|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Pseudodiaptomidae ( Family ) |
|
|
|
Pseudodiaptomus ( Genus ) |
|
|
| |
Pseudodiaptomus inopinus - species complex Burckhardt, 1913 (F,M) | |
| | | | | | | Syn.: | Non Pseudodiaptomus japonicus Kikuchi,1928;
Schmackeria inopinus : Marsh, 1933 (p.44, figs.F,M, Rem.); Shen & Tai, 1962 (p.101, Rem.); Shen & Lee, 1963 (p.578, Rem.); Chen & Zhang, 1965 (p.81, figs.F); .
Non Pseudodiaptomus inopinus : Smirnov, 1929 (p.318, fig.F, M); Tanaka, 1966 (p.42, fig.F); Mashiko & Inoué, 1952 (p.184, fig.F, M); Kos, 1976 (Vol.II, figs.F,M, Rem.); Kikuchi & al., 1978 (p.25, fig.F, M); Kos, 1984 (1985) (p.239, fig.F, M, Rem.); Oka & al., 1991 (p.85, fig.F); Chihara & Murano, 1997 (p.893, fig.F, M); Eyun & al., 2007 (p.265, molecular biology); Lee & al., 2007 (p.140, fig.F, M); Sakagychi & Ueda, 2010 (p.62, fig.F, M, Rem.); 2011 (p.124, fig.F, M, Rem.); Soh & al., 2012 (p.233, fig.F, M, Rem). | | | | Ref.: | | | Pseudodiaptomus inopinus sensu stricto:Burckhardt, 1913 (p.379-394, Figs.F, M: 11E, 12H, Rem. p.392: phylogeny); Sewell, 1948 (p.421); Walter, 1986 (p.132); 1986 a (p.503); 1987 (p.367); Walter & al., 2006 (p.203, Rem.; Ueda & Sakaguchi, 2019 (p.35, descr. F, M, Rem.). |  issued from : H.Y. Soh, H.-L Suh, O.H. Yu & S. Ohtsuka in Hydrobiologia, 2001, 448. [p.218, Fig.3, C]. Female (from W Korea): scanning electron micrographs of the genital area.
|
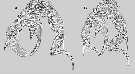 issued from : J.R. Cordell, M. Rasmussen & S.M. Bollens in Mar. Ecol. Prog. Ser., 2007, 333. [p.216, Fig.4]. Male (from Chehalis River estuary): P5 of 2 morphotypes; A, paddle-shaped; B, thumb-shaped.
|
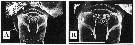 issued from : T.C. Walter, S. Ohtsuka, S. Putchakarn, K. Pinkaew & S. Chullasorn in Proc. Biol. Soc. Washington, 2002, 115 (3). [p.664, Fig.9, A-B]. Female: genital double somite (ventral); B, same (magnification). Scale bars: 0.1 mm.
|
 Issued from : H.Y. Soh, S.W. Kwon, W. Lee & Y.H. Yoon in Zootaxa, 2012, 3368. [p.233, Fig.3]. Female: SEM photographs of the genital double-somite from P. koreanus (A) [= P. japonicus] and P. inopinus (B). White arrows indicate process between gonopores; black arrows indicate the posterior process of genital flap.
|
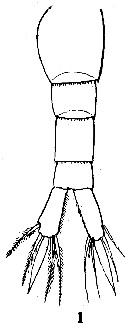 Issued from : C.D. Marsh in Proc. U.S. natn. Mus., 1933, 82 (18) (2959). [Pl. 22, fig.1]. As Schmackeria inopinus. Female (from Yangtze Kiang): 1, urosome dorsal).
|
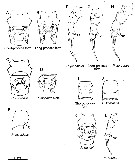 Issued from : S.O. Sakaguchi & H. Ueda in Plankton Benthos Res., 2018, 13 (4). [p.174, Fig.1]. Published illustrations of Pseudodiaptomus inopinus species complex (A-D, K) genital operculum, ventral view; (E) genital double-somite, right lateral ,view; F-H, L) female left P5; (I, J) ventral surface of the male 2nd urosomite. Sources of figures: A, B and G, Sakaguchi & Ueda (2011), 2A-C, respectively; C, H and J, Soh & al (2012), figure 2C, 6B and 7C, respectively; I, K and L, Sakaguchi & Ueda (2010), figure 8C, 2C and 5E, respectively; D, Burckhardt (1313), figure 9 of plate 11F; E and F, Kikuchi (1928), figures 13 and 14 of plate 19, respectively. Arrowheads a, b and c indicate the posterior process of the genital operculum, the central part of the genital flap surrounding gonopores, and the tip of the distomedial process on the 1st exopodal segment , respectiively. Scale bar of 0.1 mm is applicable for A-C and G-I. Only as ''LP-form'' = P. yamato.
