|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Acartiidae ( Family ) |
|
|
|
Acartia ( Genus ) |
|
|
|
Acanthacartia ( Sub-Genus ) |
|
|
| |
Acartia (Acanthacartia) californiensis Trinast, 1976 (F,M) | |
| | | | | | | Syn.: | Acartia sp.I Uye & Fleminger, 1976 (p.253, resting egg, hatching). | | | | Ref.: | | | Trinast, 1976 (p.54, figs.F,M); Dawson, 1979 (1980) (p.1, figs.F,M); Dawson & Knatz, 1980 (p.8, figs.F,M); Gardner & Szabo, 1982 (p.420, figs.F,M); Trujillo-Ortiz, 1986 (p.188, figs. Nauplius, juv,F,M); Trujillo-Ortiz & al., 1995 (p.39, genetic); Ferrari & Dahms, 2007 (p.71, Rem.) |  issued from : E.M. Trinast in Crusraceana, 1976, 31 (1). [p.55, Figs.1-10]. Female (from 33°37'10''N, 117°53'40''W): 1-2, habitus (dorsal and lateral, respectively); 3, forehead (ventral); 4, A1; 5, Md (masticatory edge); 6, P4; 7, P5; 8, idem (lateral); 9, urosome (dorsal); 10, idem (lateral). Scale bars for 1 represents 0.5 mm; those for 5 and 7 = 0.05 mm. Nota : Rostral filaments long and slender. A1 reacing just beyond end of prosome. Prosome :urosome = about 3.2. Genital segment longer than wide. Ornamentation (presence of spines, hairs or setae variable ; when present, posterolateral margin of last prosome segment armed with 2 small spines on each side, genital segment and suceeding segment with row of minute spinules on posterodorsal margins. Anal segment short, without spines or setae. Caudal rami longer than wide (length : width = about 1.6). Segment 1 of P5 fused medially, segment 2 longer than broad, segment 3 (terminal spine) bulbous posteriorly at base, spinules mimited to central part (distal segment often curved at tip) ; external seta nearly as long as terminal spine.
|
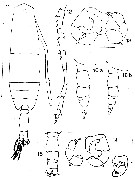 issued from : E.M. Trinast in Crusraceana, 1976, 31 (1). [p.56, Figs.11-16]. Male: 11, habitus (dorsal); 12, A1; 13, P5 (posterior surface); 14, idem (anterior surface); 15, urosome (dorsal); 16 a, 16b, urosome (lateral). Scla bars: for 11 represents 0.5 mm; those for 13 and 14 = 0.05 mm. Nota Prosome : urosome = abiut 2.9. A1 extending about to end of last prosome segment. Ornamentation variable, when present, last prosome segment armed with small dorsal spine and several (3-6) more slender, ventral setae . 1st urosomal segment narrower than 2nd ; 2nd, 3rd and 4th urosomal segments each with row of minute spinules on posterodorsal margin ; anal segment not ornamented. Each basal segment of P5 with plumose seta on outer margin; basal segment of right P5 with triangular inner lobe, exopodite 1 with proximal inner lobe bearing small spine, exopodite 3 with a large, bilobed inner projection bearing small spine on distal margin, exopodite 3 outer margin with 3 or more small hairs near curved region, inner margin smooth except for small spine near midpoint, segment terminating in a small spine; left P5 exopodite 1 without spines or seta, exopodite 2-3 terminating in long, blunt apical spine, slightly fexed, sustended by slender, simple accessory spine.
|
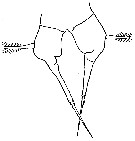 issued from : J.K. Dawson in Harbors Environmental Projects, Univ. Southern California, 1979. [p.10, Fig.4]. Female (from Los Angeles Harbors): P5.
|
 issued from : J.K. Dawson in Harbors Environmental Projects, Univ. Southern California, 1979. [p.8, Fig.2]. Male: P5.
