|
|
 |
|
Monstrilloida ( Order ) |
|
|
|
Monstrillidae ( Family ) |
|
|
|
Cymbasoma ( Genus ) |
|
|
| |
Cymbasoma rigidum Thompson, 1888 (F,M) | |
| | | | | | | Syn.: | Thaumaleus rigidus : T. Scott, 1904 (p.248, figs.F, Rem.); Pearson, 1906 (p.33); van Breemen, 1908 a (p.213, figs.F); Pesta, 1920 (p.634, fig.F); Rose, 1925 e (p.307); 1927 f (p.18); Davis, 1949 a (p.250); Shih & al., 1971 (p.57); Isaac, 1974 (p.130); 1974 a (p.133, Rem.); 1975 (p.3, 7, 8, figs.F,M); Tremblay & Anderson, 1984 (p.8, as T. rigidum); Dias de O., 1996 (p.255, Rem.);
? Thaumaleus claparedii Giesbrecht, 1892 (p.578, 585, figs.F);
Non Thaumaleus germanicus Timm, 1893 (p.419); van Breemen, 1908 a (p.213, figs.F); Caullery & Mesnil, 1914 (p.19, figs.F, M);
Thaumaleus germinacus : Razouls, 1981 (lapsus calami);
Monstrilla ostroumowi Karavaev, 1894 (1895) (p.33, figs.F);
Monstrilla rigida : Bourne, 1890 b (p.575, figs.F);
? no Haemocera danae Malaquin, 1901 (p.88, 109, fig.F: Pl II, figs. 1, 2, 3, 6); Sewell, 1949 (p.144, figs.F);
Haemocera danae : Razouls, 1973 (p.467); Razouls & Durand, 1991 (p.75) | | | | Ref.: | | | Thompson, 1888 b (p.154, Descr.F, figs.F); 1888 d (p.149); Sars, 1921 (p.21, figs.F,M); Wilson, 1932 a (p.396, figs.F); Rose, 1933 a (p.348, figs.F,M); Porumb, 1961 (p.1224, figs.F,M); Threlkeld, 1977 (p.227); Sekiguchi, 1982 (p.28, figs.F); Grygier, 1995 a (p.45, 74); Chihara & Murano, 1997 (p.1002, Pl.234: F); Suarez-Morales, 2001 (p.247: Rem.); Bernier & al., 2002 (p.651, tab.1, 652, figs.F, Rem.) ; Vives & Shmeleva, 2010 (p.166, figs.F,M, Rem.); Suarez-Morales, 2011 (p.6: Rem) | 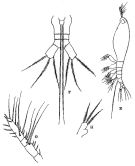 issued from : R.B.S. Sewell in The John Murray Expedition, 1933-34, Scientific Reports, IX (2), 1947. [p.143, Fig.40 E-H]. As Haemocra danae. With doubt. Female (from Nankauri Harbour, Nicobar Is.): E, habitus (lateral left side); F, urosome (ventral); G, A1; H, P5. Nota: Head and 1st thoracic segment fused. Mouth situated far forward near the anterior end; the pre-oral and post-oral parts of the cephalon being in the ptoportion of 12 to 43. A pair of well-developed eyes, with hemispherical lenses and brown pigment, occupied a grat prt of the anterior region of the head (dorsal view). Proportional lengths of the various segments of the body as: 562: 90:82:86:56:60:15:15:34 = 100. Genital segment twice the length of the two following segments together. Caudal rami divergent, slightly longer than the two preceeding segments together, each ramus bears 3 setae, of approximately equal length. A1 are about equal in length to the pre-oral region of the cephalon;, composed of 4 segments: 12:28:14:46 = 100. P5 have a rounded projection on the inner border about half-way along their length, no seta arises from the inner projection, the distal end carries 3 setae of about equal length
|
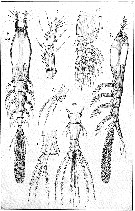 issued from : G.O. Sars in An Account of the Crustacea of Norway, with short descriptions and figures of all the species, 1921 a, 8. [Pl. XI]. Female (from West coast of Norway). Urs, urosome (dorsal). Nota : Cephalic segment fully as long as the remaining part of the body and, viewed dorsally, somewhat dilated in the middle, ventral face evenly convex throughout. Tail scarcely exceeding half the length of the exposed part of the trunk. Eye well developed, with all 3 lenses distinct. A1 rather short, scarcely attaining 1/3 of length of the cephalic segment, terminal segment somewhat shorter than the other 3 combined, with none of the setae ramified. Oral tubule small, occurring at about the end of the first ¼ of the cephalic segment. Genital segment narrowed behind ; ovigerous spines nearly 3 times as long as the tail ; distal segment somewhat flattened and gradually widening behind, exhibiting in front of the middle, on each side, a slight notch, as an attempt to a subdivision, the notch not being continued in any difining suture. Caudal rami slightly longer than they are broad, and not much divergent with the inner edge somewhat concaved, the outer gibously produced, each ramus provided with 3 strong subequal setae, the outermost of which is attached to a rather prominent edge ; a very small appendicular bristle is generally found attached near the inner corner to the ventral face. Natatory legs with the outer ramus only slightly longer than the inner, its terminal segment not much produced. Last pair of legs with 3 apical setae, the innermost much smaller than the other 2, inner edge produced to a narrow linguiform lobe curving outwards along the terminal part of the leg, and in most cases extending as far as the latter.
