|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Centropagidae ( Family ) |
|
|
|
Centropages ( Genus ) |
|
|
| |
Centropages abdominalis Sato, 1913 (F,M) | |
| | | | | | | Syn.: | Centropages mcmurrichi Willey, 1920 (p.10), Rem.; Campbell, 1929 (p.314, Rem.); Sewell, 1948 (p.391, 496); Davis, 1949 (p.54, Rem.F,M); Brodsky, 1950 (1967) (p.316, figs.F,M, Rem.); Chen & Zhang, 1965 (p.72, figs.F,M); Cross & Small, 1967 (p.60, hydrologic indicator); Vinogradov, 1968 (1970) (p.51); Chen Q-c, 1980 (p.794); Zheng Zhong & al., 1984 (1989) (p.243); Shih & Marhue, 1991 (tab.3); Shih & Young, 1995 (p.69); Wang R. & al., 1998 (p.265); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Sun & al., 2010 (p.1006, Table 2); Huo & al., 2012 (p.1, Table 1: dominance) | | | | Ref.: | | | Mori, 1937 (1964) (p.58, figs.F,M); Chiba, 1956 (p.58, figs.F, Rem.); Shen & Bai, 1956 (p.220, figs.F,M); Johnson, 1961 (p.317, Rem.); Kos, 1963 (p.229, figs.F,M); Tanaka, 1963 (p.11, Rem.F,M); Vervoort, 1964 b (p.309); Koga, 1968 (p.17, fig.: egg); Kasahara & al., 1974 (p.170, fig. O); Fleminger, 1975 (p.397); Kos, 1976 (Vol. II, figs.F,M, Rem.); Sullivan & al., 1975 (p.176, figs.Md); Gardner & Szabo, 1982 (p.352, figs.F,M); Sazhina, 1985 (p.57, figs.Nauplius); Kos, 1984 (1985) (p.229, figs.F,M, Rem.); Hirakawa, 1986 (p.296, figs.F,M, Rem.); Ohtsuka & al., 1996 b (p.153, figs.F: mouth parts, gut contents); Chihara & Murano, 1997 (p.765, Pl. 82, 85: F,M); Ferrari & Ueda, 2005 (p.334, figs. juv, F, M); Ferrari & Dahms, 2007 (p.100, Rem.); Prusova & al., 2019 (p.525, Table 3). | 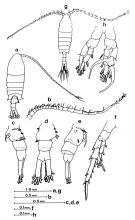 Issued from : Hirakawa K. in Crustaceana, 1986, 51 (3). [Fig.1, p.297]. Female (from 45°27'S, 72°49'W): a, habitus (dorsal view); c, urosome (lateral view, left side); d, idem (dorsal view); e, idem (lateral view, right side); f, P5. Male: b, right A1; g, habitus (dorsal view); h, P5.
|
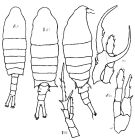
 issued from : Mori T. in The Pelagic Copepoda from the neighbouring waters of Japan. S. Shirai ed., Tokyo, 1937; 2nd edit. 1964. [Plate 28, Figs.1-6]. Female: 1, habitus (dorsal view); 2, P5; 3, genital complex (ventral view). Male: 4, habitus (dorsal view); 5, P4; 6, P5.
|
 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.24, 9-13]. As Centropages mcmurrichi. Female (from E China Sea): 9, habitus (dorsal); 10, urosome (lateral right side); 11, P5 (posterior). Male: 12, habitus (dorsal); 13, P5 (posterior).
|
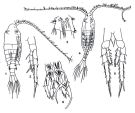
 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.316, Fig.219]. As Centropoages mcmurrichi. Female (from Sea of Japan): habitus (dorsal and lateral right side); S5, P5. Male: habitus (dorsal); S5, P5 (Le = left leg; Ri, right leg).
|
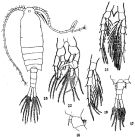 issued from : C.-j. Shen & S.-o. Bai in Acta Zool. sin., 1956, 8 (2). [Pl.III, Figs.17-22]. Female (from Chefoo): 17,urosome (dorsal); 18, genital segment (lateral left side); 19, P5. Nota: Genital segment more convex on the right side, bearing fine setae on both lateral borders, and in front of the genital pore, there is a posteriorly pointed spine Male: 20,habitus (dorsal); 21, P4; 22, P5. Nota: The right leg of P4 is longer than the left.
