|
|
 |
|
Cyclopoida ( Order ) |
|
|
|
Oithonidae ( Family ) |
|
|
|
Oithona ( Genus ) |
|
|
| |
Oithona nana Giesbrecht, 1892 (F,M) | |
| | | | | | | Syn.: | ? Oithona helgolandica Claus, 1863 (p.105);
Oithonina nana : Sars, 1913 (1918) (p.5); Wilson, 1932 a (p.316, figs.F); 1942 a (p.197); 1950 (p.271); Fagetti, 1962 (p.40); Haq, 1965 a (p.555, figs. Nauplius, C1, key of Nauplius); Harder, 1968 (p156, Table 1, behaviour vs density discontinuity); Shih & al., 1971 (p.56);
Non Oithona nana : Yamazi, 1955; Hirota & Hara, 1975.
Oithona plumosa Lindberg, 1947; 1950 e (p.265); Wellershaus, 1970 (p.480); Shuvalov, 1980 (p.152, figs.F);
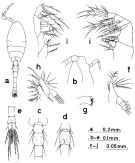
O. plumosa pseudoplumosa Lindberg, 1950 e (p.265) | | | | Ref.: | | | Giesbrecht, 1892 (p.538, 549, 774, figs.F,M); Karavaev, 1894 (p.58, figs.F, Rem.); Thompson & Scott, 1903 (p.236, 255); Esterly, 1905 (p.209, figs.F,M); Burckhardt, 1913 (p.425); Rosendorn, 1917 a (p.40, figs. F,M); Pesta, 1920 (p.552); Murphy, 1923 (p.449, figs., key juv. stages); Sewell, 1924 (p.791); Früchtl, 1924 b (p.70); Gurney, 1927 (p.159, Rem.: 2 forms); Mori, 1929 (p.199, figs.F); Kiefer, 1929 g (p.9, Rem.F,M); Sewell, 1933 (p.30); Dakin & Colefax, 1933 (p.207); Rose, 1933 a (p.281, figs.F,M); Mori, 1937 (1964) (p.113, figs.F,M); Sewell, 1947 (p.254, Rem.); Marques, 1951 a (p.24); Carvalho,1952 a (p.167, figs.F); Marques, 1958 a (p.134); Tanaka, 1960 (p.59, figs.F,M, Rem.); Grice, 1960 a (p.487, figs.F,M); Vilela, 1965 (p.12); Gonzalez & Bowman, 1965 (p.272, figs.F,M); Vilela, 1968 (p.29); Ramirez, 1969 (p.88); Corral Estrada, 1970 (p.213, Rem.); Wellershaus, 1970 (p.479); Sazhina & Kovalev, 1971 (p.1100, fig.F); Björnberg, 1972 (p.88, figs., Rem.N); Razouls, 1972 (p.95, Annexe: p.104); Chen & al., 1974 (p.33, figs.F,M); Marques, 1975 (p.50); Kos, 1976 (Vol.II, figs.F,M, Rem.); Shuvalov, 1976 (in Kos, 1976, Vol. II, figs.F), Rem.); Nishida & al., 1977 (p.138, figs.F,M, Rem.); Dawson & Knatz, 1980 (p.9, figs.F,M); Shuvalov, 1980 (p.146, figs.N,F,M); Ferrari & Bowman, 1980 (p.14, figs.F,M); Nishida, 1985 a (p.30, 52, Redescr. F,M, figs.F,M, Rem.: 2 forms, p.139); Björnberg & al., 1981 (p.665, figs.F,M); Dussart & Defaye, 1985 (p.10); Sazhina, 1985 (p.85, figs.N); Nishida, 1985 a (p.52, figs.F,M); 1986 (p.386); Huys & Boxshall, 1991 (p.205, 442, 465, figs.F,M); Yoo & Lim, 1993 (p.91, 95, , Table 1, fig.2, Key: p.100); Kim & al., 1993 (p.271); Chihara & Murano, 1997 (p.937, Pl.198,203: F,M); Bradford-Grieve & al., 1999 (p.886, 966, figs.F); McKinnon, 2000 (p.108, Rem.); Conway & al., 2003 (p.203, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.612: figs.F); Conway, 2006 (p.21, copepodides 1-6, Rem.); Avancini & al., 2006 (p.123, Pl. 91, figs.F,M, Rem.); Vives & Shmeleva, 2010 (p.65, figs.F,M, Rem.); Al-Yamani & al., 2011 (p.96, 98, figs.F,M); Gubanova & al., 2013 (in press, Rem: p.2, 5, Table 1, 2, 4, fig.2); Wend-Heckmann, 2013 (p.114, 117, 126, Table 3, 4, 7.10); Cornils & Wend-Heckmann, 2015 (p.243, Table 1, Fig. 2: molecular biology); Madoui & al., 2017 (p.4467, genome). |  issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.15, Fig.8]. Female (from Caribbean area): a, habitus (lateral right side); b, urosome (lateral right side); c, P4. Nota: Prosome/Urosome = 1.1. Endopodal segment 1 of P4 with a distinct row of 4 long spines Endopodal segment 2 of P4 with both setae mofified, neither seta strongly curved, both with flange on distal 3/5; endopodal segment 3 of P4 proximal seta similar, with flange on distal 3/5. P5 elongate , with several long hairs dorsal to it on posterior margin of urosomal segment 1. Knob near genital opening armed with short, thick spine and longer seta. Male: d, habitus (lateral right side); e, cephalosome and flap; f-g, left and right \"pore signature\" of second male; h-i, same of third male. Nota: Prosome/Urosome = 1.3. Flap of cephalosome relatively broader and shorter than in O. hebes and O. fonsecae; Anterodorsal cluster of \"pore signature\" roughtly circular. Intraspecific variation in the \"pore signature\" has been observed in this species.