|
 issued from : G.O. Sars in An Account of the Crustacea of Norway, with short descriptions and figures of all the species, 1921 a, 8. [Pl. X, 2]. Male (from West coast of Norway). Urs, urosome (ventral). Nota : Smaller than female and of shorter and more robust form of the body, with the cephalic segment somewhat clavate in outline. Tail rather narrow, 3-segmented, the last segment exhibing on each side a quite similar notch to that observed in female. Copulative appendage divided at the end into 2 comparatively large diverging lobes of a somewhat sausage-shaped form. Caudal rami more pronouncedly clavate than in female, eacg provided with 4 slender subequal setae attached to the obtusely truncated and distinctly thickened extremity, one of them apparently answering to the small appendicular bristle found in the female. A1 much more elongate than in female, 5-segmented, last segment very movably articulated to the preceding one, and terminating in a slender, slightly curved spine. Natatory legs of same structure as in female. P5 not detected.
|
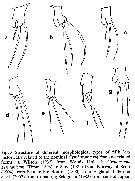 issued from : E. Suarez-Morales in Helgol. Mar. Res., 2006, 60. [p.178, Fig.5]. Structural patterns of the P5 of C. rigidum reported and depicted in the litterature between 1890 and 2002. Nota: There are strong variations in this appendage, the main ones were found on the length and development of the inner lobe. 1 - structure very long, reaching the distal end of the outer lobe as in C. rigidum (recorded as Monstrilla rigidum by Bourne (1890) from Plymouth and Jersey, England. 2 - structure extremely short, weakly developed inner lobe, as in Scott (1904) from Scotland, G. of St. Lawrence (Canada). 3 - structure intermediate reported by Sekiguchi (1982) from Ago Bay (Japan). Another variable character of the P5 is the relative length of the inner terminal seta of the outer lobe; it can be nearly as long as the other setae, as in (Timm, 1896), and in reports of C. rigidum by Bourne (1890) and Scott (1904). A short inner seta (about half the length of the other two setae) is present in the specimens reported by Sars (1921), C.B. Wilson (1932), Sekiguchi (1982) and bernier & al. (2002). It seems unlikely that all these mixed patterns can be reliably associated to a single species. It is necessary to obtain and analyse specimens from the type locality (Madeira, Canary Islands) in order to establish a neo-type of C. rigidum.
|
 issued from : E. Suarez-Morales in Helgol. Mar. Res., 2006, 60. [p.177, Table 1]. Comparative analysis of characters used in the identification of Cymbasoma rigidum and related forms.
|
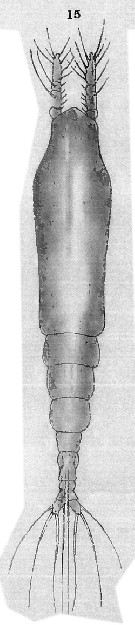 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XIII, fig.15]. As Thaumaleus rigidus. Female (Scotland): 15, habitus (dorsal). Nota : Cephalothorax moderately stout, rather wider near the anterior end, and nearly twice as long as the entire length of the remaining segments.
|
 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XIII, figs.16-17]. As Thaumaleus rigidus. Female: 16, A1; 17, P5. Figs. 16 and 17 at the same scale. Nota: A1 short and stout, appear to be 4-segmented, the 1st and 3rd segments being very small. P5 rather larger than broad, terminal portion distinctly small and wider than the proximal half ; each foot carries 3 moderately long apical setae.
|
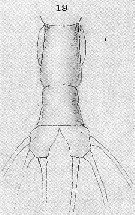 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XIV, fig.19]. As Thaumaleus rigidus. Female: 19, abdomen and caudal rami. Nota: Abdomen 2-segmented ; 1st segment about one and a half times the length of the last segment of the thorax and the proximal half more or less dilated ; 2nd segment, which is smaller than the first, probably composed of 2 coalescent segments, as a slight constriction, dividing the segment into 2 portions. Caudal rami of moderate length, each fusnished with 3 setae.
|
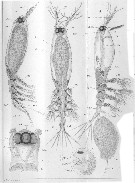 Issued from : A. Malaquin in Arch. Zool. Exp. Gen., 1901, 3è ser., IX. [P.221, Pl. II, figs.1, 2, 3, 7]. As Haemocera danae doubtful. Female (from Roscoff): 1, habitus (lateral); 2a, habitus (dorsal); 2b, forehead with eyes and oral aperture (b) (ventral view); 3, habitus (lateral) with ovigerous spines and egg masses; 7, nauplius before hatching. an 1 : Antennule; Moc : ocular muscle; cer : brain; St : stomodeum; Ovd : oviduct; Pth 5; ab ; Oel : lateral eye; oem : median eye; Ov : oovary.
