|
|
 |
|
Cyclopoida ( Order ) |
|
|
|
Oithonidae ( Family ) |
|
|
|
Oithona ( Genus ) |
|
|
| |
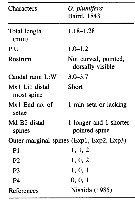
Oithona plumifera Baird, 1843 (F,M) | |
| | | | | | | Syn.: | no Oithona plumifera var. atlantica : Früchtl, 1923 a (p.141, figs.F, Rem.);
no O. plumifera : Mori, 1937 (1964) (p.109, figs.F,M); Kos, 1972 (figs.B, X).Oithona plumiphera : Fernandez de Puelles & al., 1996 (p.97, occurrence: p.101, lapsus calami) | | | | Ref.: | | | Giesbrecht, 1892 (p.537, 548, 774, figs.F,M); Wheeler, 1901 (p.186, fig.F, Rem. F,M); Thompson & Scott, 1903 (p.236, 255); Esterly, 1905 (p.207, figs.F,M); Farran, 1908 b (p.89, Rem.); A. Scott, 1909 (p.194, Rem.); Wolfenden, 1911 (p.362); Pesta, 1912 a (p.56); 1913 (p.33); Burckhardt, 1913 (p.429); Lysholm, 1913 (p.7); Farran, 1913 a (p.183); Rosendorn, 1917 a (p.10, figs.F,M); Pesta, 1920 (p.553); Früchtl, 1924 b (p.64, figs.F); Gurney, 1927 (p.158, Rem.); Kiefer, 1929 g (p.4, Rem.F,M, fig.F); Farran, 1929 (p.210, 282); Rose, 1929 (p.55); Wilson, 1932 a (p.311, figs.F,M); Dakin & Colefax, 1933 (p.208); Rose, 1933 a (p.282, figs.F,M); Farran, 1936 a (p.123, Rem.); Dakin & Colefax, 1940 (p.115, figs.F); Lysholm & al., 1945 (p.42, Rem.); Sewell, 1947 (p.255, Rem.F, Rem.); Davis, 1949 (p.74, fig.F, Rem.F,M); Sewell, 1951 (p.371, fig.F, Rem.: parasites); Krishnaswamy, 1953 (p.64, figs.F); Lindberg, 1955 a (p.465: Rem.); Chiba & al., 1957 (p.310); 1957 a (p.12); Crisafi, 1958 (p.179, figs.M, juv.); 1959 c (p.70, figs.F,M); Tanaka, 1960 (p.61, figs.F); Fish, 1962 (p.23); Fagetti, 1962 (p.38); Kasturirangan, 1963 (p.74, 77, figs.F); Tanaka, 1964 (p.13); Vilela, 1965 (p.12); Saraswathy, 1966 (1967) (p.89); Owre & Foyo, 1967 (p.107, figs.F,M); Vilela, 1968 (p.29); Ramirez, 1969 (Rem: p.85); Wellershaus, 1970 (p.483, 484); Corral Estrada, 1970 (p.210, Rem.); Minoda, 1971 (p.44); Shih & al., 1971 (p.55, 156); Razouls, 1972 (p.95, Annexe: p.107, figs.F); Björnberg, 1972 (p.88, fig., Rem.N); Shuvalov, 1972 (1975) (p.180, figs.F, Rem.); Marques, 1973 (p.246); Chen & al., 1974 (p.36, figs.F,M); Marques, 1975 (p.50); 1976 (p.1000); Nishida & al., 1977 (p.140, figs.F,M, Rem.); Séret, 1979 (p.158, figs.F); Dawson & Knatz, 1980 (p.9, figs.F,M); Shuvalov, 1980 (p.91, figs.F, Rem.M); Ferrari & Bowman, 1980 (p.17, figs.F,M, Rem.); Björnberg & al., 1981 (p.663, figs.F); Marques, 1982 (p.769); Nishida, 1985 a (p.68, Descr.F, figs.F, Rem., p.123); Sazhina, 1985 (p.88, figs.N); Nishida, 1986 (p.386); Lin & Nakamura, 1993 (p.449); Yoo & Lim, 1993 (p.91, 95, 97, Table 1, fig.2); Kim & al., 1993 (p.271); Chihara & Murano, 1997 (p.938, Pl.195,200: F,M); Bradford-Grieve & al., 1999 (p.886, 966, figs.F,M); Al-Yamani & Prusova, 2003 (p.105, figs.F); Conway & al., 2003 (p.203, figs.F,M, Rem.); Avancini & al., 2006 (p.124, Pl. 92, figs.F, Rem.); Vives & Shmeleva, 2010 (p.67, figs.F,M, Rem.); Al-Yamani & al., 2011 (p.96, 100, figs.F,M); Cornils & Wend-Heckmann, 2015 (p.243, Table 1, Fig.2, molecular biology) | 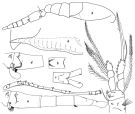 issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.18, Fig.10]. Female (from Caribbean area): a, urosome (lateral left side); b, urosomal segment 4 (ventral); c, urosomal segment 5 and caudal rami (dorsal); d, urosomal segments 1 and 2 with attached sprmatophore) (lateral left side); e, A1; f, P4. Nota: Prosome/urosome = 1.1. A1 with tiny hairs on 1-10 free segments. Urosomal segment 2 with tuft of hairs on anteroventral surface and small row of spinules posterior to knob near genital opening; knob with small, thin spine and spinule ventral to it. Urosomal segment 4 with 2 rows of 4 thick hairs on anteroventral margin. Urosomal segment 5 with lateral and and dorsal rows of hairs. Caudal rami with hairs on posteromedial edge. Endopodal segment 2 of P4 both setae modified, thick, curved with flange over distal 1/3. Endopodal segment 3 of P4 with proximal seta modified, thick, slightly curved toward tip, with flange on distal 1/3 Male: g, habitus (lateral right side); h, cephalosome paortion and flap (lateral right side). Nota: Prosome/Urosome = 1.5. Cephalosome flap digitiform, attenuate end reaching beyond posterior edge of pediger segment 1. \\\\\\\\\\\\\\\"Pore signature\\\\\\\\\\\\\\\" similar to O. hebes. Exopodal segments of P1-4 of this species agree with Nishida & al. (1977), not with Wellershaus (1970) . Giesbrecht's descriptions and illustrations show: 1-1-2, 1-1-3, 1-1-3, 1-1-2 external setae.