|
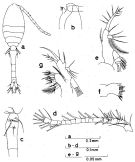 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.56, Fig.27]. As Oithona nana typical form. Female: a, habitus (dorsal); b, forehead (lateral); c, thoracic segment 5 and genital segment (lateral left side); d, A1; e, Md (mandibular palp); f, Md (biting edge); g, mx1.
|
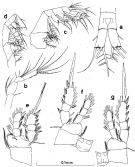 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.57, Fig.28]. As Oithona nana typical form. Female: a, anal segment and caudal rami (dorsal); b, A2; c, Mx2; d, Mxp; e, P1; f, P2; g, P3.
|
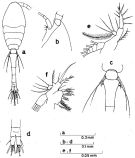 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.58, Fig.29]. As Oithona nana \"plumosa\" form. Female: a, habitus (dorsal); b, forehead (lateral); c, last thoracic segments and genital segment (dorsal); d, anal segment and caudal rami (dorsal); e, Md (mandibular palp); f, Mx1.
|
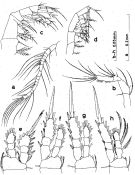 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.59, Fig.30]. As Oithona nana \"plumosa\" form. Female: a, A1; b, A2; c, Mx2; d, Mxp; e, P1; f, P2; g, P3; h, P4. Nota: after Vives & Shmeleva (2010, p.67): Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 3; 1, 1, 4. P2: 1, 1, 3; 1, 1, 5 P3: 1, 1, 3; ; 1, 1, 5. P4: 1, 1, 2; 1, 1, 5.
|
 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.61, Fig.31]. As Oithona nana \"plumosa\" form. Male: a, habitus (dorsal); b, forehead (lateral); c, last thoracic segments and genital segment (dorsal); d, thoracic segment 5 and genital segment (ventral); ; e, last urosomal segments and caudal rami (dorsal); f, Md (mandibular palp); g, Md (biting edge); h, Mx1; i, Mx2; j, Mxp.
|
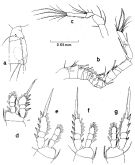 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.62, Fig.32]. As Oithona nana \"plumosa\" form. Male: a, thoracic segment 5 and genital segment (lateral right side); b, A1; c, A2; d, P1; e, P2; f, P3; g, P4.
|
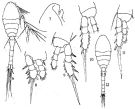 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.2, Figs.6-12]. Female (from China Seas): 6, habitus (dorsal); 7, forehead (lateral); 8, P1; 9, P2; 10, P3; 11, P4. Male: 12, habitus (dorsal).
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.209, Fig.51]. Female (from San Diego): b, 3rd segment of exopod of P1. Male: a, habitus (dorsal); c, 3rd segment of exopod of P4.
|
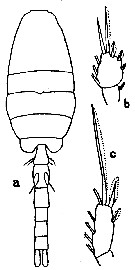
 issued from : G.D. Grice in Bull. mar. Sci. Gulf Caribb., 1960, 10 (4). [p.486, Figs.7-11]. Female (from NE Gulf of Mexico: off Alligator Harbor): 7, habitus (dorsal); 8, forehead (lateral); 9, P4. Nota: Rostrum absent. Exopod segments of P4 with 1, 1, 2 spines. Male: 10, habitus (dorsal); 11, basipodal segment 2 of Md. Nota: Exopod segments of P1 with 1, 1, 2 spines; 2nd basipodal segment of Md with 1 strong and 1 fine seta.
|
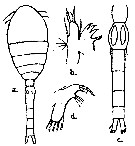 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. \"Valdiviella\", 1917, 23. [p.41, Fig.24]. Female: a, habitus (dorsal); b, Mx1. Nota: Proportion of lengths (p.cent) Prosome : 54.72, Urosome : 45.28 . Relative lengths of urosomal segments and caudal rami: 10 : 33 : 17 : 17 : 13 : 13. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae ; Si = inner setae), P1 : 1, 1, 3 Se ; 1, 1, 4 Si ; P2 : 1, 1, 3 Se ; 1, 1, 5 Si ; P3 : 1, 1, 3 Se ; 1, 1, 5 Si ; P4 : 1, 1, 1 Se ; 1, 1, 5 Si . Male: c, urosome; d, Md. Nota: Proportion of lengths (p.cent) Prosome : 56.82, Urosome : 43.18 . Relative lengths of urosomal segments and caudal rami: 7 : 10 : 9 : 7: 6 : 4 : 4. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae ; Si = inner setae), P1 : 1, 1, 3 Se ; 1, 1, 4 Si ; P2 : 1, 1, 3 Se ; 1, 1, 5 Si ; P3 : 1, 1, 3 Se ; 1, 1, 5 Si ; P4 : 1, 1, 2 Se ; 1, 1, 5 Si .
|
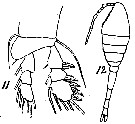 issued from : T. Mori in Zool. Mag. Tokyo, 1929, 41 (486-487). [Pl. VII, Figs. 11-12]. Female (from Chosen Strait, Korea-Japan); 11, P1; 12, habitus (dorsal).
|
 issued from : T. Mori in The Pelagic copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl. 63, Figs.1, 3-8]. Female: 1, habitus (dorsal); 3, Md; 4, P1; 5, P2; 6, P3; 7, P4; 8, Mx1.
|
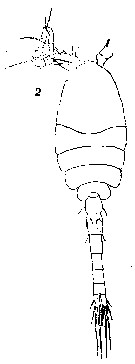 issued from : T. Mori in The Pelagic copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl. 63, Fig.2]. Male: 2, habitus (dorsal).