|
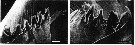 issued from : B.K. Sullivan, C.B. Miller, W.T. Peterson & A.H. Soeldner in Mar. Biol., 1975, 30. [p.180, Fig.5, C-D]. Centropages abdominalis (from 44°40'N, 124°10'W) female: C, SEM of right Md (posterior surface); D, detail od dorsal end.
|
 Issued from : R. Hirota & S. Uno in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.80, Fig.3, A]. Pelagic eggs of Centropages abdominalis from vicinity of Amakusa-Matsushima (western Kyushu, Japan).
|
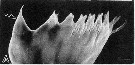 Issued from : S. Ohtsuka, M. Shimozu, A. Tanimura, M. Fukuchi, H. Hattori, H. Sasaki & O. Matsuda in Proc. NIPR Symp. Polar Biol., 1996, 9. [p.158, Fig.4, A]. Centropages abdominalis Female (from Bering Sea, Chukchi Sea): Mandibular cutting edge, ventralmost tooth (arrowed). Scale bar = 10 µm.
|
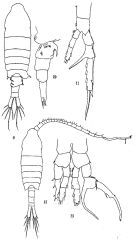
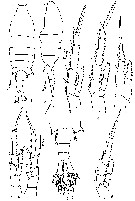 Issued from : M.S. Kos in Issled. Fauny Morei, 1984 (1985), 30 (38). [p.230, Fig.1]. Female (from South Sakhalin Islands): 1-2, habitus (dorsal and lateral, respectively); 3, P3; 4, P4; 5, P1; 6, P5; 7, abdomen; 8, P2.
|
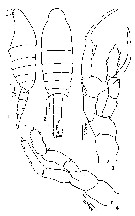 Issued from : M.S. Kos in Issled. Fauny Morei, 1984 (1985), 30 (38). [p.231, Fig.2]. Male (from South Sakhalin Islands): 1-2, habitus (lateral and dorsal, respecticely); 3, Right P5; 4, Left P5.
|
 Issued from : I.Yu Prusova, E.A. Galagovets & E.V. Popova in Arthropoda Selecta, 2019, 28 (4). [p.525, Table 3]. Centropages abdominalis: Characters for distinguishing between species of the hamatus group of the genus Centropages. Compare with the species C. hamatus, C. kroyeri, C. ponticus, and C. tenuiremis.
| | | | | Compl. Ref.: | | | Yamazi, 1958 (p.149, Rem.); Minoda, 1958 (p.253, Table 1, abundance); M.W. Johnson, 1961 (p.311, Table 2); Northcote & al., 1964 (p.1069, Table II); Motoda, 1966 (p.260); Shih & al., 1971 (p.146, 203); Peterson & Miller, 1975 (p.642, 650, Table 3, interannual abundance); Kasahara & al., 1975 (p.25, eggs, cycles); Peterson & Miller, 1977 (p.717, Table 1, seasonal occurrence); Hirota & Uno, 1977 (p.77, fig. egg, seasonal abundance); Hanaoka, 1977 (p.267, 300, abundance); Gibson & Grice, 1977 (p.85, Table 1, copper pollution); Peterso & al., 1979 (p.467, Table 1); Grice & Marcus, 1981 (p.125, Dormant eggs, Rem.: p.133, 135, 137); Hirota 1981 (p.19, Table 1, length-weight-CHN); Uye S-i., 1982 (p.149, relation length-weight-C-N); Mackas & Sefton, 1982 (p.1173, Table 1); Yamamoto & Nishizawa, 1986 (p.1729, horizontal distribution); Hirakawa, 1986 (p.296, Rem.); Mackas & Anderson, 1986 (p.115, Table 2); Nagasawa, 1987 (p.101, Zoothamnium attached); Springer & al., 1989 (p.359, Fig.5), Coyle & al., 1990 (p.763); Hirakawa & al., 1990 (tab.3); Shih & Marhue, 1991 (tab.3); Hattori, 1991 (tab.1, Appendix); Yoo, 1991 (tab.1); Kimmerer, 1993 (tab.2); Hwang & Choi, 1993 (tab.3); Myung & al., 1994 (tab.1); Liang & al., 1994 (p.131, egg production); Uye & Kaname, 1994 (p.43, length v.s. fecal pellet volume); Shih & Young, 1995 (p.69); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Kotani & al., 1996 (tab.2); Liang & al., 1996 (p.527, population dynamics, production); Marcus, 1996 (p.143); Park & Choi, 1997 (Appendix); Liang & Uye, 1997 (p.415, population dynamic, production); Mauchline, 