|
|
 |
|
Calanoida ( Order ) |
|
|
|
Clausocalanoidea ( Superfamily ) |
|
|
| |
| | | |
| Aetideidae Giesbrecht, 1892 ( Clausocalanoidea ) | | Syn.: | Aëtidiina Giesbrecht,1892 (p.52); Aetidiinae : Esterly, 1905 (p.143) | | Ref.: | Sars, 1901 a (1903) (p.23); With, 1915 (p.72); Gurney, 1931 a (p.84); Rose, 1933 a (p.88); Brodsky, 1950 (1967) (p.82, 141); Vervoort, 1952 (n°41, p.2, Genera Key); Mazza, 1967 (p.106); 1968 (p.531); Bradford, 1969 b (p.473, 502: Table 3): Björnberg, 1972 (p.25); Andronov, 1974 a (p.1005); Campaner, 1978 (p.865); Bradford & Jillett, 1980 (p.11); Vaupel-Klein, 1982 a (p.1); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.153); 1993 (p.309); Brodsky & al., 1983 (p.144, 145, 147); Zheng Zhong & al., 1984 (1989) (p.235, genera Key); Mauchline, 1988 (p.426, 731, 740: cuticular pores); Huys & Boxshall, 1991 (p.460); Markhaseva, 1993 (p.51, phylogeny); Vyshkvartzeva, 1994 (p.118 & suiv.); Markhaseva, 1996 (p.3, Genera Key, Rem.: p.8); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.681, Genera Key); Bradford-Grieve & al., 1999 (p.879, 903, 904, 917: Genera & spp. Key); Ohtsuka & Huys, 2001 (p.445, 461); Markhaseva & Schnack-Schiel, 2003 (p.116 & suiv., Rem.); Boxshall & Halsey, 2004 (p.12, 15, 16; 49; p. 54: Def.; p.57: Genera Key); Vives & Shmeleva, 2007 (p.533, part. Genera Key); Markhaseva & Schulz, 2008 (p.48, Fig.12: geographical distribution of benthopelagic genera); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, molecular biology, phylogeny); Laakmann & al., 2012 (p.535, Rem.: p.543, phylogeny); Markhaseva & Renz, 2015 (p.1041, Rem.: differences between Clausocalanidae and Aetideidae); Laakmann & al., 2019 (p.330, fig.1, 2, 3, Table A, phylogenetic relationships); Hirai & al., 2020 (p.1, Fig.4: metabarcoding, Fig.8: OTUs distribution patterns, Fig.9: phylogenetic analysis)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | Type-genus: Aetideus Brady, 1883. Total: 30 G.: Aetideopsis, Aetideus, Azygokeras, Batheuchaeta, Bradyetes, Bradyidius, Chiridiella, Chiridius, Chirundina, Chirundinella, Comantenna, Crassantenna, Euchirella, Gaetanus, Gaidiopsis, ['Gaidius'= Gaetanus], Jaschnovia, Lutamator, Mesocomantenna, Paivella, Parabradyidius, Paracomantenna, Prolutamator, Pseudeuchaeta, Pseudochirella, Pseudotharybis, Pterochirella, Sursamucro, Undeuchaeta, Valdiviella.
Senecella represents the only brackish- and freshwater genus.
For Markhaseva (1996, p.8) this family comprises only 26 genera (with the new genus Parabradyidius); Gaidius is synonym with Gaetanus; Wilsonidius is considered as a synonym of Chirundina; Gaidiopsis is insufficiently described and the type species lost; Pseudotharybis is intermediate between this family and the Tharybidae; Valdiviella could constitute the type of a new Family.
Diagnosis from Boxshall & Halsey (2004, p.54) :
- Cephalosome and pedigerous somite 1 fused, or rarely with partial suture dorsally, pedigerous somites 4 and 5 sometimes separate.
- Rostrum variable, may be simple, bifid or divided into paired rostral filaments, rarely absent.
- Anterior part of cephalosome may form crest.
- Frontal cephalic spine present in some species of Gaetanus.
- Nauplius eye present.
- Last prosomal somite wometimes rounded, usually with pointed and posteriorly produced epimeral lobes.
- Urosome 4-segmented in female ; with 3 free abdominal somites.
Female genital double-somite symmetrical or asymmetrical, with common genital aperture located medially on ventral surface ; copulatory pore (s) contained within median genital aperture.
- Urosome 5-segmented in male ; single genital aperture located ventrolaterally at posterior rim of genital somite (1st urosomal segment) on left side.
- A1 23 to 25-segmented in female ; often longer than prosome ; segment 2 triple (II-IV), segments 8 (X) and 9 (XI) typically fused, segments 24 (XXVI) and 25 (XXVII-XXVIII) fused or separate. Aesthetascs typically present on segments III, VII, XI, XIV, XVI, XXI, XXVIII in female. A1 male typically non geniculate ; additional segmental fusions may include X-XI and XII , XV and XVI or XXIII and XXIV. Aesthetascs present on all segments dxcept I, XXII and XXV (based on Euchirella rostrata). Right A1 of male Azygokeras with 6 distal segments enlarged, possibly geniculate but mechanism may not be homologous with the neocopepoden geniculation.
- A2 biramous with separate coxa and basis ; coxa with 1 seta ; basis with 1 or 2 setae ; endopod 2-segmented, distal segment bilobed wuth maximum setation formula 2, 9+7, often reduced ; exopod usually larger than endopod, 7-segmented with compound segments I-II, III-IV and IX-X ; basic setation formula 2, 2, 1, 1, 1, 1,3, but setation sometimes reduced and additional segmental fusions sometimes present.
- Md biramous, coxa with well developed gnathobase and distal palp consisting of basis, 2-segmented ebndopod and 5-segmented exopod ; basis with 0 to 4 setae, usually 2 or 3 ; endopodal segments 1 and 2 wiyj up to 3 and 9 setae ; exopodal setation formula 1, 1, 1, 1, 2, setation often reduced.
- Mx1 with well developed praecoxal arthrite bearing 14 elements (9 distal spines, 4 posterior surface setae and 1 proximal seta) ; coxa with endite bearing 1 to 5, usually 4, setae and with 9 setae on epipodite ; basis without oiter seta, with small proximal endite bearing up to 5, usually 3 or 4 setae and distal group of 3 to 5 setae representing distal enditr ; endopod sometimes fused to basis, unsegmentrd to 3-segmented ; setation formula 4, 4, 7 ; exopod 1-segmented, usually armed with 9 to 11 setae ; maxillulary setation often very reduced as in Chiridiella/
- Mx2 7-segmented ; praecoxa and coxa separate, setation formula of endites 3, 3, 3, 3 ; basis with 3 setae including large claw-like element fused to basis ; free endopod 4-segmented, usually with 6 major setae but full setation formula 2, 3, 3, 3 ; maxilla highly modified in some Chiridiella species, carrying 2 or 3 strong, curved apical claws.
- Mxp 7-segmented, with distal part notated at synncoxa-basis joint ; syncoxa with endite setation formula 1, 2, 3, 4 ; most proximal seta sometimes absent ; 1 distal endite element sometimes modified, aesthetasc-like ; basis with 3 setae, plus 2 setae on incorporated 1st endopodal segment ; free endopod 5-segmented, segmental setation formula 4, 3, 3, 3+1, 4 ; setation often reduced ; syncoxa bearing large blade-like lateral pprocess in many Gaetanus species ; basis with strongly sclerotized outer margin in Chirundina.
- Male mouthparts often reduced compared to female.
- Swimming legs P1 to P4 biramous, typically with 3-segmented rami, except P1 with 1-segmented endoipod and P2 with 1 or 2-segmented endopod. P1 exopod 1-segmented in Chiridiella or 2-segmented in some species of Gaetanus and Euchirella ; additional segmental fusions sometimes present, as between proximal exopodal segmenys of Valdiviella species.
- Coxa of P4 ornamented with cluster of spinules near base of inner seta ; with 2 transverse rows of 4 to 6 spinules in Paivella, but usually lacking surface ornamentation except for scattered spinules.
Key to genera after Boxshall & Halsey (2004, p.57) ( modified from Markhaseva, 1996) :
Females:
1 5th pedigerous somite and genital double-somite symmetrical
. 2.
1 Either 5th pedigerous somite or genital double-somite, or both, more or less asymmetrical
27.
2 Rostrum present, not reduced
. 3.
2 Rostrum absent or present as a reduced semi-circular plate
.. 15.
3 Rostrum bifurcate
4.
3 Rostrum conical, tapering to single point
9.
4 Swimming legs typically with some degree of fusion between proximal segments of each ramus ; maxillulary endopod with at most 3 setae
Valdiviella.
4 Swimming legs without fusion of proximal segments ; maxillulary endopod with more than 3 setae
5.
5 P1 lacking outer spine on 1st exopodal segment ; Mx1 with 11 setae on exopod, and 3 setae on coxal endite
. 6.
5 P1 with outer spine on 1st exopodal segment ; Mx1 with 8 to 11 setae on exopod, 4 or 5 setae on coxal endite
7.
6 Coxa of P4 ornamented with 2 parallel rows of 4 to 6 spinules on posterior surface near base of inner seta
. Paivella.
6 Coxa of P4 lacking such ornamentation on posterior surface
.. Aetideus.
7 P5 3-segmented, distal segment with 3 spinous projections
31.
7 P5 vestigial or absent
. 8.
8 Outer margin spine on 2nd exopodal segment of P1 thicker than that on 1st segment ; P2 endopod always 2-segmented
.Bradyidius.
8 Outer margin spine on 2nd exopodal segment of P1 not markedly thicker than that on 1st segment ; P2 endopod either 1 or indistinctly 2-segmented
. Aetideopsis.
9 Coxa of P4 ornamented with spinules on posterior surface near base of inner seta
11.
9 Coxa of P4 lacking such ornamentation of spinules near base of inner seta
.. 10.
10 Mxp with conspicuous sclerotized thickening extending along outer margin of basis ; Mx1 with 11 setae on exopod, 15 on endopod and 5 on coxal endite
. Chirundina.
10 Mxp without conspicuous sclerotized thickening on basis ; Mx1 with 10 setae on exopod, 11 on endopod and 4 on coxal endite
Chirundinella.
11 A2 endopod at most half as long as exopod ; Md basis unarmed ; Mx1 endopod with at most 5 setae
Euchirella (part).
11 A2 endopod more than half as long as exopod ; Md basis armed with 1 or more setae ; Mx1 endopod with at least 14 setae
. 12.
12 Frontal margin of cephalosome with long spinous process
Gaetanus (part).
12 Frontal spinous process absent
.. 13.
13 Mxp with blade-like process on lateral margin of syncoxa
.Gaetanus (part).
13 Mxp lacking outer blade-like process on syncoxa
. 14.
14 4th and 5th pedigerous somites fused ; P1 exopod with 2 outer spines ; Md basis with ½ setae
.. Gaetanus (part).
14 4th and 5th pedigerous somites usually sepatate ; P1 exopod with 3 outer spines ; Md basis with 3 setae
. Pseudochirella (part).
15 Distal syncoxal endite of Mxp with 1 modified, blunt-ending sensory element
23.
15 Distal syncoxal endite of Mxp lacking any modified, blunt-ending element
16.
16 P1 exopod 1-segmented ; Mx2 often highly specialised, armed with strong, curved apical claws
.Chiridiella.
16 P1 exopod 3-segmented ; Mx2 lacking specialised apical claws
17.
17 1st exopodal segment of P1 lacking outer spine
Jaschnovia.
17 - 1st exopodal segment of P1 armed with outer spine
.. 18.
18 3rd exopodal segment of P2 to P4 with 3 outer spines
19.
18 3rd exopodal segment of P2 to P4 with only 2 outer spines
.. Senecella calanoides.
19 Posterolateral angles of prosome drawn out into conspicuous points or spinous processes
. 20.
19 Posterolateral angles of prosome without conspicuous points or spinous processes
. 22.
20 Posterolateral prosomal processes directed straight backwards ; 4th and 5th pedigerous somites fused ; rostrum absent
Chiridius.
20 Posterolateral prosomal processes curved backwards ; 4th and 5th pedigerous somites separate ; rostrum reduced, in form of semi-circular blade
21.
21 2nd segment of Md endopod with 4 setae, basis unarmed ; A2 endopod better developed than exopod
. Crassantenna.
21 - 2nd segment of Md endopod with 9 setae, basis with 2 setae ; A2 endopod smaller than exopod
. Sursamucro.
22 Rostrum absent ; 2nd endopodal segment of Md with 10 setae, 1st endopodal segment with 2 setae, basis with 3 setae
.. Azygokeras.
22 Rostrum present in form of reduced semi-circular blade ; 2nd endopodal segment of Md with 4 or 5 setae, 1st endopodal segment and basis unarmed
. Lutamator.
23 A2 exopod shorter than endopod
. 24.
23 A2 exopod as long as, or longer than, endopod
. 26.
24 Distal segment of A2 exopod shorter than preceding 2 segments combined ; 1 or more of terminal antennary exopodal setae much shorter than others, or only 1 seta present
.. 25.
24 Distal segment of A2 exopod longer than preceding 2 segments combined ; all terlinal antennary exopodal setae similar in length
. Paracomantenna.
25 Distal segment of A2 exopod armed with 3 setae
. Comantenna.
25 Distal segment of A2 exopod armed with single apical seta
..Mesocomantenna.