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.147, Fig.41]. Female: A-B, habitus (dorsal and lateral, respectively); C, habitus (dorsal); D, forehead (dorsal); E, cephalon (lateral); F, 4th, 5th toracic segments and genital segment (dorsal); G, posterior part of abdomen and caudal rami; H, P1; I, P2; J, P3; K, P4.
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.153, Fig.44]. As Oithona plumosa. After Lindberg, 1950. Female: A, habitus (dorsal); B, forehead (dorsal); C, same (ventral); D, same (lateral); E, A1; F, A2; G, Md; H, posterior part of urosome.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.44, Figs.2, 4]. Male: 2, habitus (dorsal); 4, forehead (lateral). Ce = cephalosome.
|
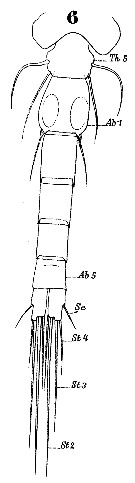 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.44, Fig.6]. Male: 6, urosome (dorsal).
|
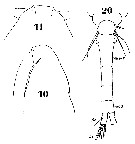 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.10, 11, 20]. Female: 10, head (lateral); 11, forehead (dorsal); 20, urosome (dorsal). Th5 = thoracic segment 5; Ab 1-2 = genital double-somite; Ab 5 = anal somite; Se = outer marginal seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.24, 26]. Female: 24, Md; 26, Mx1. B2 = basis; Ri = endopodite; Li 3 = inner lobe 3.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.34, 35, 42]. Female: 34, P1; 35, P2; 42, P4.
|
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.138, Fig.11]. Female (from Suruga Bay and adjacent waters): a, habitus (dorsal), b, forehead (lateral); c, Md; d, Mx1; e, P1; f, P2; g, P3; h, P4. Nota: Setae formula of exopod P1 to P4 (outer setae in first, inner setae in second). P1: 1,1,3; 1,1,4. P2: 1,1,3; 1,1,5. P3: 1,1,3; 1,1,5. P4: 1,1,2; 1,1,5. Prosome length/urosome length = 1.04-1.25.
|
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.138, Fig.12]. Male (from Suruga Bay and adjacent waters): a, habitus (dorsal), b, forehead (lateral); c, Md; d, Mx1; e, P1; f, P2; g, P3; h, P4. Nota: Exopod of P1-P4 armed as in the females. Prosome length/urosome length = 1.38-1.43.
|
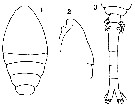 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [Pl. XXVI, 1-3]. Female (from Indian Ocean): 1, Cephalothorax (dorsal); 2, head (lateral); 3, urosome (dorsal). Nota: Prosome and urosome in proportional lengths 52 to 48. Rostrum absent. urosomal segments and caudal rami in the proportional lengths 12 : 32 : 17 : 15 : 11 : 13 = 100. Caudal rami 2 times as long as wide. Genital segment produced laterally about on the proximal 1/5 of the segment.
|
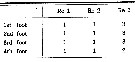 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [p.60]. Female: Number of seta on the exopodal segments in the swimming legs P1 to P4. Nota: The terminal spine of the exopod of the P1 to P4 are longer than the 3rd segment of the exopod of the respective leg.
|
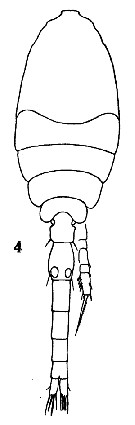 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [Pl. XXVI, 4]. Male: 4, habitus (dorsal). Nota: Prosome and urosome in proportional lengts 54 to 46. Urosomal segments and caudal rami in the proportional lengths 13 : 21 : 17 : 16 : 13 : 10 : 10 = 100.
|
 Issued from K.-I. & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2). [Key of female]: Morphological characters of Oithona nana female in Korean waters :
1 - Anterior part of prosome rounded in dorsal view.
2 - Rostrum blunt. 3 - P5 with 1 seta. 4 - Length of A1 same as that of prosome. 4 - Length of 1st spine in inner lobe 1 of Mx1 same as that of other spines.
|
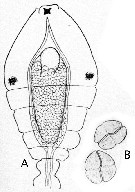 Issued from : E. Chatton in Arch. Zool. Exp. & Gen., 1920, 59, [p.147, Fig.47]. Oithona nana parasited by Blastodinium oviforme (Dinoflagellate). A, Ovoid-like, diblastic individual; B, two sporocysts.
|
 Issued from : A. Cornils & B. Wend-Heckmann in Helgol. Mar. Res., 2015, 69. [p.246, Table 1]. Oithona nana charcteristics in the semi-enclosed bight in the northern Germann Wadden Sea (North Sea). Compare with O. similis, O. plumifera, O. atlantica and O. davisae in the North Sea.
