|
|
 |
|
Calanoida ( Order ) |
|
|
|
Clausocalanoidea ( Superfamily ) |
|
|
| |
| | | |
| Diaixidae Sars, 1902 ( Clausocalanoidea ) | | Ref.: | Sars, 1902 (1903) (p.57); van Breemen, 1908 a (p.81); Gurney, 1931 a (p.85); Rose, 1933 a (p.159); Bradford, 1969 b (p.502, Table 3); Andronov, 1974 a (p.1005); 1979 (p.100, spp. Key); Connell, 1981 (p.493); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.377); 1993 (p.310); Bradford & al., 1983 (p.122); Brodsky & al., 1983 (p.144, 147); Mauchline, 1988 (p.734, 740, Rem.: cuticular pores); Huys & Boxshall, 1991 (p.54, 419); Othman & Greenwood, 1994 (p.989, Redef.); Bradford-Grieve & al., 1999 (p.881, 903, 933); Ohtsuka & Huys, 2001 (p.461); Ohtsuka & al., 2003 (p.53, 62, Rem.); Boxshall & Halsey, 2004 (p.15; 49; 95: Def., Genera Key); Ferrari & Markhaseva, 2005 (p.45, Rem.); Vives & Shmeleva, 2007 (p.638); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, molecular biology, phylogeny); Markhaseva & al., 2014 (p.63, 67, Table 1, 2, 3, 4, 5, Def., Rem. p.81); Laakmann & al., 2019 (p.330, Table 1, fig. 1, 2, Table A, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | Boxshall & Halsey (2004, p.96) exclude the Pseudophaenna genus from the family of Tharybidae to be included in this family.
10 G.: Anawekia, Byrathis, Diaixis, Procenognatha, Pseudophaennna, Ranthaxus, + 1 G (provisionally): Sensiava, Xancithrix, Thoxancalanus. For Laakmann & al., (2019, p.330, Table 1): 16 Genera after transfer of 7 genera prior in the Scolecitrichidae (Cenognatha, Falsilandrumius, Grievella, Landrumius, Neoscolecithrix, Paraxantharus, Xantharus).
The types of setae on the appendage Mx2 include this family in the 'Bradfordian' group of families.
For Markhaseva, Laakmann & Renz (2014, p.81) the family includes 15 genera: Byrathis, Cenognatha, Diaixis, Falsilandrumius, Grievella, Landrumius, Neoscolecithrix, Paraxantharus, Procenognatha, Ranthaxus, Sensiava, Xantharus, Xancithrix, Thoxancalanus. Anawekia Othman & Greenwood, 1994 is poorly described, and probably synonymus with Diaixis, but left in the family until the genus is revised. Precise differentiation between Diaixidae and Tharybidae is uncertain, and is based mostly on the enlarged and vaulted praecoxal arthrite of the Tharybis Mx1, which is, as proposed by Ferrari & Markhaseva (2005), probable an autapomorphy of Tharybis. The taxonomic status of the Diaixidae may be clarified when both sexes of all species have been discovered. The taxa here placed in the Diaixidae are ''ancestral'' to the remaining lineages of ''Bradfordians'' having the most plesiomorphic setation of the Mxp praecoxa (1, 2 and 3), a character state they share with Tharybis (Tharybidae).
Type genus: Diaixis Sars, 1902.
Diagnosis after Markhaseva & al. (2014, p.82) :
1 - Proximal part of the right A1 in males only ancestral segments X to XI fused (Cenognatha, Sensiava, Neoscolecithrix, Xancithrix), or segments X to XII fused (Byrathis, Diaixis, Paraxantharus, Procenognatha, Xantharus); the right A1 with a geniculation or morphologically transformed segments (Sensiava, Procenognatha).
2 - A2 coxa with 1 seta; basis with 2 setae; endopod segment 1 with 2 setae (1 seta in Sensiava, Diaixis (?) and some species of Byrathis); proximal exopod segment with 1 seta and setal formula of exopod 1, 1-1-1, 1, 1, 1, 1, 1, 3 (Byrathis, Cenognatha, Diaixis, Neoscolecithrix, Paraxantharus, Procenognatha, Ranthaxus, Thoxancalanus, or exopod setal formula 0, 0-1-1-1, 1, 1, 1, 1, 3 (Grievella), 0, 0-0-1, 1, 1, 1, 1, 1, 3 (Sensiava, Xantharus), 0, 0-0-1-1, 1, 1, 1, 1, 3 (Falsilandrumius, 0, 0-0-0-1, 1, 1, 1, 1, 3 (Landrumius, 0, 0-0-0, 1, 1, 1, 1, 1, 3 (Xancithrix).
3 - Md endopodal segment 2 with 9 setae (4-5, or 7 setae in Diaixis); exopod 5-segmented with 6 setae (with 5-6 setae in Byrathis, Falsilandrumius, Diaixis); gnathobase without hump/crest, or with hump in Byrathis (see Table 2).
4 - Mx1 with setal formula 11-14, 2-3, 4, 4-5, 7-12, 6-11, 9: it deviates in Diaixis, Byrathis at praecoxal arthrite (8 or 9-13 setae respectively), at proximal and distal basal endites (2-3 setae), and at epipodite (7-9 or 8-9 setae respectively), differs at setation of coxal endite in Falsilandrumius, Sensiava (4 setae) and Thoxancalanus, Xancithrix (5 setae); distal basal endite separate from endopod (see Tableb 3).