|
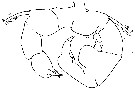
 issued from : J.K. Dawson in Harbors Environmental Projects, Univ. Southern California, 1979. [p.15, Fig.9, A, C]. Female: C, urosome (dorsal). Male: A, urosome (dorsal).
|
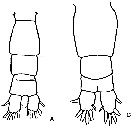
 issued from : J.K. Dawson in Harbors Environmental Projects, Univ. Southern California, 1979. [p.4, Table 1]. Differential morphometrics of Acartia californiensis and Acartia tonsa in the Los Angeles Harbors (California). Nota: 20 females and 20 males of each species were studied. In male, he small digitiform appendages on the left terminal segments of A. tonsa clearly differentiate it from the single large digitiform appendage on the same segment of A. californiensis. Similarly, the presence and absence of a lobe on the 2nd basipodal segment of female A. tonsa and A. californiensis respectively, separate the females of these species. A search for an easily observed and reliable character for differentiating these two species was found fro females in the width to length ratio of the genital segment and for males in the width to length ratio of the 2nd urosomal segment.
| | | | | Compl. Ref.: | | | Johnson J.K., 1980 (p.568, production & hatching of dormant eggs); Grice & Marcus, 1981 (p.125, Dormant eggs, Rem.: p.131, 135, 136); Brinton & al., 1986 (p.228, Table 1: Rem.); Trujillo-Ortiz, 1990 (p.1, Table I, II, III, IV, V, figs. 1, 2, 3, 4, 5: population dynamic); Huntley & Lopez, 1992 (p.201, Table A1, egg-adult weight, temperature-dependent production); Kimmerer, 1993 (tab.2); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Marcus, 1996 (p.143); Cordell & al., 1997 (p.7, % change vs 1980-1997); Suarez-Morales & Gasca, 1998 a (p107); Mauchline, 1998 (tab.40, 45, 47, 51, 61); Elliott & Kaufmann, 2007 (p.418); Galbraith, 2009 (pers. comm.); Bollens & al., 2011 (p.1358, Table III, fig.6); Gusmao & al., 2013 (p.279, Table 3, sex-specific predation by fish); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, pers. comm.) | | | | NZ: | 2 | | |
|
Distribution map of Acartia (Acanthacartia) californiensis by geographical zones
|
| | | 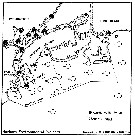 issued from : J.K. Dawson in Harbors Environmental Projects, Univ. Southern California, 1979. [p.16, Fig.10]. issued from : J.K. Dawson in Harbors Environmental Projects, Univ. Southern California, 1979. [p.16, Fig.10].
Distribution of A. californiensis and A. tonsa in Los Angeles-Long Beach Harbors, California. November 1978.
Nota: As can be seen from the distribution of the two species, A. californiensis appears to dominate most of the inner habor channels and A. tonsa predominates in the outer harbor.
There is an mixing of the two species, probably by tidal action.
The strongest and most consistent abiotic differences between the inner and outer harbor are in pH and dissolved oxygen; inner harbor with low pH (7.75) opposed to the outer harbor (8.02) and also a lower dissolved oxygen (6.1 ppm) compared to the outer harbor (8.02 ppm). But whether or not such abiotic characteristics are responsible for the observed distribution of these species is yet to be fully determined. |
 Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.10, Table I]. Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.10, Table I].
Number of eggs deposited in a day period by 100 fertilized females of A. californiensis distributed in five replicates each with 20 females.
Mean (x) and standard deviation (SD).
Individuals collected in the Estero Punto Banda (Todos Santos Bay, Ensenada, Baja California, Mexico): 31°42'-31°47' N, 116°37'-116°40' W. in Febrero 1983..
Experimental conditions: 17°C ±1°C, feeding: Isochrysis tahitiana and Tetraselmis sp., in equal part, concentration of 7.5 x 10,000 cells/ml.