|
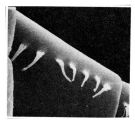 issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.24, Fig.15 f]. Female (SEM): f, hairs on A1; without associated pores, on 1-10 free segments.
|
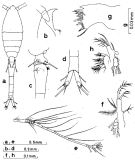 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.69, Fig.37]. Female: a, habitus (dorsal); b, forehead (lateral); c, last thoracic segments and genital segment (lateral right side); d, anal segment and caudal rami (ventral); e, A1; f, Md (mandibular palp); g, Md (biting edge); h, Mx1. Nota: Proportional lengths of urosome segments and caudal ramus 10:28:15:16:16:15.
|
 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.70, Fig.38]. Female: a, A2; b, Mx2; c, Mxp; d, P1; e, P2; f, P3; g, P4.
|
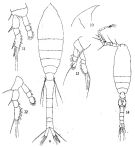 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.4, Figs.9-14]. Female (from China Seas): 9, habitus (dorsal); 10, forehead (lateral); 11, P2; 12, P3; 13, P4. Male: 14, habitus (dorsal).
|
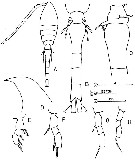 issued from : F.Y. Al-Yamani & I. Prusova in Common Copepods Northwestern Arabian Gulf : Identification Guide. Kuwait Institute for Scientific Research, 2003. [p.106, Fig.39]. Female: A, habitus (dorsal); B, urosome (dorsal); C, genital segment (lateral left side); D, forehead (lateral); E, exopod of P1; F, exopod of P2; G, exopod of P3; H, exopod of P4. Nota: Exopod of P1-P4 with 1.1.2, 1.0.2, 1.0.1, 0.0.1 outer marginal spines.
|
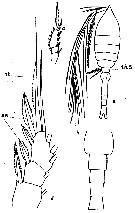 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.208, Fig.50]. Female (from San Diego Region): a, habitus (dorsal; th5 = thoracic segment 5); d, exopodite of P1 (se = outer marginal bristles; st = terminal bristle). Male: b, abdomen; c, exopodite of P3.
|
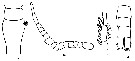 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped."Valdiviella", 1917, 23. [p.10, Fig.1]. Female: a, genital segment (lateral). Nota: Proportion of lengths (p.cent) Prosome : 53.7, Urosome : 46.2 . Relative lengths of urosomal segments and caudal rami 8 : 20 : 10 : 10 : 11 : 9. Setal formula of the exopod swimming legs P1 to P4 (Se = outer sete ; Si = inner setae), P1 : 1, 1, 2 Se ; 1, 1, 4 Si ; P2 : 1, 0, 2 Se ; 0, 1, 5 Si ; P3 : 1, 0, 1 Se ; 0, 1, 5 Si ; P4 : 0, 0, 1 Se ; 0, 1, 5 Si . Male: b, left A1; c, exopod of P2; d, posterior part of urosme. Nota: Proportion of total lengths (p.cent) Prosome : 59.15, Urosome : 40.85 . Relative lengths of urosomal segments and caudal rami : 4 : 13 : 9 : 8 : 5 : 6 : 7. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae), P1 : 1, 1, 3 Se ; P2 : 1, 0, 2 Se ; P3 : 1, 0, 1 Se ; P4 : 0, 0, 1 Se.
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.93, Fig.14]. Female: 1-2, habitus (dorsal); 3, urosome (dorsal); 4, forehead (lateral); 5, A2; 6, Mx2; 7, Mxp; 8, P1; 9, P2; 10, P3; 11, P4; 12, thoracic segments 4 and 5, genital segment (dorsal); 13, genital segment (lateral); 14, anal segment and caudal rami (dorsal). Nota after Vives & Shmeleva (2010, p.67): Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 2; 1, 1, 4. P2: 1, 0, 2; 1, 1, 5 P3: 1, 0, 1; 0, 1, 5. P4: 0, 0, 1; 0, 1, 5.
|
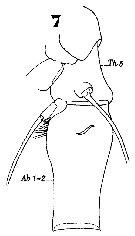 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.44, Fig.7]. Female: 7, thoracic segment 5 and abdominal segments 1-2 (lateral).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.44, Fig.1]. Male: 1, habitus (dorsal).
|
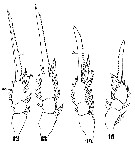 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.44, Figs.12, 13, 14, 15]. Male: 12, P4; 13, P3; 14, P2; 15, P1. After Vives & Shmeleva (2010, p.68): Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 2; 1, 1, 4. P2: 1, 1, 3; 1, 1, 5 P3: 1, 1, 3; 1, 1, 5. P4: 1, 1, 2; 1, 1, 5.
|
 issued from : Y. Al-Yamani, V. Skryabin, A. Gubanova, S. khvorov & I. Prusova in Marine Zooplankton Practical Guide for the Northwestern Arabian Gulf, 2, 2011. [p.100, Fig.243]. Female (from Kuwait): a, habitus with an eggs sac on the right side (dorsal); b, habitus (lateral).
|
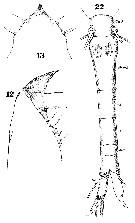 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.12, 13, 22]. Female: 12-13, forehead (lateral and dorsal, respectively); 22, urosome (dorsal).