26 1st exopodal segment of P1 with outer spine present, not reduced ; setae on free endopodal segments of Mxp ornamented with modified crescent-like spinules ; setae on syncoxal and basal endites, and on endopod of Mx2 sabre-like, not transformed into claw
Pseudeuchaeta.
26 1st exiopodal segment of P1 without outer spine, or outer spine reduced ; setae on free endopodal segments of Mxp without crescent-like spinules ; setae on syncoxal and basal endites, and on endopod of Mx2 not sabre-like , at least 1 seta on basal endite trnsformed into claw
Bradyetes.
27 Rostrum absent or vestigial
. 28.
27 Rostrum oresent, well developed
29.
28 Md basis with 2 setae ; distal syncoxal endite of Mxp with 3 setae only, lacking modified element
. Senecella siberica.
28 Md basis with 3 setae ; distal syncoxal endite of Mxp with 3 setae plus 1 modified, blunt-ending sensory element
. Batheuchaeta.
29 Coxa of P4 ornamented with spinules on posterior surface near base of inner seta ; exopod of P1 armed with 3 outer margin spines
30 .
29 Coxa of P4 ornamented without spinules on posterior surface near base of inner seta ; exopod of P1 armed with 2 outer margin spines
Undeuchaeta.
30 Exopod of P1 2-segmented ; Md basis lacking setae, 1st endopodal segment unarmed or 1 small seta
.. Euchirella (part).
30 Exopod of P1 3-segmented ; Md basis with 3 setae, 1st endopodal segment with 2 or 3 setae
.. Pseudochirella (part).
31 Md basis with 2 setae ; rami of A2 subequal in length, or exopod slightly shorter than endopod
. Parabradyidius.
31 Md basis with 3 setae ; exopod of A2 distinctly longer than endopod
. Pseudotharybis.
Males :
1 Rostrum present
.. 9.
1 - Rostrum absent
2.
2 Right A1 apparently geniculate, with distal 6 segments enlarged
Azygokeras.
2 Both A1 non-geniculate, without enlarged segments distally
. 3.
3 P56 present on left side only, uniramous
Aetideus.
3 P5 present on both sides ; legs P5 uniramous or biramous
.. 4.
4 Legs P5 uniramous, left leg 5-segmented, right leg 4-segmented
Chiridius.
4 Legs P5 biramous, left exopod 3-segmented, right exopod 2 or 3-segmented
. 5.
5 Exopod of right P5 3-segmented
. 8.
5 Exopod of right P5 2-segmented
6.
6 3rd exopodal segment of left P5 stylet-like distally ; left endopod segmented, not more than 1/3 as long as 1st exopodal segment of left leg
Jaschnovia.
6 3rd exopodal segment of left P5 not stylet-like distally ; left endopod typically 2-segmented, or if 1-segmented then longer than 1st exopodal segment
. 7.
7 2nd exopodal segment of right P5 filament-like distally, ornamented with long hair-like setules
Comantenna.
7 2nd exopodal segment of right P5 not filament-like distally, lacking hair-like ornamentation
.. Bradyetes.
8 Dorsal cephalic shield with marked transverse cleft about at level of mandibular somite and with paired posterodorsal wing-like projections
. Pterochirella.
8 Dorsal cephalic shield lacking conspicuous transverse cleft and dorsal projections
Chiridiella.
9 Rostrum bifurcate
.. 10.
9 Rostrum conical, tapering to single point
17.
10 Posterolateral angles of prosome rounded
.. 11.
10 Posterolateral angles of prosome tapering to spinous processes or with tiny spinous tooth
.. 12.
11 Legs P5 uniramous ; left leg elongate, right leg short
Paivella.
11 Legs P5 biramous with 3-segmented exopods and 1-segmented endopods
. Valdiviella.
12 Legs P5 uniramous or biramous ; posterolateral angles of prosome forming short spinous processes or tooth not reaching to posterior margin of genital somite
.. 13.
12 - Legs P5 biramous ; posterolateral angles of prosome with well developed spinous processes extending posteriorly to posterior margin of genital somite
16.
13 Legs P5 with 3-segmented exopods and indistinctly 2-segmented endopods
. Parabradyidius.
13 Legs P5 with at least one exopod 2-segmented and one endopod 1-segmented
.. 14.
14 Legs P5 with 3-segmented left exopod
. 15.
14 Legs P5 with 2-segmented left exopod and 3-segmented right exopod
. Bradyidius (part).
15 Rostrum bifurcate with divergent branches
Pseudotharybis.
15 Rostrum bifurcate with closely adpressed, parallel branches
. Aetideopsis (part).
16 Legs P5 with 2-segmented right exopod and 3-segmented left exopod
Aetideopsis (part).
16 legs P5 with 3-segmented right exopod and 2-segmented left exopod
. Bradyidius (part).
17 Basis of right P5 swollen, almost circular in outline ; left exopod complex with swollen irregular segments, left endopod irregular, tapering proximally
Senecella ;
17 legs P5 not of this form, basis and ramal segments typically cylindrical
.. 18.
18 Rostrum rudimentary ; exopod of right P5 3-segmented
. Pseudeuchaeta.
18 Rostrum not rudimentary ; exopod of right P5 2-segmented
.. 19.
19 Frontal margin of cephalosome with spinous process
. Gaetanus (part).
19 Frontal margin of cephalosome without spinous process
. 20.
20 basis of left P5 with 2 processes on inner margin
Batheuchaeta.
20 Basis of left P5 without processes on inner margin
21.
21 Right P5 with elongate endopod and 2nd exopodal segment forming chela-like mechanism
Euchirella (part).
21 Right P5 not forming chela-like mechanism
. 22 ;
22 Dorsal cephalic shield with crest
.. 23.
22' - Dorsal cephalic shield without crest
. 25.
23 P1 exopod 2-segmented
... Euchirella (part).
23' - P1 exopod 3-segmented
.. 24.
24 3rd exopodal segment of left P5 bilobed ; 1st exopodal segment of P1 with outer spine
.. Chirundinella.
24 3rd exopodal segment of left P5 not stylet-like distally, more triangular in outline ; 1st exopodal segment of P1 with or without outer spine
.. Chirundina.
24 3rd exopodal segment of left P5 stylet-like distally ; 1st exopodal segment of P1 without outer margin spine
. Undeuchaeta (part).
25 Left P5 lacking teeth on 2nd exopodal segment ; without processes on other segments of both rami, or with general complex configuration
26.
26 P1 exopod 2-segmented
Euchirella (part).
26 P1 exopod 3-segmented
. 27.
27 1st exopodal segment of P1 with outer spine
Pseudichirella.
27 1st exopodal segment of P1 without outer spine
Undeuchaeta (part).
For Schulz & Markhaseva (2000, p.87) the genus Parabradyidius differs from Aetideopsis, Bradyidius, Pseudotharybis in relative length of A2 endo- and exopod: while in these three genera the exopod is distinctly longer than the endopod (ca. 1.2 - 1.5 times), in Parabradyidius these rami are subequal or the exopod may be even slightly shoryter. In addition, Parabradyidius lacks the medial seta of the A2 7th exopodal segment present in these three taxa.
The male of the new genus is the only aetideid male known so far to have both exopods of P5 equipped with 3-segmented rami as well as, indistinctly , 2-segmented endopods. Such segmentation can be considered the most primitive among Aetideidae. Only Azygoceras, Chiridiella, Pseudeuchaeta share 3-segmented exopods on both rami, while P5 endopods are 1-segmented in these three genera. 2-segmented left endopod is typical only of the aetideids Bradyetes matthei, Bradyidius arnoldi, Comantenna brevicornis. All remaining aetideids share a 1-segmented endopod (if present at all) in the male. However, in Parabradyidius angelikae exopodal segment 3 of the left ramus is much more specialized than that in all other Aetideidae in that it has a deep excavation furnished with short hairs at the medial inner part of the segment, opposed to a segment of cylindrical shape lacking any excavation in remaining taxa. This combination of plesiomorphic as well as apomorphic characters appears to be a general feature of benthopelagic copepod taxa.
|  issued from : E.L. Markhaseva & K. Schulz in Mitt. hamb. zool. Mus. Inst., 2008, 105. [p.49, Fig.12]. Geographical distribution of benthopelagic Aetideidae in the Atlantic Ocean including Polar regions (Markhaseva, 1996, 1997; Markhaseva & Schnack-Schiel, 2003; Markhaseva & Schilz, 2006; Schulz, 2002; Schulz & Markhaseva, 2000, and unpublished original data). |
 issued from : E.L. Markhaseva & V.YU. Razzhivin in Okeanol., 1984, 29 (3). [p.612, Gig.1]. Vertical distribution of the Aetideidae in the Kuril-Kamchatka trench region. Thickness of bars indicates frequency of occurrence (percentage of total number of collections in the depth interval in question); Numbering of the species in circle: 1- Aetideus pacificus; 2- Aetideopsis multiserrata; 3- A. rostrata; 4- A. retusa**; 5- Batheuchaeta animala**; 6- B. gurjanovae; 7- B. hepneri**; 8- B. lamellata; 9- Pseudeuchaeta spinata**; 10- Batheuchaeta peculiaris**; 11- Chiridius pacificus; 12- C. polaris; 13- Chiridiella abyssalis; 14- Ch. bichela**; 15- Ch. gibba**; 16- Ch. pacifica; 17, Ch. smoki**; 18- Ch. subaequalis**; 19- Euchirella curticauda; 20- E. formosa; 21- Gaetanus intermedius; 22- G. paracurvicornis; 23- G. brevispinus; 24- G. brevirostris; 25- G. inermis*; 26- G. robustus; 27- G. tenuispinus; 28- G. minutus; 29- Pseudochirella accepta; 30- P. batillipa*; 31- P. dubia; 32- P. pacifica; 33- P. palliata**; 34- P. obtusa; 35- P. tanakai**. The species not previously identified in the region are marked by a siigle asterisk and those not previously reported in the Pacific by a double asterisk. |
 issued from : E.L. Markhaseva & F.D. Ferrari in Invert. Zool., 2005, 2 (2). [p.162, Table 4] Setation of oral parts in females Aetideidae (Clausocalanoidea) and ancestral condition of setation. |
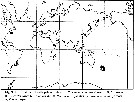 issued from : E.L. Markhaseva in Pelagic Biogeography ICoPB II. Proceedings of the 2nd International Conference. Final report of SCOR/IOC working group 93. Noordwijkerhout, The Netherlands 9-14 July 1995. Workshop Report No.142. UNESCO, 1998. [p.252, Fig.3] Distribution of benthopelagic aetideids in the World Ocean. |
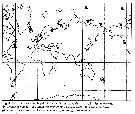 issued from : E.L. Markhaseva in Pelagic Biogeography ICoPB II. Proceedings of the 2nd International Conference. Final report of SCOR/IOC working group 93. Noordwijkerhout, The Netherlands 9-14 July 1995. Workshop Report No.142. UNESCO, 1998. [p.253, Fig.4] Distribution of benthopelagic aetideids in the World Ocean. |
 Issued from : E.L. Markhaseva in Marine Plankton II, Explorations of the Fauna of the Seas, 1993, 45 (53). [p.57, Table I] Phylogenetic trends within Aetideidae (Copepoda, Calanoida). Setal armature of A2. A: exopodal segment 1; B: exopodal segment 2; C: medial seta on the exopodal segment 7; Re/Ri: Exopod length/endopodal length ratio. |
 Issued from : E.L. Markhaseva in Marine Plankton II, Explorations of the Fauna of the Seas, 1993, 45 (53). [p.58, Table 2] Phylogenetic trends within Aetideidae (Copepoda, Calanoida). Setal armature of mandibular palp. B2: basipodite 2; Ri: endopod. |
 Issued from : E.L. Markhaseva in Marine Plankton II, Explorations of the Fauna of the Seas, 1993, 45 (53). [p.59, Table 3] Phylogenetic trends within Aetideidae (Copepoda, Calanoida). Setal armature of Mx1. Gnathobase: inner lobe 1; B2: distal part of inner side of Ri |
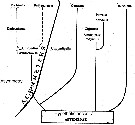 Issued from : E.L. Markhaseva in Marine Plankton II, Explorations of the Fauna of the Seas, 1993, 45 (53). [p.67, Fig.4] Phylogenetic trends within Aetideidae (Copepoda, Calanoida). Ac..... : asymmetry. Three main phylogenetic trends within Aetideidae are revealed: The first is retraced for Aetideus, Paivella, Aetideopsis, Badyidius, Chiridius. Among them Aetideus and Paivella are considered as most specialized; the second; Gaetanus, Pseudochirella, Chirundina, Chirundinella, Undeuchaeta, Euchirella and Batheuchaeta, the later: two genera are the most specialized; the third trend of Aetideidae evolution is represented by Chiridiella which may be a good example of feeding specialization. Epibenthic aetideids show weak resemblance with pelagic ones and are not referred to any of the three mentioned trends, but altogether might be characterized by having in their morphology a mosaic combination of primitive and progressive features. Aetideidae developed by expansion of nearly all pelagic vertical zones and by broadening of feeding types from the mixed type of feeding to predation ( Pseudeuchaeta) or sucking predation ( Chiridiella). |
 issued from : E.L. Markhaseva in Pelagic Biogeography ICoPB II. Proceedings of the 2nd International Conference. Final report of SCOR/IOC working group 93. Noordwijkerhout, The Netherlands 9-14 July 1995. Workshop Report No.142. UNESCO, 1998. [p.252, Fig.2] Distribution of aetideids in the World Ocean. |