5 - Mx2 endites setal formula 5, 3, 3, 3, 4, (praecoxal endite with 4-5 setae in some species of Byrathis, Neoscolecithrix, Xantharus and with 4 setae in Diaixis); endopod setal formula diverse: 5w (worm-like) +3br (brush-like) + 1sc (sclerotized seta) (Neoscolecithrix), 5w + 3br (Byrathis), 3sc + 6br (Procenognatha), 3w + 5br (Buyrathis, Diaixis, Cenognatha, Sensiava, Xantharus), 3w + 6br (Falsilandrumius, Grievella, Landrumius), 3w + 5br + 1sc (Paraxantharus, Xancithrix, Xantharus, Thoxancalanus), or 6w (Ranthaxus) (see Table 4).
6 - Mxp praecoxal lobes setal formula 1, 2, 3, with all setae sclerotised (Grievella) fig.6e); 1sc + 2sc + (2sc +1w); 1sc + 2sc + (2sc + 1br); 1sc + (1sc + 1w) + (2sc + 1w); 1sc + (1sc + 1w) + (2sc + 1 br); 1w + (1sc + 1w) + (2sc +1w); 1w + (1sc + 1w) + (2sc + 1br); 1w + (1sc + 1br) + (2sc + 1br); basis with 3 proximal and 2 distal setae (Table 5).
7 - Male P5 biramous or uniramous; in Diaixis of complex structure while of simple structure in the remaining diaixid genera. Right and left coxa and basis asymmetrical (Procenognatha, Cenognatha, Neoscolecithrix, Xancithrix , in the three latter genera of the phaennid type), or nearly same length (Byrathis, Paraxantharus, Sensiava, Xantharus); exopods 3-segmented, except for 2-segmented one on the right in Cenognatha and Diaixis, and on the left in Sensiava.
For Markhaseva & al. (2014, p.83) the taxa placed in the Diaixidae are ''ancestral'' to the remaining lineages of ''Bradfordians'' having the most plesiomorphic setation of the maxilliped praecoxa (1, 2 and 3), a character state they share with Tharybis (Tharybidae). The Diaixidae have the greatest number of genera (8) among the ''Bradfordians'', with the most plesiomorphicd antenna exopod armament 1, 1-1-1, 1, 1, 1, 1, 1, 3 (which they share with the ''scolecitrichid'' genus Omorius and the taxonomically unresolved Puchinia). Many diaixids have the most plesiomorphic setation of the maxilla endopod with 9 setae (which they share with the phaennids Brachycalanus and Xanthocalanus, versus usually 8, rarely 6 setae or less in other ''Bradfordians'').
Precise differentiation between Diaixidae and Tharybidae is uncertain, and is based mostly on the enlarged and vaulted praecoxal arthrite of the Tharybis maxillule, which is, as proposed by Ferrari & Markhaseva, 2005, probable an autapomorphy of Tharybis (ca. a derived character unique to a given species or other monophyletic taxon).
The taxonomic status of the Diaixidae may be clarified when both sexes of all species have been discovered.
Markhaseva (2015) creates the new genus Vensiasa.
Pseudophaenna (Sars, 1902) is conside as incertae sedis genera in the '' Bradfordian'' genera (see Markhaseva & Ferrari, 2005; Markhaseva & al. , 2014). |  Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.70, Fig.5]. Schematic view of right A1 in males of ''Bradfordian'' families. Diaixidae. Roman numerals: ancestral segments; black arrows: fused ancestral segments; black dotted arrows: position of geniculation or morphologically transformed segments. A1 of Sensiava and Procenognatha schematised after Markhaseva & Schulz (2006, 2010), remaining genera after Markhaseva (pers. obs.). |
 Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.73, Table 1]. A2 armament (number of seta) in different ''Bradfordian'' genera (females). Diaixidae. c: coxa; b: basis; End1: endopodal segment 1; End2: endopodal segment 2; Exp: exopod. |
 Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.74, Table 2]. Md armament (number of seta) in different ''Bradfordian'' genera (females). Diaixidae. b: basis; End1: endopod segment 1; End2: endopod segment 2; Exp: exopod; gn: gnathobase. |
 Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.76, Table 3]. Mx1 armament (number of seta) in different ''Bradfordian'' genera (females). Diaixidae. pa: praecoxal arthrite (setal formula of praecoxal arthrite: terminal+posterior+anterior setae); ce: coxal endite; bp: proximal basal endite; bd: distal basal endite; End: endopod; Exp: exopod; Epi: epipodite.. |
 Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.78, Table 4]. Mx2 armament (number of seta) in different ''Bradfordian'' genera (females). Diaixidae. at: attenuation; pe: praecoxal endite; ce: coxal endite; bp: proximal basal endite; bd: distal basal endite; el: enditic-like lobe of proximal endopopdal segment; End: endopod; w: worm-like seta; br: brush-like seta; sc: sclerotised seta. |
 Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.80, Table 5]. Mxp setation (number of seta) in different ''Bradfordian'' genera (females). Diaixidae. at: attenuation; pes: praecoxal endites of syncoxa (from proximal to distal); ces: coxal endite of syncoxa; bp: basis proximal; bd: basis distal; End: endopod; . For more detailed morphology of the setae on the praecoxal of the maxilliped syncoxa see Markhaseva & Ferrari (2005) and Markhaseva & al. (2008). |
 Issued from : E.L. Markhaseva, S. Laakmann & J. Renz in Mar. Biodiv., 2014, 44 [p.82, Fig.8]. P5 of males in Diaixidae. a: Xancithrix ohmani Markhaseva 2012; b: Neoscolecithrix caetanovi Alvarez 1985; c: Cenognatha farrani (Smirnov 1935); d: Procenognatha semisensata Markhaseva & Schulz 2010; e: Sensiava longiseta Markhaseva & Schulz 2006; f: Xantharus formosus Andronov 1981; g: Xantharus renatehaassae Schulz 1998; h: Paraxantharus brittae Schulz 2006; i: Byrathis arnei Schulz 2006; j: Diaixis trunovi Andronov 1981; k: Diaixis helenae Andronov 1981. Exp: exopod; End: endopod; R: right leg. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.95]. Armature formula of swimming legs P1 to P4. Setation sometimes reduced, exopodal armature of P1 sometimes sexually dimorphic. Nota: Female P5 2-segmented with single apical spine in Pseudophaenna, absent in other genera. - Male P5 uniramous, qzymmetrical with bilobed proximal plate formed by fusion of intercoxal sclerite and protopodal segments in Diaixis, right leg typically 2-segmented, consisting of slender proximal segment fused to proximal plate and movable distal segment, often claw-like: left leg often subchelate; basis free or fused to coxal plate, exopod typically 3-segmented with apical segment often subchelate; slender unarmed 1st segment, slender 2nd segment often bearing distal processes and short 3rd segment armed variously with setae, lamellae or spinous processes. P5 slender in Pseudophaenna, 4-segmented on right, 5-segmented on left. - Eggs presumably released into water. |
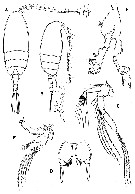 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.97, Fig.16]. Diaixidae. A, Diaixis hibernica habitus female; B, habitus male; C, female A2; D, Pseudophaenna typica female P5; E, Diaixis hibernica female Md; F, male P5. [Sars, 1902]. | | | | | (1) Anawekia Othman & Greenwood, 1994 | |
| | Ref.: | Othman & Greenwood, 1994 (p.989); Mauchline, 1998 (p.78: F; p.81: M); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.96); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type: Anawekia spinosa Othman & Greenwood, 1994. Total: 3 spp.