|
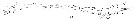 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Fig.33]. Female: 33, A1 (ventral surface).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Fig.28]. Female: 28, A2.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Fig.32]. Female: 32, Md.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Fig.25]. Female: 25, Mx1.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.27, 29]. Female: 27, Mxp; 29, Mx2 (posterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.44-47]. Female: 44, exopodite of P2; 45, exopodite of P3; 46, P4; 47, P1 (anterior view)
|
 issued from : C. Razouls in Th. Doc. Etat Fac. Sc. Paris VI, 1972, Annexe. [Fig.63]. Female (from Banyuls, G. of Lion): A, P1; B, P4; C, P3; D, P2; E, forehead (lateral); F, habitus (dorsal). ST = terminal seta of exopod. Nota: Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 2; 1, 1, 4. P2: 1, 0, 2; 0, 1, 5 P3: 1, 0, 1; 0, 1, 5. P4: 0, 0, 1; 0, 1, 5. The inner seta on the exopodite segment 1 of P2 is absent compared with the drawings from Shuvalov (1980) and perhaps Nishida (1985).
|
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.141, Fig.14]. Female (from Suruga Bay and adjacent waters): a, habitus (dorsal), b, forehead (lateral); c, 5th thoracic segment with P5, and genital segment (lateral); d, Md; e, Mx1; f, P1; g, P2; h, P3; i, P4. Nota: Setae formula of exopod P1 to P4 (outer setae in first, inner setae in second). P1: 1,1,2; 1,1,4. P2: 1,0,2; 0,1,5. P3: 1,0,1; 0,1,5. P4: 0,0,1; 0,1,5. Prosome length/urosome length = 1.02-1.21.
|
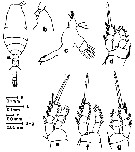 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.142, Fig.15]. Male (from of Sagami Bay): a, habitus (dorsal), b, forehead (lateral); c, Md; d, P1; e, P2. Nota: Setae formula of exopod P1 to P4 (outer setae in first, inner setae in second). P1: 1,1,2; 1,1,4. P2: 1,1,3; 1,1,5. P3: 1,1,3; 1,1,5. P4: 1,1,2; 1,1,5. Prosome length/urosome length = 1.78.
|
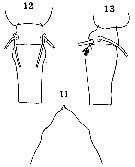 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [Pl. XXVI, 11-13]. Female (from 18°54'S, 60°39'E): 11, forehead (dorsal); 12-13, 5th thoracic segment and genital segment (dorsal and lateral, respectively). Figs. 11, 12, 13 at the same scale. Nota: Prosome and urosome in the proportional lengths 51 tp 49. Genital segment with a patch of hairs on the proximal ventral surface.
|
 Issued from K.-I. & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2). [Key of female]: ]Morphological characters of Oithona plumifera female in Korean waters :
1 - Anterior part of prosome produced into pointed rostrum in dorsal view.
2 - Exopod 3rd segment of P1 with 2 outer marginal spines. 3 - Number of setae in exopod of Md more than 3, and prosome length than 3.0 times of prosome width. 4 - Outer marginal spine in exopod 3rd segment of P4 curved smoothly. 5 - Endopod of Md with 3 setae, genital segment ventrally with tuft of hairs.
|
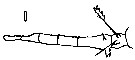
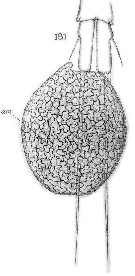 Issued from : E. Chatton in Arch. Zool. Exp. & Gen., 1920, 59, [Pl. XVI, Fig.181]. Oithona plumifera (from Banyuls) showing sporifere sac joined at the caudal rami, corresponding to the gonosphere of Paradinium poucheti (filoplasmod presporal stage).
|
 Issued from : A. Cornils & B. Wend-Heckmann in Helgol. Mar. Res., 2015, 69. [p.246, Table 1]. Oithona plumifera charcteristics in the semi-enclosed bight in the northern Germann Wadden Sea (North Sea) Compare with O. nana, O. similis, O. atlantica and O. davisae in the North Sea.
|
 Issued from : K.-I. Yoo & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2) [p.99, Fig.2, I] Female (from Korean Waters): I, abdomen (lateral view).