|
 Issued from : K.-I. Yoo & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2) [p.99, Fig.2, M] Female (from Korean Waters): M, forehead and rostrum (lateral view).
|
 Issued from : R. Gurney in Trans. zool. Soc. London, 1927, 22. [p.160]. Lengths and ratio between the two forms of Oithona nana in the Suez Canal (northern and Southern, Port Said to Ras-el Ech, respectively).
|
 Issued from : C. Castellani, C. Robinson, T. Smith & R.S. Lanpitt in mar. Ecol/ Progr. Ser., 2005, 285. [p.132, Table 1]. Oithona spp. Summary of respiration rates (range) reported in the litterature in different regions. c: converted to dry weight from wet weight assuming 81% water content (Mauchline, 1998). Cf. Oithona similis to compare to other species in Table 1 in the same authors.
| | | | | Compl. Ref.: | | | Cleve, 1904 a (p.192); Carazzi & Grandori, 1912 (p.14, 38); Chatton, 1920 (p.17); Rose, 1925 (p.152); Massuti Alzamora, 1942 (p.102, Rem.); Sewell, 1948 (p.367, 461, 487); Fleury, 1950 (p.47, fig.2); Hansen K.V., 1951 (p.231, migration vs discontinuity layer); Grice, 1956 (p.61); Yamazi, 1958 (p.153, Rem.); Gaudy, 1962 (p.93, 99, Rem.: p.116) ; Ganapati & Shanthakumari, 1962 (p.10, 16); Shmeleva, 1963 (p.141); Duran, 1963 (p.25); Giron-Reguer, 1963 (p.56); Gaudy, 1963 (p.30, Rem.); Björnberg, 1963 (p.75, Rem.); Unterüberbacher, 1964 (p.32); De Decker, 1964 (p.16, 23, 29, 32); De Decker & Mombeck, 1964 (p.13); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Neto & Paiva, 1966 (p.30, Table III, annual cycle); Pavlova, 1966 (p.44); Chakroun, 1966 (p.67, Tableau); Mazza, 1966 (p.73); 1967 (p.363); Ehrhardt, 1967 (p.742, geographic distribution, Rem.); Delalo, 1968 (p.138); Porumb, 1968 (p.417); Dimov, 1968 (p.506); Kovalev, 1968 a (p.441, fig.1); Emery, 1968 (p.293, swarm); Champalbert, 1969 a (p.642); El-Maghraby & Dowidar, 1970 (p.81); Dowidar & El-Maghraby, 1970 (p.269); Reeve, 1970 (p.894, Rem.: p.908); Carli, 1971 (p.372, tab.1); Paulmier, 1971 (p.168); Salah, 1971 (p.320); Apostolopoulou, 1972 (p.329, 373); Della Croce & al., 1972 (p.1, fig.2, Rem.); Valentin, 1972 (p.349, figs.9, 10, 13, 14, 17, 19, Table 1, 2, egg production); Binet, 1973 (p.77); Desgouille, 1973 (p.1, 131, Rem.: p.136, fig.16); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Hirota, 1974 (p.1, Table 3, 4, fig.4, 5, 6); Hirota & Hara, 1975 (p.115, fig.5); Vives & al., 1975 (p.53, tab.II, III, IV); Porumb, 1976 (p.91); Lakkis & Abboud, 1976 (p.81); Newbury & Bartholomew, 1976 (p.373, production); Gaudy, 1976 (p.77, fig.1, 7, Table I, II, III, production); Furuhashi, 1976 (p.355, Diel vertical migration); Hanaoka, 1977 (p.267, 301, abundance); Ferrari, 1977 (p.410); Boxshall, 1977 b (p.553); Grindley, 1977 (p.341, Table 2); Lampitt, 1978 (p.1228, feeding); Dessier, 1979 (p.83, 201, 207); Vaissière & Séguin, 1980 (p.23, tab.2); Gallo, 1981 (p.847); Castel & Courties, 1982 (p.417, Table II, fig.4, spatial & monthly distribution); Vives, 1982 (p.295); Lampitt & Gamble, 1982 (p.183, respiration vs. food); Kovalev & Shmeleva, 1982 (p.85); Dessier, 1983 (p.89, Tableau 1, Rem., %); Scotto di Carlo & al., 1984 (p.1045); Tremblay & Anderson, 1984 (p.7, Rem.); Dussart, 1984 (p.301); Patriti & al., 1984 (tab.1); Regner, 1985 (p.11, Rem.: p.40); ); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea); Jansa, 1985 (p.108, Tabl.I, II, III, IV, V); Garcia-Rodriguez, 1985 (p.37); 1985 a (p.41, 42); Brinton & al., 1986 (p.228, Table 1); M. Lefèvre, 1986 (p.33); Diouf & Diallo, 1987 (p.260); Ceccherelli & al., 1987 (p.571, fig.5); Lozano Soldevilla & al., 1988 (p.61); Krsinic & Vilicic, 1989 (p.12, tab.3); Krsinic, 1990 (p.337, Table III, vertical distribution)Dai & al., 1991 (tab.1); Yoo, 1991 (tab.1); Jouffre & al., 1991 (p.489, lagoon); Santos & Ramirez, 1991 (p.83); Huntley & Lopez, 1992 (p.201, Table A1, egg-adult weight, temperature-dependent production); Seguin & al., 1993 (p.23); Guerin-Ancey & David, 1993 (p.119, table 1, biovolume, vertical distribution); Santos & Ramirez, 1995 (p.133, Tabl. I, fig.2, 3); Hwang & Turner, 1995 (p.207, Table 1, 2, fig. 1, 2, 3: swimming behaviour); Shih & Young, 1995 (p.75); Krause & al., 1995 (p.81, Rem.: p.134); Böttger-Schnack, 1995 (p.92); 1996 (p.1088); Kotani & al., 1996 (tab.2); Hopcroft & Roff, 1996 (p.789, diel egg