Definition of Anawekia from Othman & Greenwood (1994, p.989) :
- Body small in size (<< 1 mm), robust, oval in dorsal view.
- Head and 1st pedigerous somite separated, 4th and 5th pedigerous somites separate.
- Head (in lateral view) anterior margin curves almost at right angles to dorsal margin.
- Urosome short, 0.25 length of prosome ; of 4 somites in females ; 5 somites in males.
- Caudal ramus with 4 subequal apical setae.
- A1 24-segmented.
- A2 exopod 7-segmented, twice length of endopod.
- Md gnathobase large and with complex cutting edge.
- Mx2 and Mxp with vermiform sensory filaments.
- P1 to P4 of both sexes biramous ; legs usually covered with spinules on both anterior and posterior surfaces.
- P1 endopod 1-segmented ; outer marginal spine on 1st exopod segment present only in females.
- Endopod of P2 2-segmented ; P3 and P43-segmented.
P5 absent in females.
- P5 males uniramous, asymmetrical, chelate and with common baipod ;. Right leg 2-segmented, terminal segment with digitiform process ; left leg 3-segmented, ends in chela.
After Othman & Greenwood (1994, p.989) the genus Anawekia can be distinguished from Diaixis s.s. by there are vermiform sensory filaments present on the 1st segment of the Mxp in Anawekia, but not in Diaixis s.s. ; the head is separated from the 1st pedigerous somite in Anawekia, fused in Diaixis s.s. ; the male A1 is 23-segmented in Diaixis s.s., 24-segmented in Anawekia ; P5 male has a row of comb-like spines on the basipod in Diaixis s.s., these spines being absent in Anawekia
RThe new genus can be distinguished from Diaixis s.l. by the combination of features : A2 exopod 6-segmented in Anawekia, 7-segmented in Diaixis s.l. ; bulb type aesthetascs are present on Mx2 of Diaixis s.l., absent in Anawekia. | | Remarks on dimensions and sex ratio: | | The mean female size is 0.860 mm (n = 6; SD = 0.0787) and the mean male size is 0.743 mm (n = 6; SD = 0.0408). The size ratio (male : female) is 0.866 (n = 3; SD = 0.0271). The sex ratio is 1. | | | | (2) Byrathis Markhaseva & Ferrari, 2005 | |
| | Ref.: | Markhaseva & Ferrari, 2005 a (p.128: Def., fig.31, Rem.); Schulz, 2006 (p.67: Rem.); Markhaseva & Renz, 2011 (p.49, 66: Rem.); Markhaseva & al., 2014 (p.81, Table 1, 2, 3, 4, Rem.); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type: Xanthocalanus macrocephalon. Grice & Hulsemann, 1970. Total: 7 spp. + 1 unident.
Diagnosis female from Markhaseva & Ferrari (2005, p.128) :
- Cephalosome and pediger 1, pedigers 4 and 5 fused dorsally, separated laterally.
- genital complex with small spinules dorsally and laterally.
- A1 24-segmented.
- A2 coxa and basis with 1 and 2 setae respectively; exopod with 1, 3, 1, 1, 1, 1, 1, 3.
- Md gnathobase with broad medial edge and posterior tooth-like knob.
- Mx1 praecoxal endite with 2 posterior setae and 9 terminal setae; proximal 2 terminal setae curved proximally; coxal and proximal basal endite with 2 setae; endopod articulating with basis.
- Mx2 praecoxal endite with 4 sclerotized setae; proximal basal endite with 1 worm-like seta; distal basal endite + ramus with 5 worm-like setae and 3 short brush-like setae, 1 of which is thicker.
- Mxp praecoxal endites with 1, 2 and 3 setae (distal endite with 1 worm-like seta).
- P1-P4 of clausocalanoidean segmentation and setation.
- P5 3-segmented, distal segment with 3 setae and terminal attenuation.
- Male unknown.
For Markhaseva & Ferrari, a synapomorphy for Byrathis is proximal 2 terminal setae on praecoxal endite of Mx1 curved proximally.