| | | | | Compl. Ref.: | | | Mrazek, 1902 (p.517, 524); Cleve, 1904 a (p.193); Damas & Koefoed, 1907 (p.395, tab.II); Chatton, 1920 (p.17, parasited); Rose, 1925 (p.152); Wilson, 1932 (p.49); 1942 a (p.196); Massuti Alzamora, 1942 (p.101, Rem.); Oliveira, 1945 (p.191); Sewell, 1948 (p.397, 461, 514); C.B. Wilson, 1950 (p.270); Yamazi, 1958 (p.153, Rem.); Minoda, 1958 (p.253, Table 1, 2, abundance); Deevey, 1960 (p.5, Table II, annual abundance) ; Zoppi, 1961 (p.219);; Ganapati & Shanthakumari, 1962 (p.10, 16); Grice & Hart, 1962 (p.287, table 3, 4: abundance); Gaudy, 1962 (p.93, 99, Rem.: p.117) ; Giron-Reguer, 1963 (p.56); V.N. Greze, 1963 a (tabl.2); Shmeleva, 1963 (p.141); Duran, 1963 (p.25); Gaudy, 1963 (p.30, Rem.); Björnberg, 1963 (p.74, Rem.); De Decker, 1964 (p.16, 23, 30); De Decker & Mombeck, 1964 (p.13); Shmeleva, 1965 b (p.1350, lengths-volume-weight relation); Pavlova, 1966 (p.44); Furuhashi, 1966 a (p.295, vertical distribution vs mixing Oyashio/Kuroshio region, Table 9); Neto & Paiva, 1966 (p.29, Table III, annual cycle); Mazza, 1966 (p.73); 1967 (p.367); Ehrhardt, 1967 (p.742, geographic distribution, Rem.); Séguin, 1968 (p.488); Evans, 1968 (p.14); Delalo, 1968 (p.138); Champalbert, 1969 a (p.641); Dowidar & El-Maghraby, 1970 (p.269); Deevey, 1971 (p.224); Gamulin, 1971 (p.381, tab.2); Salah, 1971 (p.320); Paulmier, 1971 (p.168); Binet & al., 1972 (p.71); Apostolopoulou, 1972 (p.329, 373, fig.9); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %, Table IV); Subbaraju & Krishnamurphy, 1972 (p.25, 26 ); Binet, 1973 (p.77); Björnberg, 1973 (p.357, 388); Chatton & Soyer, 1973 (p.27, Rem.: p.33, parasite); Desgouille, 1973 (p.1, 131, Rem.: p.136); Guglielmo, 1973 (p.399); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Palet, 1975 (p.660); Vives & al., 1975 (p.53, tab.II, III, IV); Falconetti & Seguin, 1977 (p.188); Zalkina, 1977 (p.339, tab.1); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Ferrari, 1977 (p.410); Boxshall, 1977 b (p.552); Deevey & Brooks, 1977 (p.156, tab.2, Station "S"); Klekowski & al., 1977 (p.467, respiration); Rudyakov, 1982 (p.208, Table 2); Dessier, 1979 (p.201, 207); Vaissière & Séguin, 1980 (p.23, tab.2); Castel & Courties, 1982 (p.417, Table II, spatial distribution); Vives, 1982 (p.295); Kovalev & Shmeleva, 1982 (p.85); Dessier, 1983 (p.89, Tableau 1, Rem., %); Tremblay & Anderson, 1984 (p.7, Rem.); Bityukov & al., 1984 (tab.1); Guangshan & Honglin, 1984 (p.118, tab.); Scotto di Carlo & al., 1984 (p.1041); Dussart, 1984 (p.25, 64: occurrence): Binet, 1984 (tab.3); 1985 (p.85, tab.3); Kimmerer & McKinnon, 1985 (p.150); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea); Regner, 1985 (p.11, Rem.: p.40); Jansa, 1985 (p.108, Tabl.I, II, III, IV, V); Brinton & al., 1986 (p.228, Table 1); M. Lefèvre, 1986 (p.33); Turner, 1986 (p.289, fig. 8: pellet contents): Diouf & Diallo, 1987 (p.260); Lozano Soldevilla & al., 1988 (p.61); Williams D. & al., 1988 (p.580); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Pancucci-Papadopoulou & al., 1990 (p.199); Othman & al., 1990 (p.561, 564, Table 1); Hirakawa & al., 1990 (tab.3); Fransz & al., 1991 (p.8); Lindo, 1991 (tab.3); Yoo, 1991 (tab.1); Hirakawa, 1991 (p.376: fig.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Seguin & al., 1993 (p.23); Paffenhöfer, 1993 (p.37, egg production); Lopes, 1994 (tab.1); Kouwenberg, 1994 (tab.1); Palomares Garcia & Vera, 1995 (tab.1); Hirakawa & al., 1995 (tab.2); Shih & Young, 1995 (p.75); Roff & al., 1995 (p.165, Table 3: nauplii, copepodites growth rates); Webber & Roff, 1995 a (p.481, table 1, 2, 3, 4, 5, 6, 7, 8, biomass, production); ); Krause & al., 1995 (p.81, Rem.: p.134); Hajderi, 1995 (p.542); Webber & al., 1996 (tab.1); Kotani & al., 1996 (tab.2); Go & al., 1997 (tab.1); Park & Choi, 1997 (Appendix); Suarez-Morales & Gasca, 1997 (p.1525); Hopcroft & Roff, 1996 (p.789, diel egg production); Sharaf & Al-Ghais, 1997 (tab.1); Hure & Krsinic, 1998 (p.77, 103); Gilabert & Moreno, 1998 (tab.1, 2); Hopcroft & al., 1998 (tab.2); Noda & al., 1998 (p.55, Table 3, occurrence); Alvarez-Cadena & al., 1998 (t.1,2,3,4); Hsieh & Chiu, 1998 (tab.2); Suarez-Morales & Gasca, 1998 a (p.112); Siokou-Frangou, 1999 (p.478); Hernandez-Trujillo, 1999 (p.284, tab.1); Lopes & al., 1999 (p.215, tab.1); El-Serehy, 1999 (p.172, Table 1, occurrence); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Siokou-Frangou & al., 1999 (p.205, Table 5); Bragina, 1999 (p.196); Razouls & al., 2000 (p.343, Appendix); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Ueda & al., 2000 (tab.1); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Suarez-Morales & Gasca, 2000 (1247, tab.1); Suarez-Morales & al., 2000 (p.751, tab.1); Seridji & Hafferssas, 2000 (tab.1); Lopez-Salgado & al., 2000 (tab.1); Plounevez & Champalbert, 2000 (p.175, Table III, IV, V, abundance vs fish); Moraitou-Apostolopoulou & al., 2000 (tab.I, fig.7); Holmes & Gotto, 2000 (p.3, Rem.); Alvarez-Silva & Gomez-Aguirre, 2000 (p.163: tab.2); Sautour & al., 2000 (p.531, Table II, abundance); d'Elbée, 2001 (tabl.1); Dalal & Goswami, 2001 (p.22, fig.2); Fransz & Gonzalez, 2001 (p.255, tab.1); Hidalgo & Escribano, 2001 (p.159, tab.2); El-Serehy & al., 2001 (p.116, Table 1: abundance vs transect in Suez Canal); Bressan & Moro, 2002 (tab.2); Zerouali & Melhaoui, 2002 (p.91, Tableau I); Hernandez-Trujillo & Suarez-Morales, 2002 (p.748, tab.1); Dunbar & Webber, 2003 (tab.1); Vukanic, 2003 (p.139, tab.1); Fernandez de Puelles & al., 2003 (p.123, fig.5); McKinnon & al., 2003 (p.101, tab.2, fig.11); Vieira & al., 2003 (p.S163, Table 2, abundance); Hsieh & al., 2004 (p.398, tab.1, p.399, tab.2); Rezai & al., 2004 (p.486, tab.2, 3, abundance, Rem., p.495, tab.8); Chang & Fang, 2004 (p.456, tab.1); Daly Yahia & al., 2004 (p.366, fig.4, tab.1); Lan & al., 2004 (p.332, tab.1, tab.2); Fernandez de Puelles & al., 2004 (p.654, fig.7); Satapoomin & al., 2004 (p.109, tab.6); Lo & al., 2004 (p.89, tab.1); Vukanic & Vukanic, 2004 (p.9, tab. 2, 3); Vukanic & Vukanic, 2004 (p.361, tab.2, fig.2); Kazmi, 2004 (p.231, Rem.: p.232); Mazzocchi & al., 2005 (p.155); Lakkis & al., 2005 (p.152); Camisotti & al., 2005 (p.99); Alvarez-Silva & al., 2005 (p.39); Uriarte Villate, 2005 (p.863, tab.I); Wiggert & al., 2005 (p.1013: feeding behavior); Berasategui & al., 2005 (p.485, tab.1); Obuid Allah & al., 2005 (p.123, occurrence % vs metal contamination); Prusova & Smith, 2005 (p.76, 78); Baker & al., 2005 (tabl.); Berasategui & al., 2006 (p.485: fig.2); Zuo & al., 2006 (p.164: tab.1); Isari & al., 2006 (p.241, tab.II); Marques & al., 2006 (p.297, tab.III); Hwang & al., 2006 (p.943, tabl. I); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Dias & Araujo, 2006 (p.70, Rem., chart); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño); Lavaniegos & Jiménez-Pérez, 2006 (p.149, tab.2, Rem.); Mageed, 2006 (p.168, Table 4); Zervoudaki & al., 2006 (p.149, Table I); Hwang & al., 2007 (p.25); Fernandez de Puelles & al., 2007 (p.338, fig.7); Albaina & Irigoien, 2007 (p.435: Tab.1); Valdés & al., 2007 (p.104: tab.1); Busatto, 2007 (p.26, Tab.2); Porri & al., 2007 (p.711); Khelifi-Touhami & al., 2007 (p.327, Table 1); Marques S.C. & al., 2007 (p.725, Table 1, fig.4, climate variability); McKinnon & al., 2008 (p.843: Tab.1, fig.7); Lopez Ibarra, 2008 (p.1, Table 1, 2, figs.11, 14: abundance); Cabal & al., 2008 (289, Table 1); Humphrey, 2008 (p.84: Appendix A); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.514, 522); Rakhesh & al., 2008 (p.154, abundance vs stations); Fernandes, 2008 (p.465, Tabl.2); Muelbert & al., 2008 (p.1662, Table 1); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Rossi, 2008 (p.90: Tableau XII); Lan Y.-C. & al., 2008 (p.61, Table 1, % vs stations); Ohtsuka & al., 2008 (p.115, Table 4, Rem.: in Kawasaki Harbor); Chiba & al., 2009 (p.1846, Table 1, occurrence vs temperature change); Pagano, 2009 (p.116); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Primo & al., 2009 (p.341, Table1, interannual variations); Licandro & Icardi, 2009 (p.17, Table 4); Lan & al., 2009 (p.1, Table 2); C.E. Morales & al., 2010 (p.158, Table 1); Brugnano & al., 2010 (p.312, Table 2, 3); Lidvanov & al., 2010 (p.356, Table 3); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Hidalgo & al., 2010 (p.2089, Table 2); Dias & al., 2010 (p.230, Table 1); Mazzocchi & Di Capua, 2010 (p.428); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hafferssas & al., 2010 (p.1281, Table III, abundance vs spatial distribution); Fazeli & al., 2010 (p.153, Table 1); W.-B. Chang & al., 2010 (p.735, Table 2, 4, fig.5, abundance); Hsiao S.H. & al., 2011 (p.475, Appendix I); Salah S. & al., 2011 (Tableau 1); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Magris & al., 2011 (p.260, abundance, interannual variability); Moscatello & al., 2011 (p.80, Table 4); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); S.C. Marques & al., 2011 (p.59, Table 1); Chen H; & al., 2011 (p.84, spatial & temporal variations); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Hsiao & al., 2011 (p.317, Table 2, indicator of seasonal change); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Mazzocchi & al., 2011 (p.1163, Table II, fig.6, long-term time-series 1984-2006); Beltrao & al., 2011 (p.47, Table 1, density vs time); Mazzocchi & al., 2012 (p.135, annual abundance 1984-2006); Pagano & al., 2012 (p.538, Table 1); Delpy & al., 2012 (p.1921, Table 2); Glushko & Lidvanov, 2012 (p.138, Tableau 1, 3); Uysal & Shmeleva, 2012 (p.909, Table I); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Gonçalves & al., 2012 a (p.16, abundance, diel vertical behavior); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Aubry & al., 2012 (p.125, table 3, interannual variation); Naz & al., 2012 (p.61, Table 4, relative abundance); Dorgham & al., 2012 (p.473, Table 3: abundance %); Gonçalves & al., 2012 (p.321, abundance vs climatic variability); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Lavaniegos & al., 2012 (p. 11, Appendix); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Bode & al., 2012 (p.108, spatial distribution vs time-series, % biomass); Johan & al., 2012 (2013) (p.1, Table 1); Zamora-Terol & Saiz, 2013 (p.376, Rem.: Table 3, egg production); Gubanova & al., 2013 (in press, p.4, Table 2); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Palomares-Garcia & al., 2013 (p.1009, Table I, V, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; as Oithona plumifera plumifera; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Melo Jùnior & al., 