Nota:
- Number of abdominal segments between the genital segment and the caudal rami : 2.
- Number of caudal setae : 4.
- Number of antennular segments : 5.
- Localization of the mouth : foremost.
| | | | | Compl. Ref.: | | | I.C. Thompson, 1888 d (p.149); C.B. Wilson, 1950 (p.197); Marcus, 1970 (p.11); Hure & Krsinic, 1998 (p.98, 105); Holmes, 2001 (p.61); Kazmi, 2004 (p.231); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Dias & Bonecker, 2007 (p.270, 272, fig.3, tab.II); Oliveira Dias & Costa Bonecker, 2007 (p.282: Rem); Oliveira Dias & al., 2008 (p.248, tab.1) | | | | NZ: | 12 | | |
|
Distribution map of Cymbasoma rigidum by geographical zones
|
| | | | | | | | | | | | | Loc: | | | Guinea-Bissau, Canary Is. (Tenerife), Brazil (Vitoria Bay, Natal-Sao Sebastiao, Bahia), G. of Maine, off Woods Hole, Bay of Fundy, S Gulf of St. Lawrence, W & E Ireland, W Norway, Firth of Clyde, Firth of Forth, North Sea, English Channel, Medit. (Algiers, Banyuls, Malta, N Adriatic Sea, Black Sea), Arabian Sea, Philippines, Viet-Nam, Japan, Ago Bay, Polynesia (Takapoto Atoll), G. of Alaska | | | | N: | 20 ? | | | | Lg.: | | | (45) F: 2,5-2,2; M: 1,75-1,5; (449) F: 2,7-2,2; (663) F: 2,5; M: 1,75; (723) F: 1,4; (727) F: 2,7; (734) F: 3,175; (735) F: 0,88-0,75; M: 0,79-0,57; (907) F: 2,16; (995) M: 0,93*; (997) M: 0,52-1,13*; {F: 0,750-3,175; M: 0,520-1,750}
*: caudal rami excluded. | | | | Rem.: | After T. Scott (1904, p.249), the author has adopted Thompson's name for his form in preference to that of Giesbrecht, beeing convinced that Thaumaleus rigidus (I.C. Thompson) and T. claparedii are identical.
According to Sewell (1949, p.145) the synonymy establishes by Sars (1921, p.21) between Haemocera danae and Cymbasoma rigidum is not valid.
For Isaac (1974, p.137) the development was described by Malaquin (1901) as Haemocera danae in the gregarious serpulid polychaete Salmacina dysteri (Huxley).
Dias' specimens collected off Spirito Sancto (Brazil): T° = 20.60-27.00 °C; Salinity = 29.17-36.80 x 10-3; Dissolved O2 = 3.91-7.90 mg L-1.
For Suarez-Morales (2006, p.171, Rem.: p.178-179) Cymbasoma rigidum would have to be assigned to different species or represents a complex of different morphotypes. The morphology of P5 reported in the litterature shows a strong variation ( length and development of the inner lobe; relative length of the inner terminal seta of the outer lobe; the length of A1 ) as its cosmopolitan geographical range. It seems unlikely that all these mixed patterns can be reliably associated to a single species. After the authors (2011, p.6) the nominal species is known to contain at least four distinct taxa and most records should be revised.
See remarks in Cymbasoma davisi.
For Suarez-Morales & al. (2017, p.1804), the incomplete original description of C. rigidum by Thompson (1888) from the Canary Islands is an obstacle to morphologically define the true C. rigidum. Among the discernible characters of the original description, the large, unconstricted anal somite, representing about 70 % of the genital double-somite length, the unconstricted genital double-somite with weakly expanded anterior half, the strongly pigmented inner half of the medially conjoined lateral eye cups, and the relatively short antennulary element 3, allow some comparison.
The structure of P5 was incompletely described by Thompson (1888), but Bourne (1890) stated that his specimens from Plymouth are identical to those from the Canary Islands and added that they have relatively long endopodites; thus, he illustrated the Plymouth specimens as having a well-defined P5 inner lobe and 3 equally long setae on the outer lobe. So, a relatively large, unconstricted anal somite is an additional character related to the true C. rigidum. This strict pattern diverges in one or more characters from the reports assignated to this species by Scott (1904), Sars (1921), Wilson (1932) and Bernier & al. (2002), which should be revised (see above, Table 1, p.177, Suarez-Morales, 2006). | | | Last update : 28/07/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed November 18, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