The following plesiomorphies or convergences help separate Byrathis from most other ''bradfordian'' genera: segments of the exopod of A2 with 1, 3, 1, 1, 1, 1, 1, 1 setae (shared with Neoscolecithrix, Cenognatha, some species of Diaixis, Puchinia, Omorius); more than 4 worm-like sensory setae on basal endite + ramus of Mx2 (shared with Brodskius, Neoscolecithrix, Rythabis, Omorius); praecoxal endites of syncoxa of Mxp with 1, 2, 3 setae (shared with Grievella, Xantharus, Falsilandrumius, Landrumius, Neoscolecithrix, Cenognatha, some species of Diaixis and some species of Tharybis); A worm-like seta on the distal praecoxal endite of Mxp (shared with some species of Diaixis). The posterior tooth-like knob on gnathobase of Md (shared with Omorius) has not been surveyed well enough to determine its usefulness for separating groups of ''bradfordian'' genera; e.g. it is present in Scolecithrix danae, Lophothrix frontalis, Scottocalanus helenae).
After Markhaseva & Renz (2011, p.66) the genus Byrathis can be divided into 2 groups :
1- B. divae, B. penicillatus, B. laptevorum. Medium sized copepods ; the genital double-somite has no surface ornamentation ; the Mx1 proximal basal endite and exopod are with 3 and 6 (or more) setae, respectively ; the Mx2 endopod bears 5 brush-like sensory setae.
2- B. macrocephalon, B. volcali, B. sp. Smaller size than the first group ; presence of surface ornamentation on the genital double-somite ; the M1 proximal basal endite and exopod with 2 and 5 (or less) setae, respectively ; the Mx2 endopod with only 3 brush-like sensory setae.
For the authors, the adult male B. arnei which is described for the first time, is distinguished from all other clausocalanoideans that possess sensory setae on the Mx1 and Mxp (bradfordian families) in the morphology of A1 and P5. Right P5 and left A1 are symmetrical, 22-segmented. In this genus the ancestral segments XXII-XXIII are fused on both sides (apomorphy). This is a unique case for bradfordian genera which known males in which segments XXII-XXIII are fused only in the right A1 as in Scolecitrichidae (stricto sensu) and Diaixidae s.l. (sensu Markhaseva & Ferrari, 2005), and as in Brodskius, Tharybis, Plesioscolecithrix and Xanthocalanus. In Kirnesius they are fused only in the left A1.
The authors put in the genus Byrathis in the family Diaixidae.
According to Markhaseva & Renz (2011, p.68) the morphology of Byrathis combines primitive and derived characters :
1- primitive character : Setation of A2 exopod proximal segments as 1, 1-1-1 in both sexes ; Mx2 praecoxal endite (in B. arnei) female with 5 setae ; females Mxp praecoxal endites setation with 1, 2, 3 setae from proximal to distal, and male coxa and basis nearly symmetrical.
2 derived character : Md gnathobase of female with crest ; Mx1 distal basal endite with only 2-3 setae.
Apomorphies for the genus are male A1 with both ancestral segments XXII-XXIII fused and female M1 praecoxal arthrite with 2 proximal marginal setae curved proximally.
The mosaic combination of primitive and derived features is an example of heterobathmy typical of these benthopelagic clausocalanoideans.
| | Remarks on dimensions and sex ratio: | | The mean female size is 1.951 mm (n = 12; SD = 1.2346). The mean male size for one male only is 1.475 mm, and the size ratio for the male and female is 1. The sex ratio (female: male) provisionally 8. | | | | | | Ref.: | Sars, 1902 (1903) (p.58); van Breemen, 1908 a (p.81); Rose, 1933 a (p.160); Andronov,1979 (p.100, clé spp.); Connell, 1981 (p.493); Razouls, 1982 (p.377); Bradford & al., 1983 (p.122); Mauchline, 1988 (p.734); Razouls, 1993 (p.310); Othman & Greenwood, 1994 (p.988, Redef., 2 groupes: 'pygmaea' et 'centrura'); Mauchline, 1998 (p.78, 88: F; p.81, 89: M); Bradford-Grieve & al., 1999 (p.881, 934: spp Key.); Andronov, 2002 (p.2, 11, clé spp.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.96); Markhaseva & Ferrari, 2005 a (p.111, Fig.31, Rem.); Vives & Shmeleva, 2007 (p.639, spp. Key); Markhaseva & al., 2014 (p.81, Table 1, 2, 3, 4, Rem.); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type: Scolecithrix hibernica A. Scott, 1896.
more or less hyperbenthic forms. 8 spp. | | Remarks on dimensions and sex ratio: | | The mean size of the females is 0,913 mm (n= 8; S= 0,144; Cv= 0,158) and of the males 0,791 mm (n= 7; S= 0,091; Cv= 0,115). The size ratio (M/F) is 0,910 or 91% (n= 7; S= 0,045; Cv= 0,05). The sex ratio (F/M) is 1,29. | | | | | (4) Procenognatha Markhaseva & Schulz, 2010 | |
| | Ref.: | Markhaseva & Schulz, 2010 (p.4); Markhaseva & al., 2014 (p.82, Table 1, 2, 3, 4, Rem.); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | type species: Procenognatha semisensata. Total: 1 sp.
After Markhaseva & Schulz (2010, p.5) the genus is defined by :
1- praecoxal arthrite of Mx1 with proximalmost posterior seta swollen in its basal part (vs. not swollen in other bradfordians) ; the latter character is an apomorphy of the genus.
2- Mx2 endopod with 3 sclerotized terminal setae (vs. worm-like sensory setae in remaning bradfordian genera).
3- Mx2 endopod with 9 setae (3 sclerotized setae + 1 sclerotized and brush-like seta + 5 brush-like setae).
Right A1 male with traces of geniculation.
A high number and unique combination of plesiomorphic characters exhibited by this genus suggest that the type species is the most primitive among bradfordian taxa. The following combination of characters distinguishes Procenognatha from all other bradfordian genera :
1- ancestral setation of A2 exopod (1, 1-1-1, 1, 1, 1, 1, 1 and 3).