2013 (p.363, Table 2, egg production); Rakhesh & al., 2013 (p.7, Table 1, abundance vs stations); Jagadeesan & al., 2013 (p.27, Table 3, seasonal variation); Sobrinho-Gonçalves & al., 2013 (p.713, Table 2, fig.8, seasonal abundance vs environmental conditions); Mendoza Portillo, 2013 (p.37: Fig.7, seasonal dominance); Lidvanov & al., 2013 (p.290, Table 2, % composition); Zamora Terol, 2013 (p.174: table 5); Terbiyik Kurt & Polat, 2013 (p.1163, Table 2, seasonal distribution); Eisner & al., 2013 (p.87, Table 3: abundance vs. water structure); Fernandez de Puelles & al., 2014 (p.82, Table 3, seasonal abundance); Bonecker & al., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Lopez-Ibarra & al., 2014 (p.453, fig.6, Table 2, biogeographical affinity); Pansera & al., 2014 (p.221, Table 2, abundance); Mazzocchi & al., 2014 (p.64, Table 3: occurrence); Ortega & al., 2014 (p.495, Table I, %); Fierro Gonzalvez, 2014 (p.1, Tab. 3); Zaafa & al., 2014 (p.67, Table I, occurrence); Aboul Ezz & al., 2014 (p.283, Rem.: p.284); Zakaria, 2014 (p.3, Table 1, abundance vs. 1960-2000); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance, Table 3: diversity index); Araujo & al., 2016 (p.1, Table 3, abundance, %); Zakaria & al., 2016 (p.1, Table 1, 3, 4); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); Ortega & al., 2017 (p.123, fig. 1, 6, table 2, abundance, %) ; Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Abo-Taleb & Gharib, 2018 (p.139, Table 5, p.149: occurrence %); Dias & al., 2018 (p.1, Tables 2, 5: vertical distribution, abundance vs. season); Hure M. & al., 2018 (p.1, Table 1: abundance, % composition, Table 2: correlations); Palomares-Garcia & al., 2018 (p.178, fig.3: relative frequency, Table 1); Acha & al., 2020 (p.1, Table 3: occurrence % vs. ecoregions). | | | | NZ: | 22 + 1 doubtful | | |
|
Distribution map of Oithona plumifera by geographical zones
|
| | | | | | | | | | | | | | | | | |  Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.123, Fig.69]. Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.123, Fig.69].
Indo-Pacific geographical distribution of Oithona plumifera. Dotted line: AC, Arctic Convergence; SC, Subtropical Convergence. |
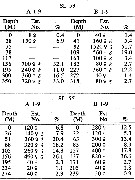
 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. "Valdiviella", 1917, 23. [Taf. I]. issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. "Valdiviella", 1917, 23. [Taf. I].
The genus Oithona: Distribution of species sampled during the Deutsche Tiefsee Expedition 1898-99.
The size of symbols indicates the relative quantitty of the species. |
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.108, Table 20]. issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.108, Table 20].
Vertical distribution of Oithona plumifera collected at the ''40-mile station'' in the Florida Current.
SL 53: 18 V 1958; SL 55: 21 VII 1958.
A: midday; B: midnight. |
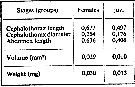 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 38]. Oithona plumifera (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 38]. Oithona plumifera (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
 Issued from : C. Séret in Thesis 3ème Cycle, UPMC, Paris 6. 1979, Annexe. [p.52]. Issued from : C. Séret in Thesis 3ème Cycle, UPMC, Paris 6. 1979, Annexe. [p.52].
Geographical occurrences of Oithona plumifera in the Indian Ocean and Antactic zone. [after publications from: Brady, 1883, 1918; Thompson, 1900; Wolfenden, 1908, 1911; With , 1915; Rosendorn, 1917; Farran, 1929; Sewell, 1929, 1947; Brady & Gunther, 1935; Steuer, 1929, 1392, 1933; Ommaney, 1936; Vervoort, 1957; Tanaka, 1960; Brodsky, 1964; Seno, 1966; Andrews, 1966; Grice & Hulsemann, 1967; Seno, 1966; Frost & Fleminger, 1968; Voronina, 1970; Zverva, 1972].
Nota: C. Séret notes the occurrence at stations 50.5°S, 71°E; 51°S, 65°E and 56°S, 70°E in which the individuals were very numerous. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.484, Table 1].
Length-weight regression of Oithona plumifera in oceanic waters of Discovery Bay (Jamaica) in 60- and 200 m vertical hauls, using 200- and 64-µm mesh aperture of plankton nets, from June 1989 to July 1991 (monthly samples). |
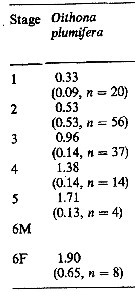 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 2].
Mean biomass (µg) of copepodite stages fot Oithona plumifera from Discovery Bay (Jamaica). SD: standard deviation and n: number of individuals measured. |
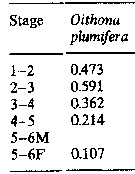 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.485, Table 3].
Stage weight increments (In mmax mmin-1) for Oithona plumifera from Discovery Bay (Jamaica), calculed from mean biomass of each stage. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 4]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 4].
Stage duration (days) for copepodite stages of Oithona plumifera |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 5]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.487, Table 5].
Mean number of eggs per female for Oithona plumifera from Discovery Bay (Jamaica), and corrected egg development time in days. n: nuber of observations. |
 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 6]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 6].