 issued from : E.L. Markhaseva in Pelagic Biogeography ICoPB II. Proceedings of the 2nd International Conference. Final report of SCOR/IOC working group 93. Noordwijkerhout, The Netherlands 9-14 July 1995. Workshop Report No.142. UNESCO, 1998. [p.253, Fig.5] Graph of the similarity of total aetideid fauna (pelagic and bentho-pelagic)). |
 issued from : E.L. Markhaseva in Pelagic Biogeography ICoPB II. Proceedings of the 2nd International Conference. Final report of SCOR/IOC working group 93. Noordwijkerhout, The Netherlands 9-14 July 1995. Workshop Report No.142. UNESCO, 1998. [p.251, Fig.1] Latitudinal zones of the World Ocean pelagial. The geographical subdivision of the pelagic is given according to Nesis, 1982 (in Zoogeography of the World Ocean: comparison of the pelagic zonality and the shelf regional subdivision. In: O.G. Kussakin ed. Marine Biogeography. Nauka, Moskva: 114-134). The similarity of faunas in the different geographical zones of the world Ocean was examined with the use of Sorensen's index of similarity (IS) and Simpson's index of inclusion (II) of less diverse fauna into the richer one. Four types of aetideid copepod distributions are denoted: 1 - Cosmopolitan genera: 7 genera, of which Euchirella is only absent in the Arctic Ocean. The mainly pelagic genera: Gaetanus, Pseudochirella, Euchirella, Aetideopsis, Chiridiella, Chiridius, Pseudeuchaeta. 2 - Tropico-boreal genera: Three pelagic genera Aetideus, Chirundina, Undeuchaeta and two benthopelacic genera Bradyidius, Comantenna. 3 - Genera found within tropical limits: Two pelagic genera Chirundinella, Paivella and five benthopelagic genera Crassantenna, Lutamator, Mesocomantenna, Paracomantenna, Sursamucro. 4 - Benthopelagic genera: Three benthopelagic genera are registered within the limits of the Arcto-boreal Jaschnovia is found in both arctic and boreal zones; Bradyetes is known from the boreal Atlantic and Azygokeras from the boreal Pacific. The pelagic genera demonstrate wide distributions, whereas 8 benthopelagic genera are mostly endemic. Comparison of faunas: The tropical Pacific aetideid fauna is the richest with 98 species, followed by the tropical Atlantic fauna with 73 species, and the tropical Indian Ocean fauna with 50 species. nearly equal in species number are the boreal Atlantic (57 species) and boreal Pacific (53 species) faunas. The Subantarctic fauna (Not) includes 30 species and the Antarctic and Arctic faunas contain 29 and 14 species respectively. The results of the analyses of the total aetideid fauna show a high degree of similarity between tropicalAtlantic, tropical Pacific and tropical Indian ocan faunas (IS = 56 to 62 %) and between each of these fauna with fauna of boreal Atlantic (IS = 54 to 58 %) The degree of similarity between Subantarctic and Antarctic faunas is also high (IS = 61 %). An aetideid fauna of the upper 500 m layer is nearly absent in Polar basins (see fig.6). Few aetideid species occur in the upper layers in Polar basins. Nevertheless they are typical of deeper layers in the other regions of the Wiorld Ocean and it are most likely meso-, bathy-, or meso-/bathypelagic species. The tropical faunas are significantly similar to each other (IS = 63 to 77 %) and to the boreal Atlantic fauna (IS = 67 to 78 %). The latter is also similar to the Subantarctic fauna (IS = 57 %). The faunas of the tropical Atlantic and Indian Oceans are related to the tropical Pacific fauna (for the tropical Atlantic II = 81 %, the latter includes the total fauna) and the boreal Atlantic fauna is related to the tropical faunas with II values of 73 % to 93 %. The fauna of the Subantarctic is totally included into the boreal Atlantic fauna. The boreal Pacific fauna is poor in comparison to tropical faunas , and in this respect it is separated more clearly than the boreal Atlantic fauna. The similarity between the boreal Pacific and boreal Atlantic faunas is weak; and similarity is even smaller between boreal Pacific and Subantarctic zones. The deep water aetideid fauna exists in Polar basins (10 species in Arctic and 22 species in Antarctic zones (see fig.7). The similarity between these faunas is moderate (IS = 38 %), and due to the presence of the bipolar species ( Aetideopsis minor, Pseudochirella batillipa, P. spectabilis), and the cosmopolitan species Gaetanus brevispinus, G. tenuispinus. The arctic fauna is poor and has weak similarity to boreal and tropical faunas (IS = 21 to 24 %) The boreal faunas of deep water aetideids are rather similar (IS = 52 %). The boreal Atlantic deep water fauna is related to the boreal Pacific fauna. It is very possible that boreal faunas originated from tropical faunas, but the boreal Pacific fauna probably is older than the boreal Atlantic. The boreal Pacific deep water fauna has higher similarity with the deep water aetideid fauna of the tropical Pacific (IS = 51 %) than with the upper layer aetideid faunas. The similarity of the boreal Pacific fauna with the tropical Atlantic and Indian Ocean faunas, is weak and this is comparable with the above mentioned similarity for the upper layer aetideids. The distribution of deep water aetideids showed that they have significantly more similarities than the faunas of the upper layers. The fact that the tropical Pacific fauna is the richest suggests that it is the most ancient among aetideid fauna. |
 issued from : E.L. Markhaseva in Pelagic Biogeography ICoPB II. Proceedings of the 2nd International Conference. Final report of SCOR/IOC working group 93. Noordwijkerhout, The Netherlands 9-14 July 1995. Workshop Report No.142. UNESCO, 1998. [p.254, Figs.6, 7] Graphs of the similarity. of aetideids. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.54]. Armature formula of swimming legs P1 to P4. Nota: Swimming legs P1 to P4 biramous, typically with 3-segmented rami, except P1 with 1-segmented endopod and P2 with 1 or 2-segmented endopod. -P1 exopod 1-segmented in Chiridiella or 2-segmented in some species of Gaetanus and Euchirella; additional segmental fusions sometimes present, as between proximal exopodal segments of valdiviella species. - Coxa of P4 ornamented with cluster of spinules near base of inner seta; with 2 transverse rows of 4 to 6 spinules in Paivella, but usually lacking surface ornamentation except for scattered spinules. Inner seta on basis of P1 situated on anterior surface of basis and passing across face of 1st endopodal segment. Seration sometimes reduced or modified, only 2 outer spines on distal exopodal segments of P2 to P4 in Senecella calanoides Juday. Terminal spines on exopods of P2 to P4 with conspicuous dentate outer margin. - P5 usually absent in females, rarely represented by biramous vestige, as in Comantenna recurvata Grice & Hulsemann, or by segmented vestige as in Pseudotharybis, Parabradyidius and some specimens of Sursamucro and Aetideopsis. - Male P5 typically asymmetrical; often elongate, with styliform distal segments on exopods. Right leg typically biramous with up to 3-segmented exopod and 1-segmented endopod, rarely absent as in Aetideus. Left leg biramous with up to 3-segmented exopod and 1-segmented endopod. P5 highly variable; both may be uniramous, lacking endopods. Endopods rarely 2-segmented as in Parabradyidius angelikae Schulz & Markhaseva, Bradyetes matthei Johnnessen, Bradyidius arnoldi Fleminger and Comantenna brevicornis (Boeck). - Eggs retained in paired multiseriate sacs, at least in some species; rarely in uniseriate sacs as in Euchirella pseudopulchra Park. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.55, Fig.3]. Aetideidae. A, Euchirella messinensis habitus female; B, A2; C, Mx2; D, Mxp; E, Pseudochirella pustulifera protopod* of P4. [von Vaupel Klein, 1982b: A-D; Sars, 1924: E]. *: protopod = the basal part of a limb (praecoxa+coxa+ basis) that carries the rami. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.59, Fig.4]. Aetideidae. A, Gaetanus pileayus habitus female; B, Chiridiella macrodactyka Mx2; C, Comantenna brevicornis (as Bryaxis brevicornis) Mxp; D, Pseudotharybis brevispinus female P5; E, Ubdeuchaeta incisa male P5; F, Chiridius subgracilis male P5. [Sars, 1924: A-B; Sars, 1902: C; Bradford & Jillett, 1980: D-F]. |
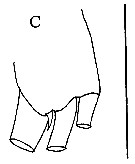 Issued from : E.L. Markhaseva & J. Renz in Crustaceana, 2015, 88 (9). [p.1043, Fig.6, C]. Aetideopsis armata. Female: C, Mx2 praecoxal endite. |
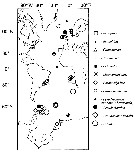 Issued from : E.L. Markhaseva & K. Schulz in Mitt. hamb. zool. Mus. Inst., 2008, 105. [p.49, Fig.2]. Geographical distribution of benthopelagic Aetideidae in the Atlantic Ocean including Polar regions (Markhaseva, 1996, 1997; Markhaseva & Schnack-Schiel, 2003; Markhaseva & Schulz, 2006; Schulz, 2002; Schulz & Markhaseva, 2000; Markhaseva & Schulz, 2008, and unpublished original data). |
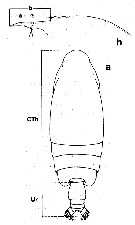 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.59, Fig.3, a, h]. a, measurements taken of cephalosome (CTh) and urosome (Ur) to compare CTh/Ur length ratio; h, two possibilities to determine the length of the rostrum. Characters used in Table I for cluster analysis. |
 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.87, Table I]. Characters as defined for use in cluster analysis. Symbols used (coding of states): - CTh: cephalosome. - Ur : urosomites 1 to 4. - A1: antennule. - A2: antenna; - Md: mandible; - Mx1: maxillule. - Max: Mx2, maxilla; - Mxp: maxilliped. - P1 to P4: swimming legs 1 to 4. Ri: endopod. Re: exopod. Ba1: basipod 1 (= coxa). Ba2: basipod 2 (= basis). E: endites. Gnsom: genital somite. |
 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.88, Table I (cont'd)]. Characters as defined for use in cluster analysis. |
 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.89, Table I (cont'd)]. Characters as defined for use in cluster analysis. |
 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.90, Table I (cont'd)]. Characters as defined for use in cluster analysis. |
 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.97, Table III]. Distribution of aesthetascs and peg-sensilla on the segments of the antennule (A1) in the various genera. |
 Issued from : J.C. von Vaupel Klein in Crustaceana, (Supplement) 9, 1984. [p.96, Figs. 20, 21]. Dendogram of Euchirella species-groups and remaining genera, obtained from the initial cluster analysis based on the matrix of the species studied. | | | | | Aetideinae Brodsky, 1950 | | Ref.: | Brodsky, 1950 (1967) (p.142): 12 genres y sont inclus: Aetideopsis, Aetideus, Bryaxis, Chiridius, Derjuginia, Euaetideus, Gaetanus, Gaidiopsis , Gaidius, Pseudaetideus, Pseudogaetanus, Undinopsis.
Vaupel Klein, 1984 a (p.56) ajoute aux genres précédents: Bradyidius, Comantenna, Crassantenna, Lutamator, Paivella, Paracomantenna, Pseudotharybis, Snelliaetideus, Sursamucro, Wilsonidius,? Pseudeuchaeta , ? Valdiviella.
Razouls,1982 (p.153); Zheng Zhong & al.,1984 (1989)(p.235, clé des G.); Mauchline,1988 (p.726, 729: pores cuticulaires) | | | | Euchirellinae Brodsky, 1950 | | Ref.: | Brodsky, 1950 (1967) (p.170) y inclut 5 genres : Batheuchaeta , Chirundina, Euchirella , Pseudochirella , Undeuchaeta. Vaupel Klein, 1984 a (p.55, 56) y ajoute : Chirundinella.
Razouls, 1982 (p.153); Mauchline, 1988 (p.731: pores cuticulaires); Zheng Zhong & al., 1984 (1989) (p.235, clé G.); Vaupel Klein & Rijerkerk, 1996 (p.567)
| | | | (1) Aetideopsis Sars, 1903 ( Aetideinae ) | |
| | Syn.: | Pseudaetideus Wolfenden,1904 (p.115); Brodsky, 1950 (1967) (p.156); Faroella Wolfenden, 1904 (p.117); 1911 (p.213) | | Ref.: | Sars, 1903 (p.159); van Breemen, 1908 a (p.33); A. Scott, 1909 (p.40); Esterly, 1911 (p.314); With, 1915 (p.72, 73); Sewell; 1929 (p.99); Wilson, 1932 a (p.46); Rose, 1933 a (p.90); Davis,1949 (p.25); Brodsky, 1950 (1967) (p.146); Vervoort, 1952 (n°41, p.2); 1952 a (n°42, p.3); Vervoort, 1957 (p.52, Rem.); Tanaka, 1957 a (p.37); Bradford, 1969 (p.73, 92, 95, Rev.); Park, 1975 b (p.272); Bradford, 1976 a (p.9, Rem.); Park, 1978 (p.111, clé spp.); Bradford & Jillett, 1980 (Def., p.17); Shih & Maclellan, 1981 (p.567); Razouls, 1982 (p.162); Mauchline, 1988 (p.726); Ferrari, 1992 (p.392, tab.3); Razouls,1993 (p.309); Markhaseva,1993 (p.54 & foll.); 1996 (p.26, spp Key., Rem.); Chihara & Murano, 1997 (p.681, spp. Key); Mauchline, 1998 (p.88, 92: F; p.92: M); Bradford-Grieve & al., 1999 (p.918, 920: spp. key); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.57: F; 60: M); Vives & Shmeleva, 2007 (p.536, spp. Key) | | Rem.: | type: Aetideopsis rostrata Sars,1903.