Concentration in the bay: 5.7 individuals per liter, but 48 ind. per liter in the laboratory; perhaps explaines the little difference in the isochronal development between the experiences and the Bay. |
 Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.10, Table II]. Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.10, Table II].
Number of daily eggs produced by two females of A. californiensis in a 10-day period.
Mean (x) and standard deviation (SD). |
 Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.11, Table IV]. Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.11, Table IV].
Hatching success of 250 eggs of A. californiensis (five replicates) during a 10-day period.
Mean (x) and SD. |
 Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.15, Table V (above) and Fig.4 (below)]. Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.15, Table V (above) and Fig.4 (below)].
Table V: Development time of A. californiensis stages at 17°C ±1°C and 35 p.1000.
E: eggs; N: nauplii; C: copepodids. Mean (x) and SD.
Fig.4: Post-embryonic progress of developmental stages of A. californiensis through time at 17°C ±1°C and salinity: 35 p.1000.N: nauplii; C: copepodids. |
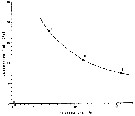 Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.17, Fig.5I]. Issued from : A. Trujillo-Ortiz in Ciencias Marinas, 1990, 16 (1). [p.17, Fig.5I].
Average generation time (days) in function of temperature of three different populations of A. californiensis: (1) Estero de Punta Banda, Baja California); (2) Yaquina bay (Oregon; (3) Elkhorn Slough (California). |
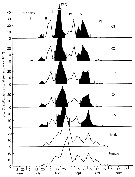 Issued from : C.B. Miller in Biological Oceanography, Blackwell Publishing, 2005/2005. [p.172, Fig. 8.12]. After J.K. Johnson, 1980. Issued from : C.B. Miller in Biological Oceanography, Blackwell Publishing, 2005/2005. [p.172, Fig. 8.12]. After J.K. Johnson, 1980.
Cohort patterns for Acartia californensis in upper Yaquina Bay (Oregon), during 1972.
Eaxh group followed through development is designated by Roman numeral.
The abundance pattern does roughly repeat in successive stages.
Establishing age-structure from the size-structure of the females and males during the time (see Brinton, 1976) |
| | | | Loc: | | | Vancouver Is., Oregon (Yaquina Bay), San Francisco Bay and Estuary, Mission Bay, California, off La Jolla, Baja California, ? Gulf of California. | | | | N: | 14 | | | | Lg.: | | | (171) F: 1,04-0,96; M: 0,96-0,87; {F: 0,96-1,04; M: 0,87-0,96} | | | | Rem.: | Salinity from Upper Newport Bay (California) = 31.3-33.1 p.1000. | | | Last update : 13/09/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 27, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




 californiensis - Plate 1 of morphological figures',750,500);)
 californiensis - Plate 2 of morphological figures',750,500);)
 californiensis - Plate 3 of morphological figures',750,500);)
 californiensis - Plate 4 of morphological figures',750,500);)
 californiensis - Plate 5 of morphological figures',750,500);)
 californiensis - Plate 6 of morphological figures',750,500);)
 californiensis - Distribution map 2',750,500);)
 californiensis - Distribution map 4',750,500);)
 californiensis - Distribution map 6',750,500);)
 californiensis - Distribution map 7',750,500);)
 californiensis - Distribution map 8',750,500);)
 californiensis - Plate 1 of morphological figures){kind=link}
 californiensis - Plate 2 of morphological figures){kind=link}
 californiensis - Plate 3 of morphological figures){kind=link}
 californiensis - Plate 4 of morphological figures){kind=link}
 californiensis - Plate 5 of morphological figures){kind=link}
 californiensis - Plate 6 of morphological figures){kind=link}
 californiensis - Distribution map 2){kind=link}
 californiensis - Distribution map 3){kind=link}
 californiensis - Distribution map 4){kind=link}
 californiensis - Distribution map 5){kind=link}
 californiensis - Distribution map 6){kind=link}
 californiensis - Distribution map 7){kind=link}
 californiensis - Distribution map 8){kind=link}