1998 (p.506, tab.25, 33, 35, 36, 40, 45, 47, 48, 58, 78); Suarez-Morales & Gasca, 1998 a (p.109); Gomez-Gutiérrez & Peterson, 1999 (p.637, Table I, II, III, VI, figs.4, 5, 6, 7, abundance, egg production); Bragina, 1999 (p.196); Pinchuk & Paul, 2000 (p.4, table 1, % occurrence); ; Peterson & al., 2002 (p.353); Peterson & al. 2002 (p.381, Table 2, interannual abundance); Peterson & Keister, 2003 (p.2499, interannual variability); Keister & Peterson, 2003 (p.341, Table 1, 2, abundance, cluster species vs hydrological events); Shimode & Shirayama, 2004 (tab.2); Park, W & al., 2004 (p.464, tab.1); Kasyan, 2004 (p.105); Hooff & Peterson 2006 (p.2610); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño); Deibel & Daly; 2007 (p.271, Table 4, Rem.: Arctic polynyas); Gaudy & Thibault-Botha, 2007 (p.84, Tab.2, Rem.: metabolism); Kang J.-H. & al., 2007 (p.82, fig.3, temporal variation vs. temperature & predation effect); Humphrey, 2008 (p.83: Appendix A); Liu & Hopcroft, 2008 (p.931: Table IV); Ohtsuka & al., 2008 (p.115, Table 5); Coyle & al., 2008 (p.1775, Table 5, abundance): Peterson, 2009 (p.73, Rem.: p. 78, indicator of long-term changes); Hopcroft & al., 2009 (p.9, Table 3, Fig.21, 24); Galbraith, 2009 (pers. comm.); Hopcroft & al., 2010 (p.27, Table 1, 2, fig.9); Bucklin & al., 2010 (p.40, Table 1, molecular biology); Keister & al., 2011 (p.2498, interannual variation); Batten & Walne, 2011 (p.1643, Table I, abundance vs temperature interannual variability); Matsuno & al., 2011 (p.1349, Table 1, abundance vs years); Beltrao & al., 2011 (p.47, Table 1, density vs time, as C. abdominales); Matsuno & al., 2012 (Table 2); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Volkov, 2012 (p.474, Table 6, fig. 6, 12, abundance, distribution, interannual variation); Lavaniegos & al., 2012 (p. 11, Appendix); Lee J. & al., 2012 (p.95, Table 2: abundance, annual & seasonal distribution); Lee D.B. & al., 2012 (p.88, co-occurrence); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Moon & al., 2012 (p.1, Table 1); Takahashi M. & al., 2012 (p.393, Table 2, water type index); Li C. & al., 2013 (p.101, feeding); Ohashi & al., 2013 (p.44, Table 1, Rem.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Questel & al., 2013 (p.23, Table 3, interannual abundance & biomass, 2008-2010); Suzuki, K.W. & al., 2013 (p.15, Table 2, 3, 4, estuaries, annual occurrence); Oh H-J. & al., 2013 (p.192, Table 1, occurrence); Eisner & al., 2013 (p.87, Table 3: abundance vs water structure); Barton & al., 2013 (p.522, Table 1: metabolism, diapause, population dynamic, feeding mode, biogeo); Coyle & al., 2014 (p.97, table 3, 10, 12); Chiba S. & al., 2015 (p.968, Table 1: length vs climate); Zakaria & al., 2016 (p.1, Table 1, 3, Rem.); Ohtsuka & Nishida, 2017 (p.565, Table 22.1, Fig.22.3: central station in Japanese waters). | | | | NZ: | 9 | | |
|
Distribution map of Centropages abdominalis by geographical zones
|
| | | | | | | | |  Chart of 1996 Chart of 1996 | |
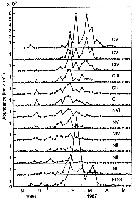 issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.529, Fig.4]. issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.529, Fig.4].
Centropages abdominalis (from Fukuyama Harbor, Inland Sea, Japan). Seasonal variation in nauplii stage-specific abundance.