Estimates of generation times (in days) from egg to first egg clutch for Oithona plumifera from Discovery Bay (Jamaica) based on minimum and maximum development time of individual life history stages. |
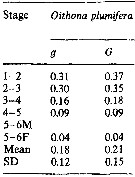 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 7]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 7].
Growth coefficient (g) and daily specific growth rate (G) for Oithona plumifera from Discovery Bay (Jamaica).
mean and range of daily specific growth rates (G, day-1) for nauplii and copepodites (all stages, excluding moult from copepodite stage 5 to adult. (Data from Chislom & Roff, 1990; Webber, 1993; Hopcroft & Roff, unpubl.). |
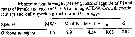 Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 8]. Issued from : M.K. Webber & J.C. Roff in Mar. Biol., 1995, 123. [p.488, Table 8].
Oithona plumifera from Discovery Bay (Jamaica).
AFDH = ash-free dry weight. |
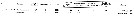 Issued from : S. Zamora Terol in Tesis Doc. Univ. Politèc. Catalunya, 2013 [p.82-83, Table 3]. Issued from : S. Zamora Terol in Tesis Doc. Univ. Politèc. Catalunya, 2013 [p.82-83, Table 3].
Comparison of egg production and clutch sizes among different species of Oithona from field and lab studies.
d: maximum specific-egg production rate.
Also, see: Oithona attenuata, O. colcarva, O. davisae, O. nana, O. aruensis, O. setigera, O. similis, O. simplex. |
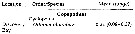 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.168, Table 3]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.168, Table 3].
Mean and range of daily specific growth rates (G, day-1) for nauplii and copepodites (all stages, excluding moult from copepodite stage 5 to adult).
Data from Chisholm & Roff (1990), Webber (1993), Hopcroft & Roff (unpubl.).
Nota: Growth increments for the commonest species in the harbour and off Discovery Bay (Jamaica) were calculated by 2 methods. In the harbour, change in naupliar size and volume over time bwas measured directly from enclosure studies (R. R. Hopcroft & J.C. Roff unpubl.) using the digitising system of Roff & Hopcroft (1986). Dry mass was calculated from relationships of Lawrence & al. (1987). For oceanic waters, consecutive naupliar stages of 12 common species off Discovery bay were digitisede from scaled drawings and volumes calculated. These data were then used with moulting rates to yield growthb rates from:
g = 1/D ln (Vs+1/Vs) or g = 1/D ln (Ms+1/Ms
where D = moulting rate in days; M = dry mass of nauplius; V = volume of nauplius; development stage.
A daily specific growth rate (G) was then derived from : G = e sup>g - 1. (Ms+1/Ms). |
 Issued from : C. Castellani, C. Robinson, T. Smith & R.S. Lanpitt in mar. Ecol/ Progr. Ser., 2005, 285. [p.132, Table 1]. Issued from : C. Castellani, C. Robinson, T. Smith & R.S. Lanpitt in mar. Ecol/ Progr. Ser., 2005, 285. [p.132, Table 1].
Oithona spp. Summary of respiration rates (range) reported in the litterature in different regions.
b: converted to carbon from dry weight assuming body carbon as 40% of dry weight (Mauchline, 1998); c: converted to dry weight from wet weight assuming 81% water content (Mauchline, 1998); d: measurements made during 17 of RV 'Academic Kurchatov' between 05°21' N, 88°05' W and 00°01' S, 139°41' W.
Cf. Oithona similis to compare to other species in Table 1 in the same authors. |
| | | | Loc: | | | Antarct. (SW & SE Atlant., Indian), sub-Antarct. (Indian, SW Pacif.), South Africa (off Cape of Good Hope, E & W), off Tristan da Cunha (N & E), Angola, Baia Farta, off SE St. Helena Is., Congo, off Trinidade Is., off S Ascension Is., G. of Guinea, off Lagos, Ivorian shelf, S Tomé and Principe Is., Brazil (S, Paranagua Bay, Ubatuba, off Rio de Janeiro, Campos Basin, Vitoria Bay, Vitoria-Cabo de Sao Tomé, off Macaé, Mucuri estuary, Guarau estuary, off Natal), Malvinas Current, Argentina, Uruguay (continental shelf), off NE St. Paul Is., Casamance, Dakar, Cape Verde Is., off Mauritania, Morocco-Mauritania, Cap Ghir, Canary Is., off Madeira, NW Spain, Portugal, Mondego estuary, Bay of Biscay, Arcachon Bay, Belon estuary, off Amazon, Barbados Is., Venezuela, Bahia de Mochima, Cariaco Basin, Caribbean Colombia, Venezuela, Caribbean Sea, Jamaica, Yucatan, lagunas de Veracruz, Caribbean, E Costa Rica, G. of Mexico, off W Mississipi River mouth), Florida, Sargasso Sea, off Bermuda: Station S (32°10N, 64°30W), Delaware Bay (outside), Chesapeake Bay, G. of Maine, Bay of Fundy, Ireland, N North Sea, Lübeck Bay, off Spitzbergen (rare), Greenland Sea, Ibero-moroccan Bay, off W Tangier, Gibraltar, Medit. (M'Diq, Alboran Sea, Habibas Is., Sidi Fredj coast, Castellon, Baleares, Algiers, Gulf of Annaba, El Kala shelf, Banyuls, Berre Lagoon, Marseille, Toulon harbour, Ligurian Sea, Tyrrhenian Sea, G. of Napoli, Milazzo, Lake Faro (Sicily), Messina, Gulf of Taranto, Taranto Harbour, NW Tunisia, G. of Gabes, Adriatic