12 spp. + 1 indet. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.05 mm (n = 26; SD = 0.687; Cv= 0.8324), and the mean males size 2.979 mm (n = 16; SD = 0.6660). The size ratio Male:Female is 0.851. The sex ratio (F:M) is 1.44. | | | | (2) Aetideus Brady, 1883 ( Aetideinae ) | |
| | Syn.: | Aetidius Brady, 1883 (p.75); Giesbrecht, 1892 (p.53); Wolfenden,1911 (p.207);
Euaetideus Sars, 1925 (p.42); Park,1978 (p.105); Mulyadi, 2004 (p.59);
Snelliaetideus Vervoort,1949 (p.3) | | Ref.: | Giesbrecht & Schmeil, 1898 (p.31); Sars, 1901 a (1903) (p.24); Esterly, 1905 (p.144); van Breemen, 1908 a (p.29); A. Scott, 1909 (p.36); With, 1915 (p.72 & suiv.); Sewell, 1929 (p.99); Wilson, 1932 a (p.44); Rose, 1933 a (p.89); Mori, 1937 (1964) (p.37); Davis, 1949 (p.24); Brodsky, 1950 (1967) (p.144); Vervoort, 1952 (n°41, p.2); 1952 a (n°42, p.3); Tanaka, 1957 a (p.31); Vinogradov, 1968 (1970) (p.109); Bradford, 1971 (p.12, 32, Def., spp. Key); 1972 (p.26, 28); Park, 1974 (p.215, Rem.); Roe, 1975 (p.305); Park, 1978 (p.104); Bradford & Jillett, 1980 (p.11, 14, Def.,); Razouls, 1982 (p.158); Gardner & Szabo, 1982 (p.199, Rem.); Mauchline, 1988 (p.726); Zheng Zhong & al., 1984 (1989) (p.235); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.11, spp. Key, Rem.); Chihara & Murano, 1997 (p.682, spp. Key); Mauchline, 1998 (p.92: F; p.92: M); Bradford-Grieve & al., 1999 (p.879, 918, 920: spp. Key); Boxshall & Halsey, 2004 (p.57: F; 60: M); Vives & Shmeleva, 2007 (p.542, spp. Key) | | Rem.: | type: Pseudocalanus armatus Boeck,1872. 11 spp.
On the basis of the females Bradford (1971) concluded that the only difference between Snelliaetideus Vervoort, 1949 and Aetidius Brady, 1883 was the absence of postero-lateral thoracic points. According to Roe (1975, p.305) the discovery of A. arcuatus male confirms Bradfords opinion and SnelliaetideusAetideus. | | Remarks on dimensions and sex ratio: | | The mean female size is 1,822 mm (n= 22; SD = 0.3671) and the mean male size is 1,562 (n = 19; SD = 0.2996). The size ratio Male:Female is 0.957. The sex-ratio (F/M) is 1,1. | | | | (3) Azygokeras Koeller & Littlepage, 1976 | |
| | Ref.: | Koeller & Littlepage, 1976 (p.1548); Razouls, 1982 (p.230); Gardner & Szabo, 1982 (p.255, Rem.); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.47); Mauchline, 1998 (p.80: F; p.81: M); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M) | | Rem.: | type: Azygokeras columbiae . 1 sp.: | | | | (4) Batheuchaeta Brodsky, 1950 ( Euchirellinae ) | |
| | Ref.: | Brodsky, 1950 (1967) (p.190); Bradford & Jillett, 1980 (Def., p.21); Markhaseva, 1981 (p.1151, 1152, spp.Key F); Razouls, 1982 (p.227); Markhaseva, 1986 a (p.837, spp. Key F,M); Vaupel Klein, 1984 a (p.50, Rem.); Mauchline, 1988 (p.731); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & follo.); 1996 (p.50, spp. Key); Mauchline, 1998 (p.88: F; p.90: M); Boxshall & Halsey, 2004 (p.58: F; 60: M) | | Rem.: | type: Batheuchaeta lamellata Brodsky,1950. 9 spp.: | | Remarks on dimensions and sex ratio: | | The mean female size is 4.616 mm (n = 12; SD = 0.3713), and the mean male size is 4.160 mm (n = 5; SD = 0.2219 ). The size ratio Male:Female is 0.901. The sex ratio (F:M) is 2, value not exceptional for the deep-living species. | | | | (5) Bradyetes Farran, 1905 | |
| | Ref.: | Farran, 1905 (p.31); van Breemen, 1908 a (p.32); With, 1915 (p.73); Rose, 1933 a (p.92); Sewell, 1947 (p.104-106, Rem.); Brodsky, 1950 (1967) (p.142); Vervoort, 1952 (n°41, p.2); 1952 b (n°43, p.3); Grice, 1972 a (p.224, Rem.); Bradford & Jillett, 1980 (Def., p.21); Razouls, 1982 (p.157); Vaupel Klein, 1984 a (p.56); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.62, spp. Key, Rem.); Mauchline, 1998 (p.88: F; p.90: M); Markhaseva & Schnack-Schiel, 2003 (p.116, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M); Ohtsuka & al., 2005 (p.226, Rem.); Vives & Shmeleva, 2007 (p.549); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography) | | Rem.: | Type: Bradyetes inermis Farran,1905. This genus could be synonym of Pseudeuchaeta (in Sewell, 1947), which is undermined by Grice (1972 a)
6 spp. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.471 mm (n = 8; SD = 1.2439), and that of the males 2.040 mm (n = 3; SD = 0.763). The size-ratio Male:Female is 0.588. The sex ratio is 2.5. | | | | (6) Bradyidius Giesbrecht, 1897 ( Aetideinae ) | |
| | Syn.: | Pseudocalanus (part.) : Brady, 1878 (p.46), non Boeck, 1872;
Undinopsis Sars, 1884 (nomen nudum ); 1902 (1903) (p.31); Rose, 1933 a (p.91); Brodsky, 1950 (1967) (p.150) | | Ref.: | Giesbrecht, 1897 b (p.253); Giesbrecht & Schmeil, 1898 (p.32); van Breemen, 1908 a (p.31); A. Scott,1909 (p.39); Mori, 1937 (1964) (p.39); C.B. Wilson, 1950 (p.172, Rem.); Vervoort, 1952 b (p.3); Matthews, 1964 (p.18, Rem.); Bradford, 1969 (p.86, 92, 95, Def.); 1969 b (p.476, 502); 1976 a (p.1, 6, 9); Campaner, 1978 (p.871); Bradford & Jillett, 1980 (Def., p.21); Razouls, 1982 (p.168); Gardner & Szabo, 1982 (p.205, Rem.); Alvarez, 1984 (p.96); Othman & Greenwood, 1987 (p.1137, 1140); Mauchline, 1988 (p.728); Razouls, 1993 (p.309); Ferrari, 1992 (p.392, tab.3); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.68, clé spp., Rem.); Chihara & Murano, 1997 (p.683); Mauchline, 1998 (p.80, 88, 92: F; p.80, 92, 93: M); Markhaseva & Schnack-Schiel, 2003 (p.119, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.57: F; 60: M); Vives & Shmeleva, 2007 (p.549); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography) | | Rem.: | Type: Bradyidius armatus Giesbrecht,1897. 16 spp. (of which 1 doubtful) + 2 unidentified.
Ten species are compared in Bradford, 1976 (table1, p.6).
Diagnosis after Vives & Shmeleva (2007, p.549) :
- Forehead without crest.
- Rostrum , strong, bifurcate, showing points being or no diverging.
- Cephalosome and 1st thoracic segment fused, segments thoracic 4 and 5 fused..
- Posterior corners of the prosome elongated ending in points.
- A1 as long as the prosome.
- Outer spines of the 2nd exopodal segment of P1 larger than the 1st segment. Endopod with 1 segment showing an outer lobe well developed.
- Endopod of P2 2-segmented, exopod 3-segmented.
- Basipodites of P1-P4 with generalemente groups of spinules on surfaces.
Male:
- Crest absent.
- Rostrum as female, but parallel rami..
A1 whether short and not reaching end of the 2nd thoracic segment or exceeding the caudal rami;
- P5 unramous or biramous; in the first instance both rami are equal, in the second instance, endopod has only 1 segment (rarement 2) and generally styliform. | | Remarks on dimensions and sex ratio: | | The mean female size is 2.558 mm (n = 28; SD = 0.7813), and for the males 1.908 mm (n = 21; SD = 0.6377). The size ratio Male: Female is 0.746. The sex ratio (F:M) is 1.25. | | | | | Bryaxis Sars, 1902 ( Aetideinae ) | |
| | Ref.: | Sars, 1902 (1903) (p.35); van Breemen, 1908 a (p.37); With, 1915 (p.73); Rose, 1933 a (p.101); Sewell, 1947 (p.104,105); Brodsky 1950 (1967) (p.143); Vervoort, 1952 b (n°43,p.3); Campaner, 1978 (p.872); Bradford & Jillett, 1980 (p.27); Razouls, 1982 (p.195); 1993 (p.309) | | Rem.: | Genusname established by Boeck,1872, but not published by this author. Cf. Comantenna (part.) & Paracomantenna (part.) | | | | (7) Chiridiella G.O. Sars, 1907 | |
| | Ref.: | Sars, 1907 (p.7); A. Scott, 1909 (p.78); Wolfenden, 1911 (p.292); Sars, 1925 (p.50); Sewell, 1929 (p.101); Rose, 1933 a (p.95); Brodsky, 1950 (1967) (p.142, 192, clé spp.); Vervoort, 1952 (n°41, p.2); 1952 c (p.3); Tanaka, 1957 a (p.56); Bradford, 1971 b (p.20, clé spp.F); Deevey, 1974 (p.439, 468, Rev., clé spp.F); Park, 1978 (p.127); Bradford & Jillett, 1980 (Def., p.22); Razouls, 1982 (p.154); Mauchline, 1988 (p.726); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.81, clé spp., Rem.); Chihara & Murano, 1997 (p.683); Mauchline, 1998 (p.66, 77: F,M; p.88: F; p.90, 93: M); Boxshall & Halsey, 2004 (p.58: F; 60: M); Vives & Shmeleva, 2007 (p.556, spp. Key) | | Rem.: | Type: Chiridiella macrodactyla Sars,1907. 18 spp.:
Diagnosis from Deevey (1974), after Bradford (1971) to, include 16 species..
- Body usually short and thick, with anterior part rather swollen.
- Forehead rounded, no trace of rostrum.
- Head and 1st pedigerous segment separate, 4th and 5th thoracic segments fused.
)- Last metasomal segment rounded.
- Urosome usually very short.
- Genital segment female, strongly protuberant ventrally.
- Caudal rami short.
- A1 of 23 or 24 segments.
- A2 female rather short, exopodite longer than endopodite; male slightly larger than exopodite.
- Md females very large and strongly toothed, palp comparatively short; mandibular blade male reduced and probably useless, palp larger.
- Mx1 with endopodal part of palp little developed in female; reatly reduced in male.
- Mx2 female varyiing in structure from 5 or 6 lobes with spines and setae to 2 or 3 highly modified lobes with hooks and pincers; much reduced in male.
- Mxp with 2nd segment often elongate, terminal part very short.
- P1 with both rami 1-segmented.
- P2-P4 with 2- or 3-segmented exopods, 1- to 3-segmented endopods.
- P5 lacking in female.