Nota: Sampling in a temperate eutrophic inlet. This area receives nutrient-rich water from a near by sewage plant, which provides secondary treatment to waste water. Red-tides and anoxic conditions at the bottom often occur in summer.
Samplings at intervals of 3 to 5 days from November 1986 to 8 November 1987. Zooplankton taken by oblique hauls by plankton net (62 µm mesh aperture) from bottom (depth 7 to 8 m) to the surface.
C. abdominalis were staged, sexed and counted, but as a congeneric species C. tenuiremis was also present, the separation between these species was made following the morphological descriptions of their developmental stages by Koga (1960) and Li & Fan (1990). |
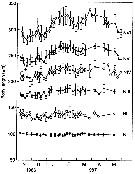 issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.530, Fig.5]. issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.530, Fig.5].
Centropages abdominalis: Seasonal variation in body size of each developmental stage from copepodites I to adults CVI females and males.
Error bars denote SD. |
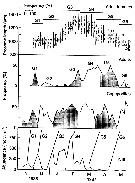 issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.531, Fig.6]. issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.531, Fig.6].
Centropages abdominalis: Generation analysis based on numerical abundance of NIII, relative abundance of copepodites and adults in total abundance, and porosome length of adults females.
Nota: The generation G1 is derived from resting eggs in the bottom sediment; then it is impossible to determine a survival curve
Six generations appears during the year.
A survival curve for each generation could be generated by using the mean egg production rate and the mean daily abundance of each developmental stage of a given generation. |
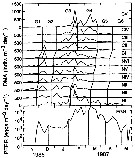 issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.532, Fig.9]. issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.532, Fig.9].
Centropages abdominalis: Seasonal variation in population egg production rate (PEPR, in logarithmic scale) and daily midsstage abundance (DMA) of each stage.
Each generation is delimited by vertical lines. |
 Issued from : A.F. Volkov in Russian Journal of Marine Biology, 2012, 38 (7). [p.483, Fig.6] Issued from : A.F. Volkov in Russian Journal of Marine Biology, 2012, 38 (7). [p.483, Fig.6]
Dynamics in the biomass of Centropages abdominalis in the eastern Bering Sea from 2003-2011 years. |
 Issued from : A.F. Volkov in Russian Journal of Marine Biology, 2012, 38 (7). [p.491, Fig.12] Issued from : A.F. Volkov in Russian Journal of Marine Biology, 2012, 38 (7). [p.491, Fig.12]
Distribution of Centropages abdominalis in the eastern Bering Sea in the fall. Biomass in mg/m3. |
 Issued from : H. Bi, K.A. Rose & M.C. Benfield in Mar. Ecol. Prog. Ser., 2011, 427. [p.157, Table 5]. Issued from : H. Bi, K.A. Rose & M.C. Benfield in Mar. Ecol. Prog. Ser., 2011, 427. [p.157, Table 5].
Instantaneous mortality rates (individuals per day) from litterature. |
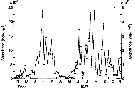 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.529, Fig.3] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.529, Fig.3]
Seasonal variations in total (nauplii + copepodites + adults abundances of Centropages abdominalis (filled circles) and Centropages tenuiremis (open circles) in Fukuyama Harbor (Inland Sea of Japan) from 7 November 1986 to 8 November 1987.
Within the thermal range between the two species (See remarks in Fig.14), the density and age structure of the C. abdominalis population are explained by temporal variations in the balance between fecundity and mortality.
The fecundity varied from 39 to 142 eggs/female/day, and the weight-specific egg production rate was positively correlated to both temperature and chlorophyll a concentration.
The population egg production rate depended largely of the density of adult females, and it was highest in February and March (see in Fig.9).
The fraction surviving from egg to Nauplius II was extremely low and variable (see in Fig. 10). In Generation G2, although the population egg production rate was lowest(, the fraction surviving to NII was highest (i.e. 4.0%) In G5, it was vice versa, and only 0.02% of the eggs spawned survived to NII. Landry (1978) reported that about 80% of the eggs never survived to NII for Acartia clausi in a temperate lagoon in Washington State (USA), and Uye (1982) found that mortality of 80 to 90% occurred from egg to NIII for Acartia omorii in Onagawa Bay (Japan).