production); Fernandez de Puelles & al., 1996 (p.97, occurrence: p.101); Böttger-Schnack, 1997 (p.410); Park & Choi, 1997 (Appendix); Sharaf & Al-Ghais, 1997 (tab.1); Noda & al., 1998 (p.55, Table 3, occurrence); Hure & Krsinic, 1998 (p.76, 103); Gilabert & Moreno, 1998 (tab.1,2); Hopcroft & al., 1998 (tab.2); Alvarez-Cadena & al., 1998 (t.4); Suarez-Morales & Gasca, 1998 a (p.111); Omori & Norman, 1998 (p.279, Rem.: anthropogenic short-term effects); Siokou-Frangou, 1999 (p.478); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Lopes & al., 1999 (p.215, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Dolganova & al., 1999 (p.13, tab.1); El-Serehy, 1999 (p.172, Table 1, occurrence); Ueda & al., 2000 (tab.1); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Seridji & Hafferssas, 2000 (tab.1); Holmes Gotto, 2000 (p.4, Rem.); Sautour & al., 2000 (p.531, Table II, abundance); d'Elbée, 2001 (tabl.1); Richard & Jamet, 2001 (p.957: Rem.); Fransz & Gonzalez, 2001 (p.255, tab.1); El-Serehy & al., 2001 (p.116, Table 1, 3, 4: abundance vs transect in Suez Canal); Bressan Moro, 2002 (tab.2); Sameoto & al., 2002 (p.13); Zerouali & Melhaoui, 2002 (p.91, Tableau I, figs.5, 8); Dunbar & Webber, 2003 (tab.1); Vukanic, 2003 (p.139, tab.1); McKinnon & al., 2003 (p.101, tab.2, fig.11); Zagorodnyaya & al., 2003 (p.52); Rezai & al., 2004 (p.486, tab.2, 3, abundance); Vieira & al., 2003 (p.S163, Table 2, abundance); Chang & Fang, 2004 (p.456, tab.1); Daly Yahia & al., 2004 (p.366, fig.4, tab.1); Fernandez de Puelles & al., 2004 (p.654, fig.7); Marrari & al., 2004 (p.667, tab.1); Vukanic & Vukanic, 2004 (p.361, tab.3, fig.3); Guermazi & al., 2005 (p.280); Mazzochi & al., 2005 (p.155); Licandro & al., 2005 (p.153); Lakkis & al., 2005 (p.152); Obuid Allah & al., 2005 (p.123, occurrence % vs metal contamination); Alvarez-Silva & al., 2005 (p.39); Rawlinson & al., 2005 (p.205, tidal exchange); Prusova & Smith, 2005 (p.76, 78); Zuo & al., 2006 (p.164: tab.1); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Isari & al., 2006 (p.241, tab.II); Marques & al., 2006 (p.297, tab.III); Mageed, 2006 (p.471, Tabl.2, 4); Williams & Muxagata, 2006 (p.1055); Atienza & al., 2006 (p.221, metabolism); Albaina & Irigoien, 2007 (p.435: Tab.1); Porri & al., 2007 (p.714); David & al., 2007 (p.59: tab.2); Busatto, 2007 (p.26, Tab.2); Valdés & al., 2007 (p.104: tab.1); Krsinic & al., 2007 (p.528); Khelifi-Touhami & al., 2007 (p.327, Table 1); Leandro & al., 2007 (p.215); Zakaria, 2007 (p.238, Table 2, spatial distribution); Zakaria & al., 2007 (p.52, Rem.: p.58, Table 1); McKinnon & al., 2008 (p.843: Tab.1, p.846: Tab.II, fig.7); Isinibilir & al., 2008 (p.745: Tab.1); Cabal & al., 2008 (289, Table 1) ; Humphrey, 2008 (p.84: Appendix A); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.514); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Selifonova & al., 2008 (p.305, Tabl. 2); Shmeleva & al., 2008 (p.31, Table 1); Rossi, 2008 (p.90: Tableau XII); Tseng L.-C. & al., 2008 (p.153, fig.5, Table 2, occurrence vs geographic distribution, indicator species); Ohtsuka & al., 2008 (p.115, Table 5); Miyashita & al., 2009 (p.815, Tabl.II); Brylinski, 2009 (p.253: Tab.1, p.256: Rem.); Hafferssas & Seridji, 2010 (p.353, Table 2); Lidvanov & al., 2010 (p.356, Table 3); Drira & al., 2010 (p.145, Tanl.2); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Hidalgo & al., 2010 (p.2089, fig.2, 4, Table 2, cluster analysis); Mazzocchi & Di Capua, 2010 (p.428); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); Isinibilir, 2010 (p.233, abundance %); Hafferssas & al., 2010 (p.1281, Table III, abundance vs spatial distribution); Fazeli & al., 2010 (p.153, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I); Viñas & al., 2010 (p.177, body dimensions vs bivolume); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Saitoh & al., 2011 (p.85, Table 4, 5); El-Sherbiny & al., 2011 (p.837, seasonal abundance, Table 3: correlations), Magris & al., 2011 (p.260, abundance, interannual variability); Moscatello & al., 2011 (p.80, Table 4); S.C. Marques & al., 2011 (p.59, Table 1); Di Mauro & al., 2011 (p.69, biovolume); Mazzocchi & al., 2011 (p.1163, Table II, fig.6, long-term time-series 1984-2006); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Temperoni & al., 2011 (p.729, Seasonal variation, production); Tutasi & al., 2011 (p.791, Table 2); Tseng L.