2- Mx2 endopod with a total number of 9 setae.
3- presence of 3 sclerotized setae on Mx2 endopod terminally (a unique plesiomorphy for bradfordians).
4- syncoxa of Mxp with ancestral setation formula (1, 2, 3).
5- Male A1 with traces of geniculation ; oral parts not reduced ; biramous P5.
Procenognatha is closely related to Cenognatha. They share the following combination of characters:
1- short rostrum with 2 filaments.
2- biramous right P5 in the male.
3- a similar setation and segmentation pattern of the oral parts.
Procenognatha is distinguished from Cenognatha by rounded posterior corners of the prosome (vs. extended into 2 spines in Cenognatha) and the details of setation of oral parts: a- endopod segment 1 of Md with 2 setae (vs. 3 setae in Cenognatha); b- Mx1 endopod with 12 setae and Mx1 exopod with 7 setae (vs. 7, 9 or 11; 9-10 setae in Cenognatha, respectively); c- setal number (9) and composition of Mx2 endopod plus distal basal endite setae (vs. 7-8 setae in Cenognatha). | | Remarks on dimensions and sex ratio: | | For 0ne species, total Female: 2.50-2.75; Male: 1.95-2.30. Length ratio male : female = ± 0.81. The sex ratio is 1. | | | | | (5) Pseudophaenna Sars, 1902 | |
| | Ref.: | Sars,1902 (1903) (p.43); van Breemen, 1908 a (p.57); Rose, 1933 a (p.126); Bradford, 1973 (p.138, Rem.); Razouls, 1982 (p.287, 368); Bradford & al., 1983 (p.123, Déf., p.126); Razouls, 1993 (p.311); Mauchline, 1998 (p.87: F; p.89: M); Vyshkvartzeva, 2001 (p.77: Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.96); Markhaseva & al., 2014 (p.83, Table 1, 2, 3, 4, Rem.: p.84); Laakmann & al., 2019 (p.330, Table 1: considered as incertae sedis genera) | | Rem.: | Type: Pseudophaenna typica Sars, 1903.
Total: 1 sp.
Diagnosis from Bradford & al. (1983, p.126), after Sars, 1903 :
1 - Head and pediger segment 1 fused, pediger segments 4 and 5 separate.
- Rostrum conical.
- A1 23-segmented in female; 21-segmented in male.
2 - A2 exopod more than twice length of endopod.
3 - Mx1 exopod with 2 setae.
4 - Mxp powerfully built.
5 - Posterior mouthparts of male reduced.
6 - Swimming leg endopods without spines on posterior surface.
7 - Female P5 symmetrical and 3-segmented.
8 - Male P5 asymmetrical, uniramous and multi-segmented on each side; right leg terminates in spine.
This genus, placed among Tharybidae, is transferred in this family by Boxshall & Halsey (2004, p.96, 210). Total: 1 sp.
For Markhaseva, Laakmann & Renz (2014, p.81) this genus must be placed outside Tharybidae, his status remains taxonomically unresolved. After Boxshall & Halsey (2004, p.96) this genus shares synapomorphies with Diaixidae. However, their proposed synapomorphic characters for Pseudophaenna can also be found in other ''Bradfordians'' and do not identify this genus as diaixid. A more detailed study of Pseudophaenna is necessary to verify the taxonomic status of this genus. Characters: 1 - Reduction in antennary endopod (shared with Diaixis of Diaixidae, Tharybis of Tharybidae and Rythabis) ; 2 - loss of posterior row of setae from maxillary arthrite (shared with Diaixis, Phaenna of Phaennidae, Rostrocalanidae and Bradfordiella); 3 - absence of brush-like sensoriform elements from 3rd syncoxal endite of Mxp (except for some diaixids, this character is shared with Phaennocalanus of Phaennidae, Omorius, Diaiscolecithrix and Archeoscolecithrix from Scolecitrichidae, Rostrocalanidae and Kyphocalanidae). The armament of Mx2 endopod is unconfirmed in Pseudophaenna and therefore cannot be used to clarify its relationship to other ''Bradfordians'' families.
According to Boxshall & Halsey (2004) Mxp praecoxal enbdites setal formula is 0, 2, and 2. This differs from the Diaixidae, which are characterised by a setal formula of 1, 2 and 3. Pseudophaenna shares the Mxp praecoxal endites setal formula with only some species of Undinella (Tharybidae), but the following characters prevent Pseudophaenna to be placed in the Tharybidae: 1 - A2 endopod segment 1 without setae (versus 2 setae in Tharybis); 2 - Md endopod segment 1 without setae ( versus 2 setae in Tharybis); 3 - Mx1 setal formula 9, 0, 2, 3, ?, 6, 2, 7 (versus tipically 13-14, 2-3, 3-4, 2-3, 7 or 9, 3-5, 7 or 9 in Tharybis); 4 - Mx2 praecoxal endite with 5 setae (versus 4 in Tharybis; 5 - Mxp praecoxa setal formula 0, 2, 2 (versus 1, 2 and 3 in Tharybis; 6 - Male P5 of a simple structure and uniramous (versus P5 left biramous, right uniramous, of complex structure in Tharybis.
For Markhaseva, Laakmann & Renz (2014, p.84), the following characters prevent the insufficiently described genus from been placed in the Tharybidae: 1 - A2 endopodal segment 1 without setae (versus 2 setae in Tharybis); 2- Md endopodal segment 1 without setaze ( versus 2 in Tharybis); 3 - Mx1 setal formula: 9, 0, 2, 3?, 6, 2, 7 (versus typically 13-14, 2-3, 3-4, 2-3, 7 or 9, 3-5, 7 or 9 in Tharybis); 5 - Mxp praecoxa setal formula: 0, 2, 2 (versus 1, 2 and 3 in Tharybis (see Table 1-5 in); 6 - male P5 of a simple structure and uniramous (versus P5 left biramous, right uniramous, of complex structure in Tharybis).