- Male P5 as in Aetideidae, 1-segmented endopods larger on left leg. | | Remarks on dimensions and sex ratio: | | The mean female size is 3,096 mm (n= 17; S= 0,655; Cv= 0,212). The mean size for the 2 known males is 2,850 mm. The ratio M/F established in this case is 1,041. The sex ratio is strongly in favor of the females corresponding to their bathypelagic positioning. | | | | (8) Chiridius Giesbrecht, 1892 ( Aetideinae ) | |
| | Syn.: | Euchaeta (part.) Boeck, 1872; Giesbrecht, 1892 (p.301); ? Pseudaetideus Wolfenden, 1904 (p.115: Rem.); Wilson, 1932 a (p.548); Brodsky, 1950 (1967) (p.156); Vervoort, 1952 c (n°44, p.3); Chiridius (part.) : Farran, 1929 (p.230); | | Ref.: | Giesbrecht, 1892 (p.54, 224); Giesbrecht & Schmeil, 1898 (p.33); van Breemen, 1908 a (p.34); Sars, 1900 (p.63); 1901 a (1903) (p.27); A. Scott, 1909 (p.41); Wolfenden, 1911 (p.210); With, 1915 (p.63); Sewell, 1929 (p.100); Wilson, 1932 a (p.47); Rose, 1933 a (p.93); Brodsky, 1950 (1967) (p.152, clé spp.); Vervoort, 1952 c (n°44, p.3); Tanaka, 1957 a (p.47); Matthews, 1964 (p.6, Rem.); Tanaka & Omori, 1970 (p.117, Rem.); Park, 1975 (p.279, clé spp.); 1978 (p.122, Rem.); Bradford & Jillett, 1980 (Def., p.23); Razouls, 1982 (p.172); Gardner & Szabo, 1982 (p.211, Rem.); Mauchline, 1988 (p.729); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.108, clé spp., Rem.); Chihara & Murano, 1997 (p.683); Mauchline, 1998 (p.89, 92: F; p.90: M); Bradford-Grieve & al., 1999 (p.919, 920: clé spp.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M); Vives & Shmeleva, 2007 (p.552, spp. Key) | | Rem.: | Type: Chiridius poppei Giesbrecht,1892. Not all authors admit the synonymy between Chiridius et Pseudaetideus. 8 spp. : | | Remarks on dimensions and sex ratio: | | The mean female size is 2,820 mm (n = 14; SD = 0.8742) and for the males 2,479 mm (n = 13; SD = 0.7435). The size ratio Male:Female is 0.879. The sex ratio (F:M) is 1. | | | | (9) Chirundina Giesbrecht, 1895 ( Euchirellinae ) | |
| | Syn.: | Chirudina : With, 1915 (p.74); Wilsonidius Tanaka, 1969 (p.266); Bradford & Jillett, 1980 (p.92) | | Ref.: | Giesbrecht, 1895 c (p.249); Giesbrecht & Schmeil, 1898 (p.34); Esterly, 1906 a (p.58); van Breemen, 1908 a (p.45); A. Scott, 1909 (p.43); Wolfenden, 1911 (p.241); Sars, 1925 (p.76); Sewell, 1929 (p.119); Wilson, 1932 a (p.48); Rose, 1933 a (p.106); Brodsky, 1950 (1967) (p.180); Vervoort, 1952 (n°41, p.2); 1952 h (n°49, p.3); Tanaka, 1957 b (p.190); Park, 1978 (p.176); Bradford & Jillett, 1980 (Def., p.27); Razouls, 1982 (p.214); Gardner & Szabo,1982 (p.243); Vaupel Klein, 1984 a (p.51, Rem.); Mauchline, 1988 (p.731, cuticular pores); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.129, spp. Key, Notes. p.129); Chihara & Murano, 1997 (p.684); Mauchline, 1998 (p.92: F; p.93: M); Boxshall & Halsey, 2004 (p.57: F; 61: M); Mulyadi, 2004 (p.54); Vives & Shmeleva, 2007 (p.558) | | Rem.: | Type: Chirundina streetsii Giesbrecht,1895. 3 spp. + 1 doubtful: | | Remarks on dimensions and sex ratio: | | The mean female size is 4.470 mm (n = 5; SD = 1.0183) and that of the males 4.113 mm (n = 4; SD = 0.7793). The size ratio Male:Female is 0.920. The sex ratio F:M = 1.5. | | | | (10) Chirundinella Tanaka, 1957 ( Euchirellinae ) | |
| | Ref.: | Tanaka, 1953 (p.131: nom. nudum ); 1957 b (p.197, Def.); Bradford & Jillett, 1980 (Def., p.27, 66); Razouls, 1982 (p.228); Vaupel Klein, 1984 a (p.51, Rem.); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.135, Rem.); Chihara & Murano, 1997 (p.684); Mauchline, 1998 (p.88: F; p.90: M); Boxshall & Halsey, 2004 (p.57: F; 60: M) | | Rem.: | Diagnose after Tanaka (1957, p.197): Female
- Cephalothorax elongate, ovate.
- Head and 1st pediger segment separate, 4th and 5th pediger segments fused.
- Head with a median crest.
- Rostrum one-pointed.
- Abdomen 4-segmented in female.
- Outer marginal seta of caudal rami short; appendicular seta very long.
- A1 24-segmented, segments 8-9 fused, 24-25 separte.
- A2 exopod longer than endopod.
- Md with a small endopod, slightly longer than half the length of exopod; biting part robust.
- Mx1 with 4 setae on 2nd basal segment; 10 setae on endopod.
- Mx2 and Mxp resemble those of Pseudochirella
- P1 exopod 3-segmented, endopod 2-segmented.
P3 and P4 each exopod and endopod 3-segmented.
- P4 with a row of bristles on inner margin of coxa.
P5 female absent.
- Male unknown.
Male describes by Vervoort 1949 as Pseudochirella magna. | | | | (11) Comantenna Wilson, 1924 ( Aetideinae ) | |
| | Syn.: | Bryaxis Sars, 1902 (1903) (p.35); Bryaxona Strand, 1929 | | Ref.: | Wilson, 1924 (p.14, Rem.); Bradford, 1969 b (p.484, 502); Campaner, 1978 (Def., p.872); Bradford & Jillett, 1980 (Def., p.27); Razouls, 1982 (p.196); 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); Alvarez, 1986 (p.858, 876: Rem.); Markhaseva, 1996 (p.135, clé spp., Rem.); Mauchline, 1998 (p.88: F; p.90: M); Markhaseva & Schnack-Schiel, 2003 (p.116, 118, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography) | | Rem.: | type: Bryaxis brevicornis . 5 spp. : | | Remarks on dimensions and sex ratio: | | The mean female size is 2.841 mm (n = 7; SD = 0.9157); but there seem to be two size groups species (numbered 1, 3, 4) 2.175 mm and (species numbered 2, 5) 3.775 mm, corresponding no doubt to the sampling depths. The sex ratio is strongly biased in favor of the females. | | | | (12) Crassantenna Cole, Green, Croft & Rawlins, 1972 ( Aetideinae ) | |
| | Ref.: | Bradford, 1969 b (p.493, 502); Cole & al., 1972 (in Zool. Rec., 106, section 10, p.194); Bradford & Jillett, 1980 (Def., p.29); Razouls, 1982 (p.229); 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.143, clé spp., Rem.); Mauchline, 1998 (p.89: F); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, biogeography) | | Rem.: | Type: Crassantenna comosa. The swimming feet are not intact in the two species being the origin of this genus. Total: 2 spp.
Diagnosis from Bradford (1969, p.493) :
Female:
- head and 1st thoracic segment fused (with fusion line visible dorsally), 4yj and 5th thoracic segments fused.
- Posterior borders of prosome up-yurned.
- Genital segment slightly swollen anteriorly;
- Small single rostrum.
- A1 24-segmented, reaching at least thoracic segment 3, and covered with annulate setae.
- A2 with thick squat endopod, 2 setae on segment 2 of exopod; segment 1 of exopod with or without 2 setae.
- Md palp large with small endopod.
- Mx1, inner lobe 1 with 12 spines and setae.
- Mx2 with lobe 5 enlarged.
- Mxp basipod 1 short and thick with no sensory appendage distally.
- P1 endopod 1-segmented.
- P2 endopod 2-segmented, all other rami 3-segmented. External spines on exopods large. Terminal spines on exopods bear numerous sharp teeth. Exopod segments where external spine inserted not produced into spines externally.
- P5 absent.
- Male unknown. | | Remarques sur les dimensions et le sex-ratio: | | The mean female size is 3.667 mm (n = 3; SD = 0.3055. No male known. | | | | | Derjuginia Jaschnov, 1947 ( Aetideinae ) | |
| | Ref.: | Brodsky, 1950 (1967) (p.157); Bradford & Jillett, 1980 (Def.,p.29); Markhaseva, 1980 a (p.63); Razouls, 1982 (p.177); 1993 (p.309) | | Rem.: | Cf. Jaschnovia | | | | Euaetideus Sars, 1925 ( Aetideinae ) | | Ref.: | Sars, 1925 (p.42); Rose, 1933 a (p.90); Brodsky, 1950 (1967) (p.143); Vervoort, 1952 a (n°42, p.3); Bradford, 1971 (p.12 & suiv., Rem.); Park, 1978 (p.105, Rem.); Bradford & Jillett, 1980 (p.11, 14, Rem.); Razouls, 1982 (p.197); 1993 (p.309) | | Rem.: | type: Aetideus giesbrechti Clève,1904. Cf. Aetideus | | | | (13) Euchirella Giesbrecht, 1888 ( Euchirellinae ) | |
| | Ref.: | Giesbrecht, 1892 (p.54, 232); Giesbrecht & Schmeil, 1898 (p.34, spp. Key); Esterly, 1905 (p.150, spp. Key); Wolfenden, 1905 a (p.17); van Breemen, 1908 a (p.46); A. Scott, 1909 (p.53); Wolfenden, 1911 (p.234); With, 1915 (p.72 & suiv.); Sars, 1925 (p.65); Sewell, 1929 (p.106, Rem.); Wilson, 1932 a (p.55, clé spp.); Rose, 1933 a (p.102, spp. Key); Mori, 1937 (1964) (p.41); Sewell, 1947 (p.69, 70: 2 groups); Davis, 1949 (p.29); Brodsky, 1950 (1967) (p.171, spp. Key); Vervoort, 1952 (n°41, p.2); 1952 f (n°41, p.3); 1957 (p.63, 64, Rem.); Tanaka, 1957 b (p.176); Vervoort, 1963 b (p.134, Rem.); Owre & Foyo, 1967 (p.45, spp. Key); Tanaka & Omori, 1969 (p.33); 1969 a (p.155); Park, 1976 a (p.101, Rev., clé spp.); Vaupel Klein, 1972 (p.497, Rev.); Park, 1978 (p.147); Vaupel Klein, 1980 (p.151 & suiv.); Bradford & Jillett, 1980 (Def., p.30); Razouls, 1982 (p.201); Gardner & Szabo, 1982 (p.233); Vaupel Klein, 1984 a (.p.33, 164 female: Def., Rev., Key to the Species-Groups females: p.166); Mauchline, 1988 (p.730); Zheng Zhong & al., 1984 (1989) (p.235); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.145, spp. Key, Rem.); Chihara & Murano, 1997 (p.684, spp. Key); Mauchline, 1998 (p.92: F; p.93: M); Vaupel Klein, 1998 (p.153, Rem.: phylogeny, Key); 1998 a (p.383 & suiv., Rem. speciation); Bradford-Grieve & al., 1999 (p.919, 921: clé spp.); Boxshall & Halsey, 2004 (p.57, 58 F; 60, 61: M); Mulyadi, 2004 (p.40); Vives & Shmeleva, 2007 (p.560, spp. Key) | | Rem.: | Type: Undina messinensis Claus,1863. 24 spp.
The bathymetric distribution of these forms shows generally a very wide amplitude.
For Tanaka (1957, p/176) the genus is characterized by the head usually separates from 1st thoracic segment; head with or without a crest; last thoracic segments fused; lateral corners of last thoracic segment rounded; rostrum usually present; abdomen 4-segmented in female; genital segment often asymmetrical: caudal rami about as long as wide; A1 23-segmented; endopod of A2 shorter than half the length of exopod; 2nd basal segment of Mx1 long; exopod of P1 2-segmented; exopod of P2 to P4 3-segmented; endopod of P1 and P2 1-segmented; endopod of P3 and P4 3-segmented; coxa of female P4 with spines on the inner margin; P5 absent in female; male P5 biramous on the right; left leg with a rudimentary endopod.