-C. & al., 2011 (p.239, Table 3, abundance %); Cepeda & al., 2012 (p.1, figs.1, 3, Table 1, Molecular systematic); Van Ginderdeuren & al., 2012 (p.3, Table 1); Delpy & al., 2012 (p.1921, Table 2, fig.7); Mazzocchi & al., 2012 (p.135, annual abundance 1984-2006); Shiganova & al., 2012 (p.61, fig. 4, 5); Glushko & Lidvanov, 2012 (p.138, Tableau 1); Uysal & Shmeleva, 2012 (p.909, Table I); Salah S. & al., 2012 (p.155, Tableau 1); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Rekik & al., 2012 (p.336, Table 1, fig.7, abundance); Zizah & al., 2012 (p.79, Tableau I, Rem.: p.86); Gonçalves & al., 2012 a (p.16, abundance, diel vertical behavior); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Menéndez & al., 2012 (p.389, Table 1: seasonal abundance); Vidjak & al., 2012 (p.243, Rem.: p.253); Aubry & al., 2012 (p.125, table 3, fig. 8 a, interannual variation); Jean & al., 2012 (p.12, Table 3, protein vs environmental metal stress); Johan & al., 2012 (p.647, Table 1, 2, fig.2, salinity range); Krsinic & Grbec, 2012 (p.57, 61: abundance); Spinelli & al., 2012 (p.39, potential prey for fish); Lavaniegos & al., 2012 (p. 11, Appendix); Dorgham & al., 2012 (p.473, Table 3: abundance %); Hidalgo & al., 2012 (p.134, Table 2) ; Zamora Terol, 2013 (p.174: Tableb 5); ); Zamora-Terol & Saiz, 2013 (p.376, Rem.: Table 3, egg production); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Gubanova & al., 2013 (in press, p.4, Table 2, Rem.); Mihneva & Stefanova, 2013 (p.119: Rem ); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Melo Jùnior & al., 2013 (p.363, Table 2, egg production); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Jagadeesan & al., 2013 (p.27, Table 3, seasonal variation); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Jungbluth & Lenz, 2013 (p.630, Table I); Alvarez-Silva & Torres-Alvarado, 2013 (p.241, Table1: seasonal abundance); Terbiyik Kurt & Polat, 2013 (p.1163, Table 2, seasonal distribution); Lidvanov & al., 2013 (p.290, Table 2, % composition) ; Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Mazzocchi & al., 2014 (p.64, Table 5, abundance) ; Fierro Gonzalvez, 2014 (p.1, Tab. 3, 5, occurrence, abundance); Zaafa & al., 2014 (p.67, Table I, occurrence); Aboul Ezz & al., 2014 (p.283, Rem.: p.284, 287); Zakaria, 2014 (p.3, Table 1, abundance vs 1960-2000); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance) ; Zakaria & al., 2016 (p.1, Table 1, 3, 4); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Vidjak & al., 2016 (p.629, Rem.); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance) ; Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1, Rem.: p.502, abundance); Ohtsuka & Nishida, 2017 (p.565, Table 22.1); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Abo-Taleb & Gharib, 2018 (p.139, Table 5, p.145, 149: occurrence %); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence); Terbiyik Kurt & Besiktepe, 2019 (p.1, fig.3, 4, 8, Table 2, seasonal variations, % population, spatial distribution); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions). | | | | NZ: | 21 | | |
|
Distribution map of Oithona nana by geographical zones
|
| | | | | | | | | | | | | | | 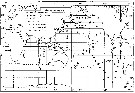 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.139, Fig.90]. Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.139, Fig.90].
Indo-Pacific geographical distribution of Oithona nana. Dotted line: AC, Arctic Convergence; SC, Subtropical Convergence. Arrows indicate stations where the "plumosa" form occurred. |
 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 39]. Oithona nana (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 39]. Oithona nana (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
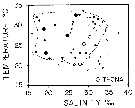 issued from : M.R. Reeve in Bull. mar. Sc., 1970, 20 (4). [p.902, Fig.4]. issued from : M.R. Reeve in Bull. mar. Sc., 1970, 20 (4). [p.902, Fig.4].
Temperature-salinity diagram for Oithona nana in Biscayne Bay (Florida, Miami). |
 issued from : A.V. Bogorov in Gidrobiol. Zh., 1968 4 (3). [p.78, Table 1]. issued from : A.V. Bogorov in Gidrobiol. Zh., 1968 4 (3). [p.78, Table 1].
Number of egg-laying from the same body size at different seasons in the Black Sea.