|
 Issued from : S.O. Sakaguchi & H. Ueda in Plankton Benthos Res., 2018, 13 (4). [p.178, Table 2]. Taxonomic status of Pseudodiaptomus japonicus and related references. Only ''LP-form\"\" = P; yamato.
|
 Issued from : G. Burckhardt in Zool. Jb., Syst., 1913, 34 (4). [p.379-394; Rearranged after Ueda (pers. comm., 2019): Taf. 11E, figs.2, 3, 5, 7; Taf.11. 11F, fig.9; Taf. 12, figs.1-2, 7-8]. Female: A, habitus (dorsal) with eggs sacs; B, posterior region of prosome; C, genital operculum; D, caudal ramus; E, P5. Male: F, habitus (dorsal); G, H, P5.
|
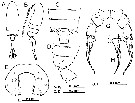 Issued from : H. Ueda & S.O. Sakaguchi in Plankton & Benthos Res., 2019, 14 (1); [p.36, Fig.4, A-E, G, H]. Female (from Lake Tahiu, China): A-B, habitus (dorsal and lateral, respectively); C-D, 3rd pediger to genital double-somite (dorsal and lateral, respectively); E, last pediger (posterior); G, P5; H, right leg of P5. Nota; - 2nd and 3rd pedigers with row of conspicuous spinules along posteromedial margin. - Last pediger with prominently protruding posterior projection and 3 groups of spinular rows on each side, i.e. row of slightly curved long spinules on distolateral corner, rows of medium or fine spinules on distomedial surface fig.4E), and short row of fine spinules on lateral surface (fig.4I, indicated by arrow). -P5 with group of a few spinules ayt midlength on medial margin of 1st exopodal segment.
|
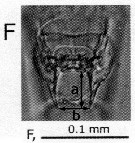 Issued from : H. Ueda & S.O. Sakaguchi in Plankton & Benthos Res., 2019, 14 (1); [p.36, Fig.4, F]. Female: F, genital operculum. Nota: Genital operculum with long posterior processes; process slightly longer than distance between tips (fig.4F, 'a' > 'b').
|
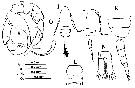 Issued from : H. Ueda & S.O. Sakaguchi in Plankton & Benthos Res., 2019, 14 (1); [p.36, Fig.4, I-L, N, O]. Male (from Lake Taihu, China): I-J, habitus (dorsal and lateral, respectively); K, last three pedigers and urosome (lateral); L, 2nd urosomite (ventral); N, anal somite and caudal rami (dorsal); O, P5 (posterior). Nota: - 2nd and 3rd pedigers with row of conspicuous spinules along posterolateral margin. - 2nd urosomite with spinulation (fig.4L). - Caudal ramus length 2.8 times width. - P5 as fig. 4O.
|
 Issued from : H. Ueda & S.O. Sakaguchi in Plankton & Benthos Res., 2019, 14 (1); [p.36, Fig.4, L, M]. Male: L, 2nd urosomite (ventral); M, photograph of ventral spinulal rows in L. Nota: 2nd urosomite with similar spinulation to P. yamato, but lateral spinular row more conspicuous than P. yamato, and spinules of transverse ventral row (fig.4L, M) as large as those of oblique ventrolateral row.
| | | | | Compl. Ref.: | | The references following does not account the synonymies from this species complex.
Cordell & al., 1992 (p.260); 1992 a (p.171, occurence); Ohtsuka & al., 1995 (p.159); Cordell & Morrison, 1996 (p.629, invasive species); Cordell & al., 1997 (p.2, invasive species); Mauchline, 1998 (p.507); Hairston & Bohonak, 1998 (p.23, Table 3, genetic); Uye & al., 2000 (p.193, fig.7 : abundance vs t°, Sal.); Humphrey, 2008 (p.64, 84: Appendix A); Ohtsuka & al., 2008 (p.115, Table 4, 5, Rem.: in Osaka Harbour); Sakaguchi & al., 2011 (p.18, Table 1, 2, occurrences); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Moon & al., 2012 (p.1, Table 1); Suzuki, K.W. & al., 2013 (p.15, Table 2, 3, 4, fig.6, estuaries, annual occurrence); Park E-O. & al., 2013 (p.1, annual variation, sex ratio); Sabia & al., 2015 (p.460, Table 3, Rem.); Ohtsuka & Nishida, 2017 (p.578, 579, Rem.) | | | | NZ: | 4 | | |
|
Distribution map of Pseudodiaptomus inopinus - species complex by geographical zones
|
| | | | | |  Chart of 1996 Chart of 1996 | |
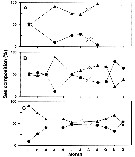 Issued from : E-O. Park, J.R. Cordell & H.Y. Soh in Zool. Stud., 2013, 52 (7). [p.7, Fig.7]. Doubtful identification of the species. Issued from : E-O. Park, J.R. Cordell & H.Y. Soh in Zool. Stud., 2013, 52 (7). [p.7, Fig.7]. Doubtful identification of the species.