Liang & al. (1994) assessed the loss of eggs by comparing incubation fecundity and egg-ratio fecundity of C. abdomionalis: 37 to 99% (mean: 87.5%) of the spawned eggs were lost during egg stage within the water column.
Hence, mortality in early life is probably concentrated in the egg stage. A significant part of the loss might be attributed to predation, including cannibalism, in addition to loss by sinking (see in Liang & al., 1994).True carnivores such fish larvae, chetognaths, medusae and ctenophores were absent or extremely rare in the Fukuyama's site during February-May while adults and late copepodites (CIV-CV) of C. abdominalis and Acartia omorii (both omnivorous) occurred (as many as 39 000 ind/m3) during this period. Assuming the clearance rate of each individual is 50 ml/ind./day), 1 to 86% of the water column was estimated to be cleared by day, thus the main source of mortality (by cannibalism) impacts the young stages (see in Hada & Uye, 1991 for Sinocalanus tenellus in a brackish-water pond of Fukuyama). Several species of diatoms inhibited copepod embryonic development causing higher egg mortality. The cell diameter of diatoms may play also on the possibility of capture by nauplii (See in Ianora & Poulet, 1993 and Poulet & al., 1994) |
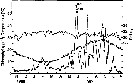 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.529, Fig.2] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.529, Fig.2]
Seasonal variations in temperature (open circles), salinity (filled triangles) and chlorophyll a concentration (filled circles) in Fukuyama Harbor from November 1986 to November 1987. |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.534, Fig.14] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.534, Fig.14]
Relationship between abundance and temperature for Centropages abdominalis (filled circles) and Centropages tenuiremis (open circles).
Nota: A clear pattern of alternation in the site was evident for C. abdominalis and C. tenuiremis. The abundance of both species and temperature were linked.: C. abdominalis appeared in the site when temperature was below 21.1 °C, and its density was higher at temperature below 10°C. On the contrary C. ternuiremis appeared when temperature was higher than 14.5°C, and became abundant at temperatures above 21°C. Replacement seems to occur within a temperature range between 14.5 and 21.1°C.
During an unfavorable temperature period, either species can exist as resting eggs on the sea-bottom (see Kasahara & al., 1974, 1975).
Temperature is the prime determinant of their occurrence in the Fukuyama's plankton. |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.532, Table 2] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.532, Table 2]
Centropages abdominalis (from Fukuyama Harbor, Japan). Conparison between generation times in the field and development times from egg-laying to adult predicted from laboratory experiments. |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.532, Fig.10] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.532, Fig.10]
Centropages abdominalis (from Fukuyama Harbor, Japan). Survival curves for generations G2 to G6. |
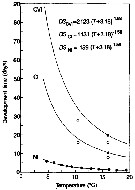 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.531, Fig.8] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.531, Fig.8]
Centropages abdominalis (from Fukuyama Harbor, Japan). Relationship between water temperature and development time from egg-laying to hatching, to CI and adult.
Continuous lines calculated from egg development times using equations '6) and (7). Open and filled circles denote minimum development time and median development time, respectively.
Equation 6 expressed by the Belehradek's function: DSnI = 159 (T + 3.18) exp. -1.38, assuming that the species follows the ''equiproportional'' rule of development (Corkett, 1984) as observed for Centropages typicus (Fryd & al., 1991), the time required from egg-layying to reach developmental Stage i (DSi, d) can be calculated from the embryonic duration by multiplication of a proportional constant (M) (Corkett & McLaren, 1970) i.e equation 7: DSi = M x DSnI.
The multipliers for successive stages were: 1.68 (NII), 3.10 (NIII), 4.10 (NIV), 5.00 (NV), 5.95 (NVI, 78.10 (CI), 8.40 (CII), 9.68 (CIII), 10.9 (CIV), 12.2 (CV), 13.4 (adult).
Thus, the times from egg-laying to CI and adult were expressed by:
DSCI = 1131 (T + 3.18)-1.58
and
DSCVI = 2123 (T + 3.18)-1.58 . |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.535, Table 3] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.535, Table 3]
Comparison of development time from egg-laying to adult (D) and instantaneous growth rate (g) at various temperatures within the genus Centropages. (- no data). |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.530, Table 1] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.530, Table 1]
Centropages abdominalis (from Fukuyama Harbor, Japan). Regression equations describing the relationship between body size and water temperature (T, °C). Body length (BL, µm) for nauplii and prosome length (PL, µm) for copepodites and adults.
r : correlation coefficient.