Sea, Vlora Bay, Po delta, Trieste, Ionian Sea, Aegean Sea, Thracian Sea, Marmara Sea, Iskenderun Bay, Black Sea, Lebanon Basin, W Egyptian coast, Alexandria, Bardawill Lagoon, Port Said, Suez Canal, Port Taufik, Gulf of Suez, off Sharm El Sheikh, Taba, Hurghada, Safâga, Red Sea, Gulf of Oman, G. of Aden, Arabian Sea, Arabian Gulf, UAE coast, Kuwait, off S Madagascar, Nosy Bé, Rodrigues Is., Seychelles, Kavaratti & Agatti Is. (only outside of lagoon), Indian, Karachi coast, India (Saurashtra coast, W, Lawson's Bay, Porto Novo, Gulf of Mannar, Palk Bay, Godavari region, Kakinada Bay), Bay of Bengal, Andaman Sea, Straits of Malacca, off Phuket, G. of Thailand, Malaysia (Sarawak: Bintulu coast), Sulu Sea, China Seas (Yellow Sea, East China Sea, South China Sea), Taiwan Strait, Taiwan (E, S, W, Kaohsiung Harbor, NW, N, Mienhua Canyon), Okinawa, S Korea, Tsushima Straits, Korea Strait, Japan Sea, Japan, Kuchinoerabu Is., Ariake Bay, Tokyo Bay, Tanabe Bay, Kuroshio zone, Kuril Is., Bering Sea, off S Aleutian Is., Queen Charlotte Is., Strait of Georgia, Pacif. (W equatorial), Australia (G. of Carpentaria, North West Cape, Melbourne, Great Barrier, New South Wales), New Caledonia, New Zealand, Moorea Is, Samoa Is., Pacif. (tropical), California, Baja California (Bahia Magdalena, W), Gulf of California, Bahia de los Angeles, Zihuatanejo Bay, La Paz, W Mexico, Acapulco Bay, G. of Tehuantepec, W Costa Rica, Clipperton Is., Tuamotu Is. (Ahe atoll), Galapagos-Ecuador, Peru, Chile (N-S, Concepcion) | | | | N: | 450 | | | | Lg.: | | | (11) F: 1,2-1,12; (34) F: 1,22-1,1; (35: Rem.) F: 1,38-1,08; (45) F: 1,5-1; M: 1-0,75; (46) F: 1,5-1; M: 0,75; (59) F: 1,2; M: 0,7; (66) F: 1,13; (104) F: 1,2; (109) F: 1,37-1,03; M: 0,8-0,7; (114) F: 1,48; (142) F: 1,4-1; M: 1-0,75; (180) F: 1,35-1,18; M: 0,79-0,66; (208) F: 1,5-1,4; (237) F: 1,1-1,9; M: 1,2; (246) F: 1,35-1,15; (327) F: 1,54-1,16; (332) F: 1,3-1,21; M: 1,008; (333) F: 1,44-1,17; (334) F: 1,5-1; M: 1-0,75; (336) F: 1,24-1; M: 0,63-0,65; (373) F: 1,26-1,06; (449) F: 1,5-1; M: 1-0,75; (530) F: 1,1; (531) F,M: 0,93; (624) F: 1,28-1,06; M: 0,68-0,59; (634) ? F: 1,37-1,2; (649) F: 1,21; M: 0,71; (651) F: 1,23-1,1; M: 0,66-0,61; (654) F: 1,314-1,116; M: 0,95; (785) F: 1,15-0,83; (786) F: 1,39-1,12; (880) F: 1,0-1,5; M: 0,6-1,0; (920) F: 1,24; (937) F: 1,12-1,19; (991) F: 1,1-1,5; M: 0,75-1; (1085) F: 1,0-1,3; M: 0,75-0,8; {F: 0,83-1,54; M: 0,59-1,01} | | | | Rem.: | Marine, sometimes brackish.
Epi-mesopelagic-bathypelagic.
Overall Depth Range in Sargasso Sea: 0-500 m (Deevey & Brooks, 1977, Station "S"). Sampling depth (Antarct., sub-Antarct.) : 0-1000 m. 0-3010 m at Station T-1 (E Tori Is, E middle Japan) from Furuhashi (1966 a).
Gurney (1927, p.158) attributed this species, which occurs in Suez Bay, to O. plumifera in spite of no trace of the plumose setae which are characteristic of it. In every case the seta of the baipodite of the legs is smooth. Farran (1913, p.183) mentions that some among his Christmas Island specimens lacked the feathering of these setae (probably broken off). Generally this feathering is said to be coloured red, but it may be colourless. In all other respects this form differs from O. spinirostris, with which it might be confused, in having 3 setae only on the endopod of the Md and also in the length of the seta on the endopod of Mx1. O. tenuis Rosendorn (1917) does not seem to differ from O. plumifera in any essential character except in having only 2 setae of endopod of Md.
Sewell (1947, p.255, female) notes: The proportional lengths of the segments of the posterior region (Th 5 to caudal rami) as 12 : 31 : 16: 17 : 14:10 = 100. The segments of A1 are fringed with small spinules. In some ot the specimens the plumose setae arising from the swimming legs are present, but in others they were either present only on the anterior two pairs of legs or were apparently absent altogether. It seems probable that the plumose termination is a variable character and my in some cases wanting.
Nishida (1985, p.71) reports that O. plumifera is clearly distinguished from O. atlantica by trhe presence of a ventral tuft of setules on genital segment (noticed by Rosendorn, 1917, but overlooked by Farran and Früchtl), and the curved marginal spine of the exopodal segment 3 of P4. O. tenuis is distinct from O. plumifera in having 1 inner marginal seta on the exopod of P2-P4. Olson (1949) described a second short seta on the free segment of P5 in some specimens from California coast; this character has not been reported by any other workers. After Björnberg (1963, p.74) this form is one of the most common species.
Tanaka (1960, p.62) had observed a female specimen from a fresh water in Okino Erabu Shima (Anami Is., Japan) of body size 1.13 mm.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 6 corresponding to small ambush feeding omnivorous.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 28, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