For Vervoort (1957, p.63) there is a great deal of confusion concerning the limitations of the genera Euchirella and Pseudochirella, representatives of which resemble each other in many respects. The menbers of the genus Euchirella (known in 1957), all have a reduced number of setae on the 2nd basal segment and on the endopod of Mx1 (2-3 and 4-6 respectively) ; in Pseudochirella this number is increased (5 and 14-16 respectively). The endopod of A2 is reduced in nearly all Euchirellas (it is less than half the length of the exopod in many species and about half that length in some others ; it is, however, much less reduced in Pseudochirella. Both Euchirella and Pseudochirella are characterized by the presence of a spine or spinules on the posterior surface of the 1st basal segment of P4. Euchirella brevis Sars (1905), approaches genera Gaetanus by the presence of thin, tube like spinules ; all other species have stronger spinules or a single strong spine. Ann additional difficulty in both genera is the scarcity of adult males and the obstacles encountered when trying to identity both sexes of a species. | | Remarks on dimensions and sex ratio: | | The mean female size is 4,780 mm (n = 48; SD = 1.3573), and that of the males 4,282 mm (n = 33; SD = 1.1607). The size ratio Male:Female is 0,896. The sex ratio is 1,33. | | | | | (14) Gaetanus Giesbrecht, 1888 ( Aetideinae ) | |
| | Syn.: | Gaëtanus Giesbrecht, 1888 (p.335);
Chiridius (part) Sars;
Mesogaidius (part.) Wolfenden, 1911 (p.223);
Pseudogaetanus Brodsky, 1950 (1967) (p.143, 144, 169);
Gaidius Giesbrecht, 1895 c (p.249); Giesbrecht & Schmeil, 1898 (p.32); Sars, 1903 (p.161); Esterly, 1905 (p.145); van Breemen, 1908 a (p.35); A. Scott, 1909 (p.51); Wolfenden, 1911 (p.222); With, 1915 (p.72 & suiv.); Sars, 1925 (p.46, 57); Sewell, 1929 (p.100); Wilson, 1932 a (p.52); Rose, 1933 a (p.96, clé spp.); Davis, 1949 (p.26); Brodsky, 1950 (1967) (p.158, spp. Key); Vervoort, 1952 (n°41, p.2); 1952 d (n°45, p.3); Tanaka, 1957 a (p.60); Park, 1975 a (Rev., p.9,10); Park, 1978 (p.127); Bradford & Jillett, 1980 (Def., p.57); Razouls, 1982 (p.178); Gardner & Szabo, 1982 (p.217); Mauchline, 1988 (p.729); Razouls, 1993 (p.309); Chihara & Murano, 1997 (p.687, spp. Key); Mauchline, 1998 (p.88, 92: F; p.90, 93: M) | | Ref.: | Giesbrecht, 1892 (p.53, 219); Giesbrecht & Schmeil, 1898 (p.32); Esterly, 1906 a (p.56); Van Breemen, 1908 a (p.38, clé spp.); A. Scott, 1909 (p.44); Wolfenden, 1911 (p.226); With, 1915 (p.72 & suiv.); Sars, 1925 (p.53, Rem: p. 57); Sewell, 1929 (p.101); Wilson, 1932 a (p.50); Rose, 1933 a (p.98); Mori, 1937 (1964) (p.40); Sewell, 1947 (p.56, Rev.); Davis, 1949 (p.28); Brodsky, 1950 (1967) (p.163, clé spp.); Vervoort, 1952 (n°41, p.2); 1952 e (n°46, p.2); Tanaka, 1957 b (p.169); Park, 1975 a (Rev., p.9, clé spp.); 1978 (p.136, clé spp.); Bradford & Jillett, 1980 (Def., p.45); Razouls, 1982 (p.185); Gardner & Szabo, 1982 (p.225); Vaupel Klein, 1984 a (p.55, Rem.); Mauchline, 1988 (p.729); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.176, spp. Key, Rem.); Chihara & Murano, 1997 (p.686, Part., spp. Key); Mauchline, 1998 (p.87, 92: F; p.90: M); Bradford-Grieve & al., 1999 (p.919, 922: clé spp.); Boxshall & Halsey, 2004 (p.57: F; 60, 61: M); Mulyadi, 2004 (p.57); Vives & Shmeleva, 2007 (p.573, spp. Key) | | Rem.: | Type: Gaëtanus miles Giesbrecht, 1888 (p.335). Park, 1975 a (p.11-12) joins together the two genera Gaetanus and Gaidius, but does not maintain this fusion in 1978 (p.127-136): Bradford & Jillett (1980) give a definition of the two genera. Markhaseva, 1996 retrieves the synonymy of the two genera, including 2 new species and correcting some synonymies. Provisional total: 25 spp. + 4 unidentified. | | Remarks on dimensions and sex ratio: | | The mean female size is 4.473 mm (n = 49; SD = 1.6722), and for the males 3.530 mm (n = 29; SD = 1.2256). The size ratio Male:Female is 0.789. The sex ratio F:M is 1.5. | | | | | (15) Gaidiopsis A. Scott, 1909 ( Aetideinae ) | |
| | Ref.: | A. Scott, 1909 (p.52); Sewell, 1929 (p.101); Tanaka, 1957 b (p.169); Brodsky, 1950 (1967) (p.143); Bradford & Jillett, 1980 (p.55, 57, Rem.); Razouls, 1982 (p.197); 1993 (p.309); Markhaseva, 1996 (p.8, Rem.); Mauchline, 1998 (p.89: F) | | Rem.: | Type: Gaidiopsis crassirostris A. Scott, 1909. Total: 1 sp.
Diagnosis after A. Scott (1909, p.52):
Forehead without cephalic spine, wxith short one-pointed rostrum.
4th and 5th thoracic segments completely separated.
5th segment peoduced into strong lateral spines.
Abdomen 4-segmented.
A1 24-segmented.
A2, Md, Mx1 and Mxp, almost similar to those of Gaidius (= Gaetanus).
Exopodite of P1 distinctly 3-segmented.
P2, P3 and P4 similar to those of Gaidius (= Gaetanus).
Endopodite of P2 distinctly 2-segmented.
Remarks: This new genus resembles Aetideopsis and also Gaidius (= Gaetanus). | | | | Gaidius Giesbrecht, 1895 ( Aetideinae ) | |
| | Syn.: | Chiridius (part) Sars; Mesogaidius (part.) Wolfenden, 1911 (p.223); Pseudogaetanus Brodsky, 1950 (1967) (p.143, 144, 169) | | Ref.: | Gaidius Giesbrecht, 1895 c (p.249); Giesbrecht & Schmeil, 1898 (p.32); Sars, 1903 (p.161); Esterly, 1905 (p.145); van Breemen, 1908 a (p.35); A. Scott, 1909 (p.51); Wolfenden, 1911 (p.222); With, 1915 (p.72 & suiv.); Sars, 1925 (p.46, 57); Sewell, 1929 (p.100); Wilson, 1932 a (p.52); Rose, 1933 a (p.96, clé spp.); Davis, 1949 (p.26); Brodsky, 1950 (1967) (p.158, clé spp.); Vervoort, 1952 (n°41, p.2); 1952 d (n°45, p.3); Tanaka, 1957 a (p.60); Park, 1975 a (Rev., p.9,10); Park, 1978 (p.127); Bradford & Jillett, 1980 (Def., p.57); Razouls, 1982 (p.178); Gardner & Szabo, 1982 (p.217); Mauchline, 1988 (p.729); Razouls, 1993 (p.309); Bradford-Grieve, 2004 (p.287) | | Rem.: | type: Gaidius pungens Giesbrecht,1895. Markhaseva (1996, p.176) considers this genus as a junior synonym of Gaetanus. | | | | (16) Jaschnovia Markhaseva, 1980 | |
| | Syn.: | Derjuginia Jaschnov, 1947; Brodsky, 1950 (1967) (p.157); Bradford & Jillett, 1980 (Def.,p.29) | | Ref.: | Markhaseva, 1980 a (p.63); Razouls, 1982 (p.177); 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.225, Rev.); Mauchline, 1998 (p.78); Boxshall & Halsey, 2004 (p.58: F; 60: M) | | Rem.: | 2 spp. | | Remarks on dimensions and sex ratio: | | The mean female size is 2.275 mm. (n = 4, SD = 0.301), and the mean male size is 2.120 mm. (n = 2, SD = 0.). The size ratio male: female is 0.932. | | | | (17) Lutamator Bradford, 1969 ( Aetideinae ) | |
| | Ref.: | Bradford, 1969 b (p.491); Bradford & Jillett, 1980 (Def.,p.61); Razouls, 1982 (p.229); Alvarez, 1984 (Rev., p.99); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.226, spp. Key, Rem.); Mauchline, 1998 (p.89: F); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.56, 58); Ohtsuka & al., 2005 (p.230, Rem.) | | Rem.: | Type: Lutamator hurleyi Bradford, 1969. 3 spp.).
Diagnosis from Bradford (1969, p.491) [provisionally until further species are discovered] :
- head and 1st thoracic segment 1fused, 4th and 5th thoracic segments fused.
- Posterior thoracic borders extended into short points .
- Rostrum bluntly rounded.
- Posterior borders of abdominal segments edged with a weakly chitinised striated fringe.
- A1 24-segmented with long antennulate setae.
- A2 endopod and exopod equal; segment 1 of endopod wider proximally than distally; exopod tapers distally, segments 1 and 2 with 1 and 2 setae.
-- Md palp large.
- Mx1 inner lobe 1 (arthrite) with 11 spines and setae.
- Mxp basipod 1 twice as long as wide.
- P1 endopod 1-segmented.
- P2 endopod 2-segmented, all other rami 3-segmented; external spines on exopods large. Teeth on terminal spine of exopods connected by a lamella.
After Markhaseva & Schulz (2008, p.33) female specimens of Prolutamator share with Lutamator Bradford (1969) the following features: 1- rostrum as a poorly developed blunt plate; 2- A2 exopod nearly as long as endopod; 3- Md endopodal segment 2 with fewer than 7 setae; 4- coxal endite of Mxp syncoxa without sensory element; 5- P5 absent.
Except for the above characters (1-4) the genus Lutamator was defined by: 1- A2 endopodal segment 1 wider proximally than distally; 2- A2 exopodal segment 1 with 1 seta; 3- number of setae on the praecoxal arthrite of Mx1 reduced (11 setae). Its definition was considered tentative ''until further species are discovered'' (Bradford, 1969).
The genus was redefined by Alvarez (1984) and two species were included (L. hurleyi, L. elegans). The redefinition in genertal follows the earlier definition, but unfortunately has a misleading sentence that lacks the correct name of the described limb. | | Remarks on dimensions and sex ratio: | | The mean female size is 4.470 mm. (n = 5, SD = 0.914), and the mean male size is 3.380 mm (n = 2, SD = 0.014). The size ratio male: female is 0.974. | | | | | (18) Mesocomantenna Alvarez, 1986 | |
| | Ref.: | Alvarez, 1986 (p.865, 876: Rem.); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.231, Rem.); Mauchline, 1998 (p.88: F); Markhaseva & Schnack-Schiel, 2003 (p.118, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58) | | Rem.: | 1 sp.: | | | | Mesogaidius Wolfenden, 1911 | |
| | Ref.: | Wolfenden, 1911 (p.223); With, 1915 (p.73, 74, Rem.); Park, 1975 a (p.12: syn. de Gaetanus); Bradford & Jillett, 1980 (p.57: syn. de Gaidius); Razouls, 1982 (p.184); 1993 (p.309); Markhaseva, 1996 (p.176: syn. de Gaetanus) | | Rem.: | Cf. Gaetanus | | | | Mesundeuchaeta Wolfenden, 1911 | |
| | Ref.: | Wolfenden, 1911 (p.244); Bradford & Jillett, 1980 (p.80); Markhaseva, 1996 (p.301) | | Rem.: | Cf. Undeuchaeta | | | | (19) Paivella Vervoort, 1965 ( Aetideinae ) | |
| | Ref.: | Vervoort, 1965 (p.199, Rem.); Wheeler, 1970 (p.10, Rem.); Bradford, 1971 (p.31); Bradford & Jillett, 1980 (Def., p.63); Razouls,1982 (p.228); 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.233, spp. key, Rem.); Mauchline, 1998 (p.92: F; p.93: M); Bradford-Grieve & al., 1999 (p.918, 922: clé spp.F); Boxshall & Halsey, 2004 (p.57: F; 60: M) | | Rem.: | type: Paivella inaciae Vervoort,1965. 2 spp.: | | Remarks on dimensions and sex ratio: | | The mean female size is 1.360 mm (n = 4; SD = 0.0860). The size ratio M:F = 0.804 (following only 1 species) | | | | (20) Parabradyidius Schulz & Markhaseva, 2000 | |
| | Ref.: | Schulz & Markhaseva, 2000 (p.78); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, biogeography) | | Rem.: | type: Parabradyidius angelikae Schulz & Markhaseva, 2000. Total: 2 spp. (which one juvenil).
Diagnosis from Schulz & Markhaseva (2000, p.78) :
Female:
- Rostrum strongly bifurcate lacking filaments.
- 5th pedigerous somite tapering into long pointed process posteriorly.
- A2 exopod and endopod subequal in length; 1st exopodal segment with 1 seta, 2nd with 3 setae, 7th segment lacking medial seta.
- Md basis bearing 2 setae.
- Mx1 exopod with 11 setae.
- Exopod of P1 3-segmented; outer spine of 1st segment present and more slender than that of 2nd segment.
- Coxa of P1, P2 and P3 with outer patch of spinules.
- Posterior surfaces of P2 to P4 partly covered by spinules.
- P5 present, uniramous, indistinctly 3-segmented; apical segment small bearing spines or spine-like projections.
Male:
- Rostrum similar to that of female bur less diverging distally.
- Posterior corner of 5th pedigerous somite with small pointed process.
- Cephalosomal limbs more or less reduced.
- P5 biramous on both sides, simple, left leg longer than right.
The new genus differs from Aetideopsis, Bradyidius, Pseudotharybis in relative length of A2 endo- and exopod: while in these three genera the exopod is distinctly longer than the endopod (ca. 1.2 - 1.5 times), in Parabradyidius these rami are subequal or the exopod may be even slightly shoryter. In addition, Parabradyidius lacks the medial seta of the A2 7th exopodal segment present in these three taxa. | | Remarks on dimensions and sex ratio: | | For one specie only: The mean female size is 5.435 mm, and the mean male size 4.975 mm. The size ratio (male : female) is 0.915. | | | | (21) Paracomantenna Campaner, 1978 ( Aetideinae ) | |
| | Syn.: | Bryaxis (part.) | | Ref.: | Campaner, 1978 (p.871); Bradford & Jillett, 1980 (Def., p.63); Alvarez, 1986 (p.858, 876: Rem.); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.236, spp. Key, Rem.); Mauchline, 1998 (p.88: F); Bradford-Grieve & al., 1999 (p.919, 922: spp. Key); Markhaseva & Schnack-Schiel, 2003 (p.118, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58); Ohtsuka & al., 2005 (p.240, Rem.); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography); Markhaseva & Renz, 2019 (p.223: Rem) | | Rem.: | type: Bryaxis minor Farran,1905. Total: 6 spp.: | | Remarks on dimensions and sex ratio: | | The mean female size is 1.994 mm (n = 9; SD = 0.4683). Only 1 species with the male has been observed (mean = 1.225 mm; n = 2; SD = 0.353) | | | | (22) Prolutamator Markhaseva & Schulz, 2008 | |
| | Ref.: | Markhaseva & Schulz, 2008 (p.32); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, biogeography) | | Rem.: | Type: Prolutamator hadalis Markhaseva & Schulz, 2008. Total: 2 spp.. Total: 3 spp.
