posterior spine). Nota: Exopod of Md reaching distal end of endopod; masticatory edge of left side, 1st tooth from basal spine tricuspid; 2nd bicuspid and about as long as 1st; 3rd ventralmost tooth approximately 2.8 times length of the corresponding tooth on right side Md. Edge between 2nd and 3rd teeth slightly convex. In masticatory edge of right side Md, 1st tooth from basal spine shortest, bicuspid; 2nd tricuspid, longer than either 1st or 3rd; 3rd single-pointed; 4th ventralmost tooth extending to same level as 3rd and separated from it by a smoothly hollow gap. Ventralmost teeth in both Md with a well-developed longitudinal rib along ventral margin. First inner lobe (= arthrite) of Mx1 with 2 anterior and 2 posterior submarginal and 8 marginal setae and an elevated pore on posterior surface close to proximomedial corner; seta of 2nd inner lobe (= coxal endite) long, extending beyond distal end of 1st inner lobe by1/2 its lrngth. Measured along medial margins, exopod about as long as rest of appendage. |
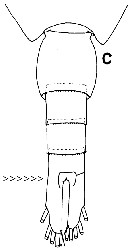 Isued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.195, Fig.43 c]. Paraheterorhabdus (Paraheterorhabdus) robustus female: c, urosome (dorsal view) Note the character of the subgenus,: left caudal ramus distinctly longer than right and completely fused with the anal somite (arrowed). |
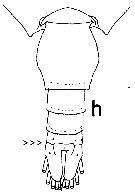 Isued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. Cal, San Diego, 2000, 31 [p.208, Fig. 56 h]. Paraheterorhabdus (antirgabdus) compactus female: h, urosome (dorsal view).
Note the character of the subgenus: left caudal ramus slightly longer than right and fully separated from anal somite (arrowed). | | | | | Antirhabdus Park, 2000 | | Ref.: | Park, 2000 (p.85, Def.) | | Rem.: | Type: Heterochaeta compacta Sars,1900. 1 species. | | | | Paraheterorhabdus Brodsky, 1950 | | Ref.: | Brodsky, 1950 (1967) (p.345, 346); Park, 2000 (p.71, Redef.) | | Rem.: | type: Heterorhabdus robustus Farran, 1908. 7 espèces | | | | | | | | | | | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2024. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 20 avril 2024] © copyright 2005-2024 Sorbonne Université, CNRS
|
|
|
|



;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
