|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Acartiidae ( Family ) |
|
|
|
Acartia ( Genus ) |
|
|
|
Odontacartia ( Sub-Genus ) |
|
|
| |
Acartia (Odontacartia) lilljeborgi Giesbrecht, 1889 (F,M) | |
| | | | | | | Syn.: | ? Acartia denticornis Brady, 1883 (Figs.F,M);
Acartia fariai Oliveira, 1945 (p.459);
Lahmeyeria turrisphari Oliveira, 1947 (p.463) | | | | Ref.: | | | Giesbrecht, 1892 (p.508, 523, figs.F,M); Giesbrecht & Schmeil, 1898 (p.155); Steuer, 1923 (p.26, figs.F,M); Carvalho, 1952 a (p.150, Rem., F,M, figs.M); Rose, 1956 (p.462); Bowman, 1965 (p.149, figs. Head: rostral filaments, Rem.); Gonzalez & Bowman, 1965 (p.255, figs.F,M, Rem.); Björnberg, 1972 (p.65, figs., Rem.N & juv.1 à 4); Björnberg & al., 1981 (p.661, figs.F,M, Rem.); Sazhina, 1985 (p.81, figs.N); Barthélémy, 1999 (p.858, 865, figs.F); 1999 a (p.9, Fig.27, E-F); Bradford-Grieve & al., 1999 (p.886, 962, figs.F,M); Pombu & Martinelli-Filho, 2012 (p.249, figs.1, 2, anomalies); Tello-Cetina & al., 2013 (p.925, genetic structure); Srinui & al., 2019, p.90, Rem. F, M); Lee S. & al., 2019 (p.85, Table 3.. | 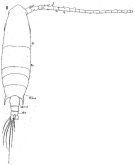 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.43, Fig.1]. Female: 1, habitus (dorsal).
|
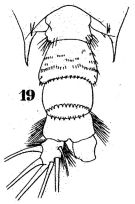 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.43, Fig.19]. Male: 19, last thoracic segment and urosome (dorsal).
|
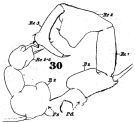 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.30, Fig.30]. Male: 30, P5 (Pd = right leg; Ps = left leg).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.30, Fig.8]. Male: 8, right A1.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.30, Fig.20]. Female: 20, P5.
|
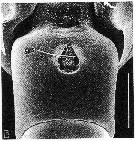 issued from : R.-M. Bathélémy in J. Mar. Biol. Ass. U.K., 1999, 79. [p.865, Fig.8, E]. Scanning electon miccrograph. Female (Punta Salinas, Ecuador): E, genital double-somite (ventral). Scale bar: 0.050 mm. Symbols: * = cuticular protuberance between the genital slits; gs = genital slit.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl. XXXII, Figs.15, 16]. With doubt as Acartia denticornis. Female: 16, P5. Male: 15, P5.
|
 Erratum: read Srinui and not Siruani. Female: 1 - Genital double-somite having paired posterodorsal processes. 2 - 2nd segment of A1 without strong curved processes. 3 - Exopod of P5 not thickened proximally. 4 - Caudal ramus longer than wide by at most ca 2 times 2nd free urosomite with small spinules dorsally and posteriorly . 5 - 5th to 7th segments of A1, each with posterior hook; genital double-somite with pair of small processes dorsally. Male: 1 - Urosomite 3 without spine-like processes dorsally. 2 - Urosomite 4 with prominence dorsally; number of dorsal prominences on urosomite 4 fewer than 5. 3 - Terminal exopodal segment of left P5 with single element. 4 - Terminal element of left P5 as fine seta.
|
 Issued from : S. Lee, H.Y. Soh & W. Lee in ZooKeys, 2019, 893. [p.84, Table 3]. Acartia (Odontacartia) lilljeborgi: Morphological characters. Compare with other Odontacartia species. Nota: 1 - Presence of spines on 1st to 2rd segments of female A1 ... 2. 2 - Small spinule row present on dorsal surface of female genital double-somite.