A more detailed study of Pseudophaenna is necessary to verify the taxonomic status of this genus. | | Remarks on dimensions and sex ratio: | | Only one specimen measured: female body length : 1,60 mm and male: 1,40 mm. The size ratio (male: female) is 0.875. The sex ratio is 1. | | | | | (6) Ranthaxus Markhaseva & Schulz, 2010 | |
| | Ref.: | Markhaseva & Schulz, 2010 (p.13, Table 1); Markhaseva & al., 2014 (p.82, Table 1, 2, 3, 4, Rem.); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, biogeography); Laakmann & al., 2019 (p.330, Table 1). | | Rem.: | Total: 1 sp.
issued from : E.L. Markhaseva & K. Schulz in Proc. Zool. Inst. RAS, 2010, 314 (1). [p.13, 17]
Remarks : Differential diagnosis.
- Mx1 : 1 worm-like sensory seta present additionally to sclerotized setae on distal basal endite and endopod (vs. Sensory setae absent on distal basal endite in other bradfordians).
- Mx2 : Endopod plus distal basal endite with 4 long plus 2 short worm-like sensory setae (vs. 6 worm-like sensory setae of equal length in Rostrocalanus).
- Mxp : Very long worm-like sensory seta on distal praecoxal endite, reaching the distal part of basis (vs. Worm-like seta short or absent on distal endite in other bradfordians). Endopod segment 1 (endopodal segment incorporated into basis) and terminal segment with 1 of setae on each segment transformed into sensory and worm-like setae (vs. Sensory setae absent from endopod in other bradfordians).
This genus is closely related to Xantharus and Paraxantharus Schulz, 2006 and shares with both genera the same number of setae on most of endites and segments of oral parts (See Table1 for these genus). However, the new genus is well distinguished from these genera by the presence of worm-like sensory setae on Mx1, Mxp basis and endopod, a smaller number (6) of sensory setae on Mx2 endopod, and by very long worm-like sensory setae on Mxp syncoxa.
This genus shares an apomorphy (character derived from and differing from an ancestral condition) with Xantharus in that segments 19 and 20 (XXII and XXIII) of A1 are markedly reduced in size compared to segment 18 (XXI) and in a very long aesthatasc on segment 18, but it differs in the primitive type of exopod setation of A2 (1, 1-1-1, 1, 1, 1, 1, 1, and 3), which Ranthaxus shares with Paraxantharus (see Table 1).
The presence of sensory setae on the distal praecoxal endite and coxal endites of Mx2 is shared with Puchinia Vyshvartzeva (1989), however, Puchinia is not allied to the new genus by number of characters. | | Remarks on dimensions and sex ratio: | | The body size for one female only is 1.200 mm. Male unknown. | | | | | (7) Sensiava Markhaseva & Schulz, 2006 | |
| | Ref.: | Markhaseva & Schulz, 2006 (p.1); Markhaseva & Renz, 2011 (p.67, Rem.); Markhaseva & al., 2014 (p.82, Table 1, 2, 3, 4, Rem.) ; Renz & Markhaseva, 2015 (p.96, Table 4, Fig.3, sex ratio, biogeography); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type species: Sensiava longisetaMarkhaseva & Schulz, 2006. Total: 3 spp. + 1 undet.
Temporarily included by the authors in Diaixidae family, but it is very probable that after discovery of the female a new family may have to be established.
For Markhaseva & Schulz (2006, p.13) excepting the A1, the new genus fits the diagnosis of the superfamily Clausocalanoida. However, in respect of the male grasping A1 Sensiava longiseta contradicts its diagnosis. The geniculated A1 of the new genus is the most striking example of a modified risht A1 in males of Clausocalanoidea, but not the only recorded case among the representatives in the superfamily. The male of the rare aetideid benthopelagic genus Azygokeras has morphologically modified ancestral segments XIX-XXVI in right A1 (Koeller & Littlepage, 1976) ; Markhaseva & Ferrari, 2005), a tharybid genus described from close to the sea floor, also has the right A1 modified . Fusion of ancestral segments XXII-XXIII of the right A1 might be considered remnants of an ancestral geniculation (Schulz, 2005) and are observed in a number of Clausocalanoidean taxa (Kirnesius, Scolecitrichopsis, Tharybis, Xantharus, Xanthocalanus and some species of Aetideopsis. As a consequence the diagnosis of the superfamily Clausocalanoidea should be emended by exclusion of the non-geniculate A1 as a diagnostic character.
Discovery of a new benthopelagic copepod genus in vicinity of the floor of the deep-sea demonstrates that the diagnoses of some families (e ;g., Bradfordian families), which have been based on the morphological characters of their pelagic representatives, are inadequate.
Diagnosis from Markhaseva & Schulz (2006, p.3);
Male:
- Cephalosome and pediger 1, and pedigers 4 and 5 incompletely separated.
- Rostrum with 2 long filaments.
- A1 asymmetrical, right one 23-segmented and geniculate, left 24-segmented.
- Mouthparts well developed.
- Praecoxal endite of Mx2 with 5 setae; endopod with 8 sensory setae (3 worm-like, 5 brush-like).
- Praecoxal lobes of Mxp syncoxa with 1 sclerotized seta on proximal lobe, 2 sclerotized setae on medial lobe and 3 setae (2 sclerotized, 1 poorly sclerotized) on distal lobe, setal formula 1, 2, 3.
- Terminal endopodal segment of Mxp with 4 setae (2 thick and long, about 1.6 times longer than basis).
- P1 endopod 1-segmented, P2 endopod 2-segmented, P3-P4 endopods 3-segmented.
- P1-P4 exopods 3-segmented.