Seasonal variations in the adult sex composition of P. inopinus from Mankyung River estuary (South Korea).
(A): Oligohaline, (B): mesohaline, (C): polyhaline. Circle : female; triangle: male.
Nota: Males were more abundant than females throughout the study period (January to December 2000) and across the entire range of salinity, but the ratio of males to females exceeded 50% under oligohaline conditions in January, mesohaline conditions in late fall and winter, and polyhaline conditions in early winter..
Gravid females appeared in oligohaline conditions in March, July, and September and accounted for 50% of all females present in September. Under mesohaline conditions, they occurred in June to October, and under polyhaline conditions, they occurred only in June and October, and >50% of all females were gravid in October.
These facts indicate that abundance of P. inopinus as P. poplesia may be controlled by Chl a concentrations and salinity conditions. |
| | | | Loc: | | | China Seas (Bohai Sea, Yellow Sea, East China Sea, South China Sea), Yangtze-Kiang Riv.), Korea (S & W), Muan Bay, Mankyung River estuary, E & W Japan, south estuaries: Chikugo, Midori & Kuma Rivers), Nansei Is., Sakhalin Is., British Columbia (Columbia River estuary).
Type locality: Lake Tahiu (China) | | | | N: | 23 ?? | | | | Lg.: | | | (97) F: 1,28-1,22; (290) F: 2-2,2; (866) F: 1,1-1,5; M: 0,9-1,36; (1062) F: 1,40; 1,36; (1099) F: 1,38-1,44; M: 1,10-1,15; (1232) F: 1,01-1,22; M: 1,1; {F: 1,01-2,28; M: 1,1-1,36} | | | | Rem.: | freshwater, brackish.
For Marsh (1933, p.45) this species resembles S; (= P.) forbesi so closely that is somewhat difficult to separate them and one might be inclined to think of inopinus as simply a variety of forbesi.
For Shen & Tai (1962, p.117, 118) this species occurs at the mouths of the big rivers in China as well as in Japan, yet it quite different in form from the typical estuarine species, such as P. poplesia.
In Lobus species group (forbesi subgroup) after Walter & al. 2006, p.203).
Introduced from Asia in rivers of Washington State and in British Colombia by transport in ballast water of ships.
According to Cordell & al., 2008 (p.753) the 1st record of this species in Columbia River (USA) is 1990.
After Soh & al. (2012, p.238) this species is very closely related to P. koreanus and P. nansei.
For Sakaguchi & Ueda (2010, p.52) this species is closely related P. nansei.
After Ohtsuka & Nishida (2017, p.567) this species dominates brackish waters of Japan, but its populations in the Nansei Islands, southernmost Japan, proved to represent a distinct sibling species, P. nansei. These authors, point to in the Inlet and brackish waters on the main islands of Japan, this species is dominant. Also, After Ohtsuka & Nishida (2017, p.579), the brackish species , aggregated just above the bottom in submerged channels outside the mouth during heavy flooding. Losses from the population, in particular of immature copepodids, were compensated by reproduction after the flood.
After genomic analysis, Sakaguchi & Ueda show that P. inopinus is a species complex, located in japanese estuarines.; these two allopatric forms are distinguishable by the length of the posterior processes of the genital operculum called SP- and LP-forms, respectively on the female, with a long lprocess for P. inopinus s.str. and a short process for P. japonicus Kikuchi, 1928.
The phylogenetic results revealed that P. koreanus from Korea belonged to the same clade as the SP-form, indicating that P. koreanus is the same species as P. japonicus, but a comparison between the two forms are in preparation.
The authors (2010) indicated morphological and genetic differences between P. nansei which was once a cryptic species of P. inopinus and P. japonicus (as P. inopinus from the Tsuri-kawa River, W Kyosho, japan). Remarkable morphological features of P. nansei are shorter posterior processes of the genital operculum than those in P. japonicus (See Fig.1 K) and a pointed distomedial process of the 1st exopodal segment of the female P5 (see fig.1 L), in addition to thin lateral medial terminal caudal setae in the female.
See remarks in P. yamato and P. japonicus
The type locality from Pseudodiaptomus inopinus Burckhardt, 1913 is the Lake Tahihu, China.
For Ueda & Sakaguchi (2019, p.36) P. yamato differs from P. inopinus s.s. by having weakly protruding posterior projections on the female last pediger and lacking medial spinules at the midlength of the 1st exopodal segment of the female P5
P. japonicus differs from P. inopinus s.S. by having the short posterior processes of the female genital operculum and the weakly protruding posterior projections on the female last pediger.
The males of P. yamato and P. japonicus differ from P. inopinus s.s. by spinules of the transverse ventral row on the 2nd urosomite, which are larger in P. inopinus sensu stricto. | | | Last update : 19/06/2023 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 29, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