| | | | | Compl. Ref.: | | | Sewell, 1948 (p.392, 454, 573); Fagetti, 1962 (p.37); Gaudy, 1963 (p.29, Rem.); Björnberg, 1963 (p.61, Rem.); Fleminger, 1967 a (tabl.1); Brinton & al., 1986 (p.228, Table 1); Zamora-Sanchez & Gomez-Aguirre, 1986 (p.340); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Lindo, 1991 (tab.3); Lopes, 1994 (tab.1); Suarez Morales, 1994 b (tab.1); Palomares Garcia & Vera, 1995 (tab.1); Roff & al., 1995 (p.165,Table 5: naupllii vs. bacteriophagy/picoplankton); Eskinazi-Sant'Anna & Tundisi, 1996 (tab.1, 2); Webber & al., 1996 (tab.1); Lenz & al., 1996 (p.1, behaviour); Lopes & al., 1998 (p.195); Hopcroft & al., 1998 (tab.2); Suarez-Morales & Gasca, 1998 a. (p107); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Lopes & al., 1999 (p.215, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Gomez-Gutierrez & al., 1999 (p.2227, egg production, growth rates); Alvarez-Silva & Gomez-Aguirre, 2000 (p.163: tab.2); Ara, 2001 a (p.205, egg production); 2001 b (p.121); Dunbar & Webber, 2003 (tab.1); Krumme & Liang, 2004 (p.407, tab.1); Ara, 2004 (p.179, figs. 3, 4, 5); Alvarez-Silva & al., 2005 (p.39); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño, Rem.: p.67, 69, 70, 72, 74); Sterza & Fernades, 2006 (p.95, Table 1, occurence), Rem.: p.103; Dias & Araujo, 2006 (p.27, Rem., chart); Youn & Choi, 2007 (p.222: Table1, egg production); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.513, 517); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Waggett & Buskey, 2008 (p.111, Table 1); Miyashita & al., 2009 (p.815, Tabl.II); Magalhaes & al., 2009 (p.187, Table 1, %); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Magris & al., 2011 (p.260, abundance, interannual variability); Costa R.G. da & al;, 2011 (p.364, Table 1, seasonal occurrence); Escamilla & al., 2011 (p.379, spatial & seasonal abundance, competition); Magalhaes & al., 2011 (p.1520, seasonal abundance); Almeida LR. & al., 2012 (p.13, Table 1, abundance); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Palomares-Garcia & al., 2013 (p.1009, Table I, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, pers. comm.); Garbosa da Costa & al., 2013 (p.756, Table 1, abundance vs tide); Alvarez-Silva & Torres-Alvarado, 2013 (p.241, Table1: seasonal abundance); Mendoza Portillo, 2013 (p.37: Fig.7, seasonal dominance); Ortega & al., 2014 (p.495, Table I, %); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Fuentes-Reinés & Suarez-Morales, 2015 (p.369, Table 1, Rem. p.372); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Araujo & al., 2016 (p.1, Table 3, 5, abundance, %); Ortega & al., 2017 (p.123, fig. 1, 4, 6, Table 2, abundance, %); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Atique & al., 2017 (p.1, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); Palomares-Garcia & al., 2018 (p.178, fig.3: relative frequency, Table 1) | | | | NZ: | 5 | | |
|
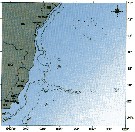
Distribution map of Acartia (Odontacartia) lilljeborgi by geographical zones
|
| | |  able 1Chart of 1996 able 1Chart of 1996 | |
 issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.27]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.27].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S).
Nota: sampling only 2 specimens. |
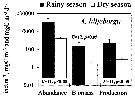 Issued from : A. Magalhaes, D.S.B. Nobre, R.S.C. Bessa, L.C.C. Pereira & R.M. da Costa in J. Coastal Res., 2011, SI 64. [p.1522, Fig.2]. Issued from : A. Magalhaes, D.S.B. Nobre, R.S.C. Bessa, L.C.C. Pereira & R.M. da Costa in J. Coastal Res., 2011, SI 64. [p.1522, Fig.2].
Seasonal variation in the abundance, biomass and productivity of Acartia lilljeborgii in the Taperaçu estuary (00°55'06.8''S, 46°44'00''W) in July (rainy season) and October (dry season), 2004, during spring tides (flood and ebb periods), at 3-hour intervals over 24-hour period. with a plankton net (300 µm mesh aperture). |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5].