A: Month and year; B: Females number analysed; C: Body lengths (limits of variation; M = mean); D: Number of eggs. |
 issued from : A.V. Bogorov in Gidrobiol. Zh., 1968 4 (3). [p.79, Table 2, 3]. issued from : A.V. Bogorov in Gidrobiol. Zh., 1968 4 (3). [p.79, Table 2, 3].
Table 2: Importance of number eggs-laying by female of the same body size from the Black Sea and the Mediterranean Sea.
A: Sea; B: Month and Year; C, Body size of females (limits and M = mean); D: Number of eggs (M = mean).
Table 3: Number of eggs by female and body sizes.
A: Lengths in mm; B: Number of eggs. |
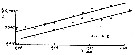 issued from : A.V. Bogorov in Gidrobiol. Zh., 1968 4 (3). [p.79, Fig.3 1]. issued from : A.V. Bogorov in Gidrobiol. Zh., 1968 4 (3). [p.79, Fig.3 1].
Relation between the body of females and the egg diameter.
Black circle: Black Sea; open circle: Mediterranan Sea and Adriatic Sea. |
 Issued from : M.D. Viñas, N.R. Diovisalvi & G.D. Cepeda in J. Braz. Oceanogr., 2010, 58 (2). [p.178, Table 1]. Issued from : M.D. Viñas, N.R. Diovisalvi & G.D. Cepeda in J. Braz. Oceanogr., 2010, 58 (2). [p.178, Table 1].
Body dimensions and estimated biovolume of Oithona nana samples on October 18th and November 11th at the permanent coastal station (38°28'S, 57°41'W).
Body wet weight can be derived from measurements of body volume by applying a factor of 1 for specific gravity. Dry weight can be obtained by mulitplyng the wet weight by 0.20 and the carbon content as 40% of the dry weight.
Prosome length, width and height, as well as urosome length and width, were measured, in order to apply the model of Chojancki & Hussein (1983) slightly modified (antenna and leg volume excluded) by Fernadez Araoz (1991). |
 Issued from : S. Zamora Terol in Tesis Doc. Univ. Politèc. Catalunya, 2013 [p.82-83, Table 3]. Issued from : S. Zamora Terol in Tesis Doc. Univ. Politèc. Catalunya, 2013 [p.82-83, Table 3].
Comparison of egg production and clutch sizes among different species of Oithona from field and lab studies.
a: body total length; b: eggs per sac.
Also, see: Oithona attenuata, O. colcarva, O. davisae, O. aruensis, O. plumifera, O. setigera, O. similis, O. simplex. |
 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2]. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2].
Table 1: Experimental protocol.
Table 2:Swimming versus non-swimming behaviour, and behavioural transitions per minute. Means (bold type), ranges, and standard deviations (SD). Number of repeated observations = 500.
Copepods collected in surface net from waters near Pitou (NE coast Taiwan: 25°N, 122°E) in daylight in July 1993.
Compare with Temora turbinata, O. similis, Oncaea venusta, Macrosetella gracilis and Undinula var. taiwanicus by the same authors. |
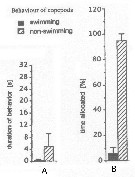 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2]. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2].
Fig.1: A, Mean durations periods (seconds) of swimming versus non swimming behaviour by Oithona nana from north-east coast of Taiwan (near Pitou) in daylight in July 1993.
Fig.2: B, mean percentages of time allocated to swimming versus non-swimming behaviour by Oithona nana. |
| | | | Loc: | | | South Africa (off Cape of Good Hope, E & W), Saldanha Bay, Namibia, Angola, Baia Farta, Rio Congo estuary, G. of Guinea, Casamance, Cape Verde Is., off Morocco-Mauritania, Cap Ghir, Canary Is., off Madeira, Portugal, Mondego estuary, Bay of Biscay, off Gironde estuary, La Pallice roadstead, Arcachon Bay, Glenans Islands, Belon estuary, Buenos Aires, Peninsula Valdés, Bahia Blanca, off Mar del Plata, Brazil (S, Rio de Janeiro, Vitoria Bay, Mucuri estuary, off Natal), Barbada Is., Caribbean Colombia (Guajira, Tayrona), Bahia de Mochima (Venezuela), Yucatan, E Costa Rica, G. of Mexico, Porto Rico, Jamaica (Kingston Harbour), Cuba, Florida, Bermuda, Woods Hole, off Nova Scotia E, off Newfoundland, S Iceland, Ireland, Lough Hyne, Scotland (Loch Thurnaig), English Channel, Strait of Dover (near the coast south of Boulogne), North Sea, Oslo fjord, S & SW Barents Sea,NW Spain, Ibero-moroccan Bay, off W Tangier, Medit. (M'Diq, Alboran Sea, Habibas Is., Sidi Fredj coast, Gulf of Annaba, El Kala shelf, laguna Mar Menor, Castellon, Baleares, Banyuls, Thau lagoon, Berre lagoon, Marseille, Toulon Harbour, Villefranche-s-Mer, Genoa Harbour, Tyrrhenian