Abyssal above the sea bed.
Diagnosis from Markhaseva & Schulz (2008, p.32) :
Female:
- Cephalosome and pediger 1 more or less separate, 4th and 5th separate.
- Posterior corners of prosome short and rounded.
- Rostrum a poorly blunt plate.
- Urosome 4-segmented.
- Genital double-somite symmetrical.
- A1 24-segmented, 1st segment with 1 seta.
- A2 endopod nearly as long as exopod; endopodal segment 1 moderately wide in proximal part bearing 2 setae; basdis and exopodal segment 1 with 2 setae each.
- Md basis with 2-3 setae; endopoda segment 2 with 5-6 setae.
- Mx1 praecoxal endite with 9 terminal, 4 posterior and 1 anterior setae; coxal epipodite with 5-6 setae; distal basal endite with 5 setae; endopod with 14-15 setae.
- Mx2 endopod with 6 long and 2 short setae.
- Mxp proximal praecoxal endite with 1 seta, middle and distal endites with 2 and 3 setae, respecyively; coxal endite without sensory element.
- P1 exopodal segments 1-3 with lateral spine each.
- Segmentation of P1-P4 as typical of Aetideidae.
- P5 absent.
Male unknown. | | Remarks on dimensions and sex ratio: | | The mean female size is 2.550 mm (n = 4; SD = 0.3028). No male observed. | | | | | Pseudaetideus Wolfenden, 1904 ( Aetideinae ) | | Ref.: | Wolfenden, 1904 (p.115); van Breemen, 1908 a (p.32); Brodsky, 1950 (1967) (p.156); Vervoort, 1952 c (n_44, p.3); Matthews, 1964 (p.6); Park, 1975 b (p.272, 273, Rem.); Bradford & Jillett, 1980 (p.19, Rem.); Razouls, 1982 (p.176); 1993 (p.309); Markhaseva, 1996 (p.26) | | Rem.: | type: Euchaeta armata (Boeck,1872). For Vervoort (1952) this species is close to Chiridius obtusifrons . Cf. Aetideopsis | | | | (23) Pseudeuchaeta Sars, 1905 ( Aetideinae ) | |
| | Syn.: | ? Bradyetes Farran, 1905 (p.31);
Autanepsius Wolfenden, 1906 (p.39); 1911 (p.350); | | Ref.: | Sars, 1905 b (p.18); With, 1915 (p.73); Sars, 1925 (p.102); Rose, 1933 a (p.124); Sewell, 1947 (p.104, 105, Rem.); Vervoort, 1952 (n°41, p.2); 1952 h (n°49, p.3); 1957 (p.70); Bradford, 1969 b (Def., p.486); Park, 1978 (p.187); Bradford & Jillett, 1980 (Def. , p.21,63); Razouls, 1982 (p.230); Vaupel Klein, 1984 a (p.57); Markhaseva, 1986 b (p.1896, clé spp.); Mauchline, 1988 (p.729); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.240, clé spp., Rem.); Chihara & Murano, 1997 (p.688); Mauchline, 1998 (p.78); Markhaseva & Schnack-Schiel, 2003 (p.118, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M); Vives & Shmeleva, 2007 (p.593); Renz & Markhaseva, 2015 (p.96, Table 4, Fig.3, sex ratio, biogeography) | | Rem.: | type: Autanepsius minor Wolfenden, 1906. 8 spp.
Definition after Bradford & Jillett (1980, p.63) :
- Robust copepods with blunt, rounded rostrum.
- Head and pediger segment 1 partially fused, 4th and 5th pediger segments partially fused.
- Posteolateral corners of last thoracic segment extended into points
- Posterior borders of urosomal segments edged with weakly scleritized fringe.
- Female A1 24-segmented, extending at most length of prosome, bears few sensory filaments and numerous annulate setae, long distally , endopod setae with as in Bradyidius
- A2 exopod and endopod equal with 2 setae on exopodal segments 1 and 2.
- Endopod of Md small.
- Mx1 inner lobe 1 (arthrite) with 11 or 13 spines and setae, posterior surface having 1 or 3 setae.
- Mxp large with sensory organ on basipodal segment 1 (as in Comantenna), endopod setae with crescent-shaped appendages along their length.
- Exopodal segment 1 of P1 without outer edge spine; external exopod spines large.
- Posterior surface of endopods of P3 and P4 with fine spinules.
- Terminal spines of leg exopods with long widely-spaced teeth joined by a lamella.
P5 absent.
Male:
- A1 23-segmented.
- Mouthparts slightly reduced.
- Basipodal segment 1 of Mxp with distal sensory appendage.
- Left P5 with exopod 3-segmented, small lobe on segment 3 lined with spinules, endopod 1-segmented, equals length of exopodal segment 1; right leg exopod 3-segmented, segments 1 and 3 with outer edge spine, segment 3 with long terminal spine extending as sword-like expansion , endopod 1-segmented, club-shaped. | | Remarks on dimensions and sex ratio: | | The mean female size is 7.903 mm (n = 9; SD = 1.9168). The size ratio (Male:Female) is 1.03 but established for a single species. and not representative. The sex ratio is in disfavor of the males, like one observes in the forms living at great depths. | | | | | (24) Pseudochirella Sars, 1920 ( Euchirellinae ) | |
| | Ref.: | Sars, 1920 c (p.5); Sars, 1925 (p.83); Sewell, 1929 (p.127); Rose, 1933 a (p.109); Sewell, 1947 (p.96, Rem.); Davis, 1949 (p.32); Brodsky, 1950 (1967) (p.185, Key spp.); Vervoort, 1952 (n°41, p.2); 1952 g (n°48, p.3); 1957 (p.63, 64, Rem.); Tanaka,1957 b (p.192); Tanaka & Omori, 1969 a (p.155); Park, 1978 (p.158); Bradford & Jillett, 1980 (Def., p.66, Rem.); Razouls, 1982 (p.220); Gardner & Szabo, 1982 (p.251); Vaupel Klein, 1984 a (p.52, Rem.); Mauchline, 1988 (p.730); Markhaseva, 1989 (p.37, spp.: Key F & M); Razouls, 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); Vaupel Klein, 1996 (p.446, Rem., Def., p.450); Markhaseva, 1996 (p.253, clé spp., Rem.); Chihara & Murano, 1997 (p.688); Mauchline, 1998 (p.87, 92: F; p.90, 93: M); Bradford-Grieve & al., 1999 (p.919, 923: Key spp.); Boxshall & Halsey, 2004 (p.58, 61); Vives & Shmeleva, 2007 (p.595, spp. Key) | | Rem.: | type not designated, Pseudochirella obesa proposed by von Vaupel Klein (1984 a, p.52).
Total: 30 spp. + 1 unidentified.
Definition after Bradford & Jillett (1980, p.66) :
Female
- Body resembling Euchirella and Chirundina.
- Head and pediger segment 1 fused, segments 4 and 5 separate or fused.
- Forehead rounded in lateral view, without crest.
- Rostrum distinct, one-pointed.
- Posterolateral corners of last thoracic segment rounded or produced into spines.
- Genital segment asymmetrical in many forms.
- A1 as long as body.
- A2 endopod strongly developed, usually one half length of exopod or slightly longer; exopod segments 1 and 2 separate.
- Mx1 with larger number of setae than in Euchirella: 5 setae on basipodal segment 2; 16 setae on endopod.
- Segments of legs as in Euchirella.
- Exopodal segment 1 of P1 with well developed outer edge spine.
- Coxa of P4 with 6-14 strong spines on posterior surface.
- Setae on outer edge of caudal rami thin and usually elongate.
Male:
- Slender body.
- A1 with reduced number of segments, some setae thickened.
- A2 endopod more than half length of exopod.
- P5 with coxa and basis swollen; endopod elongate, 1-segmented, truncate at apex; left leg exopod 3-segmented, segments short, last segment rounded distally; right leg exopod with 2 elongate segments. | | Remarks on dimensions and sex ratio: | | The mean female size is 6.175 mm (n = 50; SD = 1.3015) and that of males 5.564 mm (n = 20; SD = 0.8959). The size ratio Male:Female is 0.901. The sex ratio (F:M) is 4.33. | | | | | Pseudogaetanus Brodsky, 1950 ( Aetideinae ) | |
| | Ref.: | Brodsky, 1950 (1967) (p.169); Grice & Hulsemann, 1967 (p.24, Rem.); Park, 1975 a (p.12); 1978 (p.127); Bradford & Jillett, 1980 (p.57); Razouls, 1982 (p.195); 1993 (p.309); Markhaseva, 1996 (p.176) | | Rem.: | type: Pseudogaetanus robustus (Sars,1905). Cf. Gaetanus | | | | (25) Pseudotharybis T. Scott, 1909 ( Aetideinae ) | |
| | Ref.: | T. Scott, 1909 (p.126); Wilson, 1932 a (p.551, 553); Bradford, 1976 a (p.9, Rem.); Bradford & Jillett, 1980 (Def., p.71); Razouls, 1982 (p.198); Bradford & al., 1983 (p.123); Razouls, 1993 (p.309); Mauchline, 1998 (p.84: F); Markhaseva, 1996 (p.8, Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58: F; 60: M); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, sex ratio, biogeography) | | Rem.: | Genus excluded from the Tharybidae by Bradford (1976 a) and considered as intermediary between the Aetideidae and the Tharybidae by Markhaseva (1996). Type: Pseudotharybis zetlandicus. Total: 7 spp.
Definition after Bradford & Jillett (1980, p.71) :
- All characteristics as in Bradyidius except that females have P5 3-segmented, terminal segment with 2 terminal and 1 external spine-like extensions.
- Males also almost identical with Bradyidius in having both biramous and uniramous P5, but instead of posteriorly directed spine arising from very posterior last thoracic border; Pseudotharybis males have a small tooth placed more dorsally on last metasomal segment. | | Remarks on dimensions and sex ratio: | | The mean female size is 4.213 mm (n = 6; SD = 0.9517), and 3.972 mm for the males (n = 5; SD = 0.4358). The size ratio Male:Female 0.943. | | | | | (26) Pterochirella Schulz, 1990 | |
| | Ref.: | Schulz, 1990 (p.182); Razouls, 1993 (p.309); Markhaseva, 1996 (p.295, Rem.); Mauchline, 1998 (p.66); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.60: M) | | Rem.: | Type: Pterochirella tuerkayi Schulz, 1990. Total: 1 sp.
This genus shows affinities with Bradyetes and Pseudeuchaeta.
Diagnosis from Schulz (1990, p.182) :
Male :
- Cephalosome and 1st pedigerous somite fused, 4th and 5th pedigerous somite fused.
- Dorsal body wall of prosome transformed into deep transverse cleft encompassed by number of paired lobe-like structures.
- 1st pedigerous somite with paired winglike extension.
- Urosome 5-segmented.
- Rostrum absent, 2 delicate filaments present.
- A2 with endopod equaling exopod in length.
- Mouthparts considerably reduced.
- Endopods of P1 1-segmented, of P2 2-segmented, all other rami 3-segmented.
- P5 biramous with left and right protopod of about equal length, not swollen; endopods 1-segmented; exopods 2- to 3-segmented; distal segment of right side particularly long, tapering distally.
For Schulz (1990, p.187) according to Bradford & Jillett (1980) several features of this genus point to the family Aetideidae. In the male sex: 1 - fusion of cephalosome with 1st pedigerous somite and of somites 4 and 5; 2 - reduction or total lack of a rostrum; 3 - reduced number of segments of A1, swollen proximal segments armed with many large club-shaped aesthetascs; 4 - considerably reduced mouthparts; 5 - segmentation of P1 to P4, and pronounced, acute teeth on terminal spines of P2 to P4; 6 - presence of both endopods on P5. Furthermore, the presence of rows of spinules on the prominent outer lobe of the endopod of P1, described by Vaupel-Klein (1972) for the genus Euchirella, seems to support this classification. Pterochirella differs from both genera in the incomplete fusion of segments 9 and 10, 12 and 13, and complete fusion of segments 24 and 25 of the A1. Segments 20 and 21 are fused on the right but separate on the left A1. Furthermore, Pterochirella lacks the extraordinarily strong spine on lobe 5 of the Maxilla typical of Bradyetes. P5 of Pterochirella is distinguished from those of Bradyetes and Pseudeuchaeta by the comparatively short left protopod and the relative length of the endopods of which the right one is longer than the left. Morever, the right exopod terminates in a more or less straight distal segment, not curved as it is found in the other genera..