- P1 exopodal segment 3 with 3 medial setae; P2-P4 exopodal segment 3 with 4 medial setae.
- P2-P4 endopods with posterior surface spinulated.
- P2-P3 basis and 1st and 2nd exopod segments with distal rows of spinules.
- P4 basis, endopods and exopods spinulated.
- Right P5 uniramous, exopod 3-segmented.
Female unknown.
For Markhaseva (2014, p.218) the genus is distinguished from other diaixids by :
1- Males right A1 of 23 segments with geniculation.
2- in females and males distalmost terminal seta of Mx1 praecoxal arthrite curved at the apex.
3- In males 2 terminal endopodal setae of Mxp are very long, more than 1.6 times longer than basis.
4- male P5 of complex structure, left leg biramous, exopod 2-segmented.
Additional combination of characters from Markhaseva (2014, p.218) that define the genus among diaixids :
1- males A1 ancestral segments X-XI fused (shared with Cenognatha, Neoscolecithrix, Xancithrix) .
2- A2 endopodal segment 1 with 1 seta (shared with Diaixis ? and some species of Byrathis).
3- A2 exopod setal formula 0, 0-0-1, 1, 1, 1, 1, 1, 3 (shared with Xantharus), or 0, 0-1-1, 1, 1, 1, 1, 1, 3 (shared with Grievella).
- In females Mx1 coxal endite with 4 (shared with Falsilandrumius), or 5 setae (shared with Xancithrix, Thoxancalanus).
5- Mx2 endite setal formula : 5 setae (or 4 plus attenuation), 3, 3, 3, 4 setae.
6- Setae of the Mx2 basal endites and enditic-like lobe of endopod similar in shape and size, curved or curled distally.
7- proximal basal Mx2 endite with 1 seta worm-like (shared with Ranthaxus) .
8- Mx2 endopod setal formula : 3w + 5br (shared with Paraxantharus, Xancithrix, some species of Xantharus, and Thoxancalanus).
9- Female P5 uniramous, 3-segmented, terminal segment with 4 spines, medial spine the longest.
10- Male P5 right and left coxa and basis of the nearly same lenfgth (shared with Byrathis, Paraxantharus, Xantharus).
| | Remarks on dimensions and sex ratio: | | The mean female size is 2.992 mm (n = 6; SD = 1.0268), and in male 3.8 mm (n = 4; SD = 1.0456). The size ratio (Male: Female) is 0.93. | | | | | (8) Thoxancalanus Markhaseva, Laakmann & Renz, 2013 | |
| | Ref.: | Marhkhaseva & al., 2014 (p.65, Def., Table 1, 2, 3, 7, Rem.: p.82); Renz & Markhaseva, 2015 (p.96, Table 4, Fig.3, sex ratio, biogeography); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type: Thoxancalanus spinatus. Total: 1 sp.
Diagnosis after Markhaseva & al. (2014, p.65) :
- P5 basis and exopod ornamented with scale-like spinules (apomorphy for the genus).
- The following combination of plesiomorphic characters is diagnostic for the genus:
- A2 exopod setation formula: 1, 1-1-1, 1, 1, 1, 1, 1, 3.
- Md: basis with 4 setae, endopod segment 1 with 3, segment 2 with 9 setae; exopod 5-segmented.
- Mx1 with setal formule: 14, 5, 4, 5, 11, 8 and 9 setae.
- Mx2 with endites setal formula: 5, 3, 3, 3, 4 and endopod with 3 worm-like + 5 brush-like + 1 small sclerotized sensory setae.
- Mxp: praecoxal endites setal formula: 1, 2 and 3 setae; basis with 3 proximal and 2 distal setae, respectively; endopod 5-segmented with 4, 4, 3, 3+1, and 4 setae.
- Posterior surfaces of exopod segments 2 of P2-P3with oblique row of flat spines, in other features swimming legs as typical of Clausocalanoidea.
- P5-3 segmented, exopod with 4 articulated spines.
- Male unknown. | | | | | (9) Vensiasa Markhaseva, 2015 | |
| | Ref.: | Markhaseva, 2015 (p.183); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type species: Vensiasa incerta Markhaseva, 2015.
Diagnosis from Markhaseva (2015, p.183):
Female:
- Cehalosome and pediger 1, pedigers 4 and 5 incompletely separate.
- posterior corners, in dorsal view, triangular ; as rounded lobes, in lateral view.
- Rostrum as a poorly developed plate with 2 filaments.-
A1 24 free segments.
- Coxa of A2 with 1 seta; basis with 2 setae; endopodal segment 1 with 1 seta, exopod setal formula 0,0-0-1, 1, 1, 1, 1, 1, 3.
- Gnathobase of Md with 7-8 teeth; exopod 5-segmented with 1,1,1,1, and 2 setae; endopod segment 1 with 3 setae, 2nd endopod with 9 setae; basis with 3 setae.
- Mx1 praecoxal arthrite with 9 terminal spines, 2 posterior setae and 1 anterior seta; coxal endite with 3 setae, coxal epipodite with 8-9 setae; proximal basal endite with 3 setae, distal basal endite with 2-3 setae; endopod with 5 setae, exopod with 8 setae.
- Mx2 praecoxal endite bearing 5 setae and attenuation, coxal and basal endites with 3 setae each; enditic-like lobe of proximal endopodal segment with 4 setae; 2 setae of distal basal endite and 2 setae of enditic-like lobe equal in length and curved distally; endopod with 6 worm-like (w) and 2 brush-like (br) sensory setae, 3 terminal worm-like setae long, 3 proximal worm-like setae short and very thin and 2 brush-like setae: 1 thick with brush well developed and 1 thin with poorly developed brush.
- Mxp syncoxa with 1, 2 and 3 setae on praecoxal endites.
- P1 to P4 typical clausocalanoidean setation and segmentation.