Sea, Naples, Gulf of Taranto, Taranto Harbour, Tunisia (Sfax, G. of Gabès), Ionian Sea, Adriatic Sea, Vlora Bay, Mljet Is., Venice, Po delta, Sibenik harbour, Port of Koper; Aegean Sea, Thracian Sea, Marmara Sea, Black Sea, Sebastopol, Black River estuary, Iskenderun Bay, Lebanon Basin, W Egyptian coast, Alexandria, Bardawill Lagoon), Suez Canal, Lake Timsah, Taba, Hurghada, Safâga, , off Sharm El-Sheikh, Red Sea, Gulf of Oman, Arabian Sea, Arabian Gulf, UAE coast, Kuwait, Sri Lanka, Madagascar (Nosy Bé), Rodrigues Is., Indian, India (Lawson's Bay, Gulf of Mannar, Palk Bay, Chilka Lake), Kurau River, Christmas Is., Nicobar Is., Perai River estuary (Penang), Straits of Malacca, G. of Thailand, Indonesia-Malaysia, Viet-Nam, China Seas (Bohai Sea, Yellow Sea, East China Sea, South China Sea, Xiamen Harbour, N Yangtze Riv. estuary), Taiwan (Kaohsiung Harbor, Pitou, Kuroshio Current), Korea (E,S & W), Japan Sea, Japan ( Konshu: Maizuru Bay, Kuchinoerabu Is., Seto Inland Sea, Ariake-kai, Yatsushiro-kai, Tokyo Bay, Tanabe Bay, Kyushu: Shijiki Bay), Kuril Is., Palau Is., Pacif. (tropical), Hawaii, Kaneohe Bay, California (San Pedro, San Diego), Baja California (Bahia Magdalena, La Paz), G. of California, Bahia de los Angeles, Coyuca lagoon (Guerrero), Zihuatanejo Bay, Acapulco Bay, W Mexico, Is. Fidji, Is. Samoa, Australia (North West Cape, New South Wales), New Caledonia, New Zealand, Moorea Is., SE Pacif., Galapagos-Ecuador, Peru, off Chile, Chile (N, off Santiago) | | | | N: | 368 | | | | Lg.: | | | (11) F: 0,56; (45) F: 0,65-0,5; M: 0,57-0,48; (46) F: 0,53-0,5; M: 0,5-0,48; (66) F: 0,64-0,51; M: 0,55; (91) F: ± 0,62; M: ± 0,54; (109) F: 0,65-0,5; M: 0,55-0,45; (142) F: 0,8-0,7; M: 0,6-0,5; (155) F: 0,62-0,54; M: 0,51-0,48; (179) F: 0,66-0,6; M: 0,55-0,45; (237) F: 0,55-0,6; M: 0,40; (327) F: 0,68-0,55; M: 0,57-0,56; (373) F: 0,73-0,49; M: 0,61-0,48; (432) F: 0,95-0,55; (434) F: 0,65-0,42; M: 0,49-0,45; (449) F: 0,53-0,5; M: 0,5-0,48; (579) M: 0,63-0,54; (618) F: 0,63-0,42; M: 0,48-0,45; (624) F: 0,72-0,58; M: 0,53-0,47; (627) F: 0,59-0,49; 0,53-0,49; M: 0,48-0,47; (649) F: 0,53; M: 0,44; (651) F: 0,67-0,57; (786) F: 0,62; (880) F: 0,50-0,70; M: 0,44-0,60; F: 0,44-0,60; (991) F: 0,5-0,8; M: 0,48-0,6; (1085) F: 0,48-0,6; M: 0,45-0,5; (1185) F: 0,68-0,65; M: 0,65-0,62; (1302) F: 0,310-0,335; M: 0,301-0,336; {F: 0,31-0,95; M: 0,30-0,63} | | | | Rem.: | Euryhaline, epipelagic, neritic, estuaries, coastal, harbors.
For Gurney (1927, p.159), there are two distinct forms of this species in the Suez Canal's collection. All agree precisely in the structure of the mandible, maxilla, and in the arrangement of spines on the legs with the type, but there are slight differences:
1 - Typical O. nana
Length: 0.53-0.55 mm. Thorax slightly longer than abdomen (including last thoracic segment and furca). Furca equal to anal somite, twice as long as wide. A1 reaching at most the middle of 3rd free thoracic somite.
2 - Southern form:
- Length: 0.62-0,69 mm. Abdomen longer than thorax. Furca equal to or longer than anal somite, about 3 times as long as broad. A1 reaching usually slightly beyond 3rd free thoracic somite..
Oithona nana Form plumosa Nishida, 1985 (F)
Ref.: Nishida, 1985 a (p.55, Descr.F,M, figs.F)
Ref. compl.: Razai & al., 2004 (p.490, tab.2, Rem.)
Loc.: Arabian Gulf, Sri-Lanka, Straits of Malacca, G. of Thailand, New-Caledonia.
See in Oithona robertsoni Table 2 from McKinnon (2000, p.108).
For Mihneva & Stefanova (2013, p.119) the species is an eurythermic copepod that was widely distributed in the Black Sea in the 1970's and 1980's, where it was present ay high densities in the surface layer. Since the beginning of the 1990's, O. nana has totally disappeared from the zooplankton community.
After Gubanova & al. (2013, p.5) one of the results of the destructive invasion of the predatory ctenophore Mnemiopsis leydyi (Agassiz, 1895) in the Black Sea in the 1990s was the complete disappearance of O. nana.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 6 corresponding to small ambush feeding omnivorous.
For Atienza & al. (2006) (0.33 µg C ind-1 respiration rates are 0.18-0.26 µg C day-1 at 25.5-27°C.
Cf. Remark concerning the occurrence of this species in the waters of Japan and the species Oithona davisae.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 19, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