Summary observations on naupliar bacterovory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Kingston Harbour and Lime Cay (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB. |
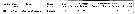 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.172, Table 6]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.172, Table 6].
Summary observations on adult female Acartia liljeborgi bacterivory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Kingston Harbour (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB. |
| | | | Loc: | | | Brazil (S, Caraguatatuba Bay, Paranagua Bay, Cananeia Lagoon, Campos Basin, Cabo Frio, Vitorio Bay, Cabo de Sao Tomé, Mucuri estuary, Guarairas lagoon, estuary do Pina, off Natal, Tocantins mouth, Curuça estuary, Ajuruteua Beach, Taperaçu estuary, Caeté Estuary, Amazon estuary, off Amazon River's mouth), Caribbean Colombia, Caribbean Sea (Chetumal Bay, Santa Lucia Is.), Yucatan (Ascension Bay, coast, Chelem lagoon), Porto-Rico, Jamaica (Kingston Harbour, Lime Cay), Venezuela, Colombia (Laguna Navio Quebrado, estuarine area) E Costa Rica, G. of Mexico, Cuba, Viet-Nam (Cauda Bay), S Pacif., Baja California (Bahia Magdalena, W), G. of California, La Paz, Coyuca lagoon, Zihuatanejo Bay, W Costa Rica, Bahia Cupica (Colombia), Ecuador (Guayaquil), Peru, Chile (Valparaiso) | | | | N: | 78 | | | | Lg.: | | | (46) F: 1,4-1,33; M: 1,1; (237) F: 1,4; M: 1,15-1,20; {F: 1,33-1,40; M: 1,10-1,20} | | | | Rem.: | In estuaries, mangroves.
The form, pointed out in Vietnam by Rose (1956) makes confirmation necessary.
The distribution of this form is different from the species of the sub-genus that are all W Indo-Pacific.
After Björnberg (1963, p.62) this species is indicator of coastal waters.
Bowman (1965, p.149 and further) notes that the specimens female and male from both localities in Santa Lucia Island (Marigot Bay) differ all the others in ne respect, the rostral filaments are completely absent; all other specimens both Atlantic and Pacific, have well developed rostral filaments. The arostrate condition of the St. Lucia population in marigot Bay is probably genetically rather than environmentally determined, because the population has been isolated from adjacent rostrate populations long enough for the arostrate condition to become completely established.
Cf Calanus pacificus (figs 3, 4 and Table 1, from Lopez-Ibarra & Palomares-Garcia). | | | Last update : 07/06/2021 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 05, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




 lilljeborgi - Plate 1 of morphological figures',750,500);)
 lilljeborgi - Plate 2 of morphological figures',750,500);)
 lilljeborgi - Plate 3 of morphological figures',750,500);)
 lilljeborgi - Plate 5 of morphological figures',750,500);)
 lilljeborgi - Plate 6 of morphological figures',750,500);)
 lilljeborgi - Plate 9 of morphological figures',750,500);)
 lilljeborgi - Distribution map 2',650,500);)
 lilljeborgi - Distribution map 3',750,500);)
 lilljeborgi - Distribution map 4',750,500);)
 lilljeborgi - Plate 1 of morphological figures){kind=link}
 lilljeborgi - Plate 2 of morphological figures){kind=link}
 lilljeborgi - Plate 3 of morphological figures){kind=link}
 lilljeborgi - Plate 4 of morphological figures){kind=link}
 lilljeborgi - Plate 5 of morphological figures){kind=link}
 lilljeborgi - Plate 6 of morphological figures){kind=link}
 lilljeborgi - Plate 7 of morphological figures){kind=link}
 lilljeborgi - Plate 8 of morphological figures){kind=link}
 lilljeborgi - Plate 9 of morphological figures){kind=link}
 lilljeborgi - Distribution map 2){kind=link}
 lilljeborgi - Distribution map 3){kind=link}
 lilljeborgi - Distribution map 4){kind=link}
 lilljeborgi - Distribution map 5){kind=link}
 lilljeborgi - Distribution map 6){kind=link}