Pterochirella is assignable to the subfamily Euchirellinae of Brodsky (1950 [1967]) characterized particularly by 1- the more complex structure of male P5 and 2- the shortened urosome in the female (absent still in 2015). | | | | | (27) Senecella Juday, 1923 | |
| | Ref.: | Juday, 1925 (p.1); Gurney, 1931 a (p.19); Dussart, 1983 (p.9); Vyshvartzeva, 1994 (p.113); Markhaseva, 1996 (p.297, clé spp., Rem.); Boxshall & Halsey, 2004 (p.58: F; 60: M) | | Rem.: | freshwater (relict) and brackish. Total: 2 spp.: | | Remarks on dimensions and sex ratio: | | The mean female size is 2.957 mm (n = 4; SD =: 0.2706), and for the males 2,670 mm n = 4; SD = 0.203). The size ratio (Male:Female) is 0.903. The sex ratio is 1. | | | | Snelliaetideus Vervoort, 1949 ( Aetideinae ) | |
| | Ref.: | Vervoort, 1949 (p.3); Bradford, 1971 (p.32); Roe, 1975 (p.305); Park, 1978 (p.108); Bradford & Jillett, 1980 (p.14); Razouls, 1982 (p.228); 1993 (p.309); Markhaseva, 1996 (p.11, Rem.) | | Rem.: | type: S. arcuatus Vervoort,1949. Cf. Aetideus | | | | (28) Sursamucro Bradford, 1969 ( Aetideinae ) | |
| | Ref.: | Bradford, 1969 b (p.488, 491); Bradford & Jillett, 1980 (p.77, Def.); Razouls, 1982 (p.229); 1993 (p.309); Markhaseva, 1993 (p.54 & suiv.); 1996 (p.299, Rem.); Mauchline, 1998 (p.80, 89: F); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.58) | | Rem.: | type: Sursamucro spinatus Bradford,1969. Total: 1 sp.
Defnition after Bradford & Jillett (1980, p.77) :
Female:
- Head and 1st pediger segment fused, 4th and 5th pediger segments separate.
- Posterolateral corners of last thoracic segment with points turned dorsally.
- Rostrum bluntly rounded.
- Posterior urosomal borders fringed with well defined teeth.
- A1 24-segmented, bearing long annulate setae with some sensory filaments.
- A2 exopod and endopod equal; exopod with 2 setae on segments 1 and 2.
- Mandibular palp large.
- Mx1 inner lobe 1 (arthrite) with 14 spines and setae.
- Mxp basipodal segment 1 with length twice width.
- P1 exopod with 3 external spines.
- External spines on exopods large on all legs.
- Posterior surface of endopod of P2 to P4 bear fine spinules.
- Terminal spines of P2-P4 exopods with coarse teeth without connecting lamella.
- P5 usually absent.
- Male unknown. | | | | | (29) Undeuchaeta Giesbrecht, 1888 ( Euchirellinae ) | |
| | Syn.: | Mesundeuchaeta Wolfenden, 1911 (p.244);
Euchaete : With, 1915 (p.72, 74) | | Ref.: | Giesbrecht, 1892 (p.54, 227); Giesbrecht & Schmeil, 1898 (p.33); Sars, 1900 (p.58); Esterly, 1905 (p.147); van Breemen, 1908 a (p.42, spp. Key); A. Scott, 1909 (p.60) ; Wolfenden, 1911 (p.243); Sars, 1925 (p.79); Sewell, 1929 (p.123); Wilson, 1932 a (p.60); Rose, 1933 a (p.107); Mori, 1937 (1964) (p.40); Sewell, 1947 (p.102, Rem.); Brodsky, 1950 (1967) (p.181, spp. Key); Vervoort, 1952 (n°41, p.2); 1952 h (n°49, p.3); Tanaka, 1957 b (p.199); Park, 1978 (p.182, Rem: synonymies); Bradford & Jillett, 1980 (Def.,p.80); Razouls,1982 (p.216); Gardner & Szabo, 1982 (p.247); Vaupel Klein, 1984 a (p.52, Rem.); Mauchline, 1988 (p.730); Razouls, 1993 (p.309); Markhaseva,1993 (p.54 & suiv.); 1996 (p.301, spp. Key, Rem.); Chihara & Murano, 1997 (p.689, spp. Key); Mauchline, 1998 (p.88: F; p.93: M); Bradford-Grieve & al., 1999 (p.919, 923: spp. Key); Boxshall & Halsey, 2004 (p.58: F; 61: M); Vives & Shmeleva, 2007 (p.603, spp. Key) | | Rem.: | type: Undeuchaeta major Giesbrecht,1888. Total: 5 spp. (of which 1 doubtful) + 1 sp.
Diagnosis after Tanaka (1957 b, p.199) :
- Head and 1st pediger segment fused, 4th and 5th pedigers fused.
- Head with or without crest.
- Posterolateral corners of last thoracic segment bluntly produced in female; rounded in male.
- Rostrum 1-pointed.
- Caudal rami short; appendicular seta short.
- A1 23-segmented.
- Endopod of A2 more than half the length of exopod.
- Mx1 exopod short, number of seta on endopod unreduced.
- Mx2 as in Aetideus.
- Mxp endopod very short.
- P2 endopod 1-segmented.
- P5 absent in female.
- P1 exopod 3-segmented.
- P5 male biramose.
Definition after Bradford & Jillett (1980, p.80):
Female:
- Head and pediger segment 1 indistinctly separate, pediger segments 4 and 5 fused.
- Forehead rounded or crested.
- Posterolateral corners of last thoracic segment more or less asymmetrical, rounded or triangular with rounded apex.
- Genital segment asymmetrical with ventral/or dorsal spine.
- Rostrum small, one-pointed.
- A1 23-segmented.
- A2 endopod more than half length of exopod.
- Mx1 endopod with 15 setae, basipodal segment 2 with 5 setae.
- P1 exopodal segments 1 and 2 partially separate, segment 1 without external spine.
- Endopod of P2 1-segmented.
- Coxa of P4 with or without internal edge spines; if present spines do not protrude posteriorly as in Pseudochirella;
Male:
- Body slender.
- P5 biramous on both sides; basal segments swollen, endopods 1-segmented; left leg exopod 3-segmented, apical segment with styliform distal portion with patch of hairs at midlength.
Diagnosis after Vives & Shmeleva (2007, p.603):
Female:
- Head and 1st pediger fused, sometimes with line of separation, 4th and 5th pedigers fused.
- Prosome longer than 3 times the urosome.
- A1 extenends to end of 3rd urosomal segment.
- Genital segment asymmetrical, sometimes with spicules on the margin posterior dorsal.
- P1 exopod with 2 or 3 segments and P2 to P4 with 3 segments.
- P1 endopod with 1 segment, P2 with 1 or 2 segments and P3-P4 with 3 segments.
-P5 absent.
Male:
- Geenerally without crest, but if exists, very small.
- Segmentation of prosome as in the female.
- Posteroangles of last thoracic segment slightly asymmetrical.
- Urosme 5-segmented.
- P5 biramous, asymmetrical and longer than urosome. the left leg longer than the right; endopod 1-segmented, exopod 3-segmented; right leg exopod 2-segmented. | | Remarks on dimensions and sex ratio: | | The mean females size is 4.780 mm (n = 9; SD = 1.3009), and the mean male size is 4.025 mm (n = 8; SD = 1.030). The size ratio (Male:Female) is 0.842. The sex-ratio is 1. | | | | | Undinopsis Sars, 1902 ( Aetideinae ) | | Ref.: | Schneider,1884 (nom. nudum ); Sars, 1902 (1903) (p.31); With, 1915 (p.73); Sewell, 1929 (p.99); Rose, 1933 a (p.91); Brodsky, 1950 (1967) (p.143, 150); Tanaka, 1957 a (p.44); Matthews, 1964 (p.18); Bradford & Jillett, 1980 (p.21); Razouls, 1982 (p.167); Alvarez, 1984 (p.96); Razouls, 1993 (p.309); Markhaseva, 1996 (p.68) | | Rem.: | type: Undinopsis bradyi Sars,1902 Cf. Bradyidius | | | | (30) Valdiviella Steuer, 1904 (? Aetideidae, Aetideinae ) | |
| | Ref.: | Steuer, 1904 (p.593, 596); A. Scott, 1909 (p.77); Wolfenden, 1911 (p.12, 247); With, 1915 (p.154); Sars, 1925 (p.98); Sewell, 1929 (p.133, Rem.); Rose, 1933 a (p.124); Sewell, 1947 (p.109, Rem.); Brodsky, 1950 (1967) (p.196, 223); Tanaka, 1957 b (p.204); Vervoort, 1957 (p.86,87, Rem.) (p.204); Owre & Foyo, 1967 (p.50); Heptner, 1971 (p.112, Rem.); Razouls, 1972 (Annexe: p.39: Rem.); Zvereva, 1975 (p.1893, Rem.); Park, 1978 (p.191); Bradford & Jillett, 1980 (Def., p.83); Razouls, 1982 (p.231); Mauchline, 1988 (p.729); Razouls, 1993 (p.310); Markhaseva, 1996 (p.8: Rem.); Chihara & Murano, 1997 (p.690); Mauchline, 1998 (p.93: F, M); Bradford-Grieve & al., 1999 (p.924, 927: clé spp.; dans la famille des Euchaetidae ); Boxshall & Halsey, 2004 (p.57: F; 60: M); Vives & Shmeleva, 2007 (p.606, spp. Key) | | Rem.: | Type: Valdiviella oligarthra Steuer,1904. Total: 7 spp.
For Tanaka (1957 b, p.205) the genus Valdiviella was created by Steuer (1904) to accomodate a new species (V. oligarthra which is closely related to Euchaeta and also to some genera of Aetideidae.
According to Steuer, the genus has the mandible similar in structure to those of Gaetanus and Chiridius; Mx1 as in Undeuchaeta; Mx2 as in Aetideus, thus the genus Valdiviella is regarded as a member of the family Aetideidae, whereas Sars (1925, p.98) doubts of this justification. A. Scott states that ''the genus is closely related to Euchaeta, but easily recognised by its bifurcate rostrum; the incomplete segmentation of the exopodites of P1 and P2, and indistinctly 3-segmented endopodites of P3 and P4''.
- The genus has general appearance as in Euchaeta; head and 1st thoracic segment fused, 4th and 5th thoracic segments fused; posterior margin of last thoracic segment rounded; abdomen 4-segmented; appendicular seta short; endopod of A2 about as long as exopod; endopod of Md about as long as exopod;
- Mx1 differs from that of Euchaeta : setae on endopod and 2nd basal segment much reduced, endopod with only 3 setae, 2nd basal with 3 setae whereas that of Euchaeta with 3+3+3 setae on endopod, and 5 on 2nd basal segment;
- Mx2 resembles that of Aetideidae, setae on 5th lobe strong; Mxp resembles that of Undeuchaeta and Euchaeta, but row of spinules on anterior margin of 2nd basal segment are situated on the proximal half, wherea, it is on the distal half in Euchaeta, and thet are feeble.
- P1 as in Euchirella : 1st and 2nd exopodal segments fused; endopod of P2 1-segmented, that of P3 2-segmented; P4 exopod and endopod 3-segmented each; P5 absent in female; male P5 resemble those of Chirundina. The genus has a closer relationship to Aetideidae : rostrum resembles that of Aetideopsis or Undinopsis in its bifurcate shape;
Mx1 has the reduced number of setae on endopod and on 2nd basal segment.
- Mx2 as those of Aetideus in having 1 strong seta on 5h lobe (= endite).
- P1 similar to those of Euchirella.
- P2 as in Euchirella or Undeuchaeta.
- The genus has resemblance to Euchaeta: general appearance, structure of A1, Md and Mxp. Thus Valdiviella has the generic characters common both Aetideidae and Euchaetidae; it is intermediate form between Undeuchaeta and Euchaeta. For the author it seems reasonable, as Steuer stated, to accomodate it in the family Aetideidae.
For Vervoort (1957, p.87) the genus, in many respects, takes an intermediate position between the Aetideidae and the Euchaetidae. It approaches the Aetideidae in the structure of the rostrum, swimming legs, in the case of males, the structure of P5. The structure of Mxp shows unmistakable affinities with the Euchaetidae.
The belonging of this genus to this family is discussed by Heptner (1971) and by Zvereva (1975).
Definition after Bradford & Jillett (1980, p.83) Female:
- General appearance as in Euchaeta.
- Rostrum bifurcate.
- Head and pediger segment 1 fused, 4th and 5th pediger segments fused..
- Posterolateral corners of last thoracic segment rounded.
- A1 23-segmented (as in Euchaeta).
- A2 exopod and endopod equal in length.
- Mx1 with reduced setattion on inner lobe 1 (arthrite), endopod and basipodal segment 2 (11, 3, 3 setae respectively); posterior surface of inner lobe 1 with only 1 seta.
- Mx2 as in Aetideus, seta on lobe 5 strong.
- Mxp as in Undeuchaeta and Euchaeta, but row of spinules on anterior margin of basipodal segment 2 situated proximally not distally as in Euchaeta
- Swimming legs with some degree of fusion between proximal segments.
- P5 absent. | | Remarks on dimensions and sex ratio: | | The mean female size is 7.278 mm (n = 11; SD = 2.4288), and the mean male size is 6.822 mm (n = 8; SD = 1.9816). The size ratio (Male:Female) is 0,937. Le sex-ratio (F:M) is 1,2. | | | | | Wilsonidius Tanaka, 1969 ( Aetideinae ) | |
| | Ref.: | Tanaka, 1969 (p.266); Bradford & Jillett, 1980 (p.92); Razouls, 1982 (p.230); 1993 (p.310); Markhaseva, 1996 (p.8, 129, Rem.); Mauchline, 1998 (p.92: F) | | Rem.: | type:Wilsonidius alaskaensis Tanaka,1969. Cf.: Chirundina | | | | | | | | | | | | | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 22, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|



;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