- P5 uniramous, symmetrical, 3-segmentesd, with 4 setal elements terminally.
Male:
- Rostrum as in female.
- Cephalosome and pediger 1, pedigers 4 and 5 separate.
- Prosome posterior corners as short rounded lobes.
- Right A1 23-segmented; left 24 free segments; ancestral segments X-XI fused.
- Oral appendages sexually dimorphic and moderately reduced compared to females.
- Setatio of A2 asin female.
- Number of setae on Md basis and endopod segment 2 as in female, endopod segment 1 with 2 setae.
- Armament of Mx1 differs from female in: praecoxal arthrite with 1 posterior seta; coxal epipodite with 7 setae; proximal basal endite with 2 setae, and exopod with 7 setae.
- Mx2 praecoxal endite with 4 setae, other lobes with the same setal numbers as in female.
- Mxp setation of syncoxa as in female, distal endopod setae more than 1.6 times longer than basis.
- P1 to P4 as in female.
- P5 nearly as long as urosome; protopods asymmetrical, right leg uniramous, 3-segmented; left leg biramous, endopod longer than 3-segmented exopod.
The new genus Vensiasa is defined among ''Bradfordians'' by the morphology of the sensory setae on the maxillary (Mx2) endopod (6w + 2br), their structure is an apomorphy for the genus: the 3 proximal worm-like setae are remarkably short and thin, 1 brush-like seta is very thin with poorly developed brush and 1 brush-like seta is very thick, brush well developed. Mx1 endopod with 5 setae (vs 7-12 in other diaixids) and male's P5 protopods asymmetrical; left P5 endopod longer than 3-segmented exopod (vs in other diaixids if protopods asymmetrical, then left endopod rudimentary or absent).
The new genus appears to be more closely related to the genus Sensiava. | | | | | (10) Xancithrix Markhaseva, 2012 | |
| | Ref.: | Markhaseva, 2012 (p.296); Markhaseva & al., 2014 (p.82, Table 1, 2, 3, 4, Rem.) ; Renz & Markhaseva, 2015 (p.96, Table 4, Fig.3, sex ratio, biogeography); Laakmann & al., 2019 (p.330, Table 1) | | Rem.: | Type species: Xancithrix ohmani Markhaseva, 2012. Total: 1 sp.
Diagnosis from Markhaseva (2012, p.296): Female:
- Cephalosome and pediger 1, pedigers 4 and 5 partly.
- Posterior corners of prosome as short obtuse-triangular lobes.
- Rostrum absent.
- A1 24-segmented.
- Coxa of A2 with 1 seta; basis with 2 setae; endopodal segment 1 with 2 setae; exopodal proximal segments 1 and 2 without setae.
- Md gnathobae cutting edge with 10 teeth ; rndopod al segments 1 and 2 with 2 and 9 setae, respectively.
- Mx1 praecoxal arthrite with 14 setal elements; coxal endite with 5 setae; proximal basal endite with 4 setae; distal basal endite with 5 setae; endopod with 11 setae; exopod with 10 setae; coxal epipodite with 9 setae.
- Mx2 praecoxal endite bearing 5 setae; proximal basal endite with 2 sclerotized plus 1 sensory setae; lobe of proximal endopodal segment with 2 sclerotized plus 2 sensory setae; endopod with 3 worm-like, 5 brush-like and 1 sclerotized setae.
- Mxp syncoxa with 1, 2, and 3 setae on praecoxal endites; coxal endite with 3 setae, small attenuation and spinules along distal edge.
- Segmentation and setation of P1-P4 typical for Clausocalanoidea.
- P5 uniramous, 3-segmented,; exopodal segment 3 with 3 terminal spines.
Male:
- Cephalosome and pediger 1 nearly completely fused, pedigers 4 and 5 completely fused.
- Posterior corners of prosome in form of short obtuse lobes.
- Rostrum rudimentary with small terminal motch.
- Right A1 ,of 23, left of 24 segments.
- Oral parts rudimentary compared to female.
- Lateral spines of P1 exopodal segments 1 to 3, and P2-P4 exopodal segment 1 shorter than in female.
- P5 biramous, rndopods rudimentary; right protopod shorter than left; exopods 3-segmented.
For Markhaseva this new genus is considered to be one of the most primitive genera among the bradfordian families and retains the following ancestral bradfordian setal pattern of oral parts: 1- a setal sequence of 1, 2, 3 on the praecoxal endites of the Mxp syncoxa (shared with the Tharybidae and Diaixidae); 2- the Mx2 praecoxal endite and endopod with 5 and 9 setae respectively (shared with diaixid genera Procenognatha, Xantharus, Landrumius, Falsilandrumius, neoscolecithrix, Paraxantharus, Grievella, and some phaennids of Xanthocalanus, Brachycalanus); 3- a setal sequence of 14, 5, 4, 5, 11, 10, and 9, respectively, on the Mx1 praecoxal arthrite, coxal endite, proximal basal endite, distal basal endite, endopod, exopod and coxal epipodite (shared with some phaennid species of Xanthocalanus).
The new genus belongs to the lineage containing ancestral bradfordian families Diaixidae and Tharybidae. It is closer to the Diaixidae in having more than 6 setae on the Mx1 exopod (vs 2-6 setae in tharybids) and in the trapezoidal shape of the Mx1 praecoxal arthrite not vaulted (vs vaulted in Tharybis).
The new genus is distinguished from the other members of Diaixidae by the combination of derived characters: rostrum absent in females, Mxp syncoxa with small distal attenuation on the coxal endite and P1 endopod bearing lateral lobe with pore. | | Remarks on dimensions and sex ratio: | | Total body length for 3 females: 4.50-4.55 mm; only one male 4.6 mm. | | | | | | | | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 27, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|



;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