Expression of relations by Belehradek's functions, non linear least-squares were applied. |
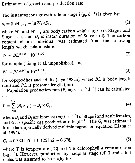 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.538] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.538]
Centropages abdominalis (from Fukuyama Harbor, Japan). Equations used for estimation of growth (g) and production (P) rate. |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.530, Table 1] Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.530, Table 1]
Centropages abdominalis (from Fukuyama Harbor, Japan). Postembryonic development in the laboratory assuming that this species follows the ''equiproportional'' rule of development, as observed for Centropages typicus. |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.535, Table 3 & 4]. Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.535, Table 3 & 4].
Table 3 : Comparison of development time from egg-laying to adult (D) and instantaneous growth rate (g) at various temperatures within the genus Centropages. (- no data).
Table 4 : Comparison of the instantaneous growth rate (g) at 10¨C among calanoid copepods occurring in the Inland Sea of Japan.
Nota: The development time among three Centropeges species (Table 3) were almost the same rate; at temperatures around 17°C it takes about 20 days for them to develop from egg to adult, although the development time of C. typicus is longer than taht other species (Smith & Lane (1987)
The instantaneous growth rate of C. abdominalis is similar toi that of the other Centropages species.
Fryd & al. (1991) found thet the growth rate is constant over the all developmental stages for C. hamatus and C. typicus. For C. abdominalis, however, the growth rate is higher in copepodites than in nauplii. Moreover, the growth rate of C. abdominalis is high compared to those of the other calanoid species occuring in the low temperature period in the Inland Sea (Table 4) |
 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.533, Fig.11]. Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.533, Fig.11].
Centropages abdominalis (from Fukuyama Harbor, Japan). Relationship between mean instantaneous growth rate and water temperature. |
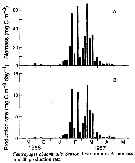 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.533, Fig.12]. Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.533, Fig.12].
Centropages abdominalis (from Fukuyama Harbor, Japan). Seasonal variation in A biomass and B production rate.
Nota: The biomass was less than 5 mgC/m3 until late January and became high in February and March.
The maximum biomass of 67.1 mgC/m3 was recorded on 11 March 1987.
The pattern of the seasonal variation in production rate was similar to that biomass.
The maximum production rate (12.4 mg C/m3/day) was also recorded on 11 Marcgh 1987.
Cumulative production between 7 November 1986 and 29 May 1987 was 355 mg C/m3 or 2.66 g C/m2.
The contribution of nauplii to the population production was minor (overall mean: 8.6%), and primarily owed to copepodites (57.8%) and adult females (33.6%) |
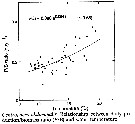 Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.533, Fig.13]. Issued from : D. Liang, S. Uye & T. Onbé in Mar. Biol., 1996, 124. [p.533, Fig.13].
Centropages abdominalis (from Fukuyama Harbor, Japan). Relationship between daily production/biomass ratio (P/B) and water temperature.
Nota:
The empirical relationship between the ratio of daily production rate to biomass (P/B) and temperature denotes that the ratio increased exponentially with increasing temperature.
The daily production rate to biomass ratio increased exponentially with temperature from 0.18 at 8.9°C to 0.37 at 19°C. |
| | | | Loc: | | | Hong Kong, China Seas (Bohai Sea, Yellow Sea, East China Sea, South China Sea), W & S Korea, Chunsu Bay, Asan Bay, Muan Bay, Geumo Is., Korea Strait, Japan(Akkeshi Bay, Honshu), Inland Sea, Bikini Is., Japan (Ariake Bay, Seto Inland Sea, Tokyo Bay, Tanabe Bay, south estuaries), off Sanriku, Sea of Japan (Amursky Bay), Okhotsk Sea, S. Sakhalin, Chukchi Sea, off NW Alaska, N & E Bering Sea, Karagin Bay, St. Lawrence Island, Anadyr Strait, Alaska, Icy Strait, Hudson Bay, Queen Charlotte Is., British Columbia, Fjord system (Alice Arm & Hastings Arm) , Portland Inlet, Georgia Strait, Vancouver Is., Nitinat Lake, Washinton coast, Oregon (coast, Yaquina, off Newport), San Francisco Bay, E Pacif. (off Mexico), Baja California (rare in Bahia Magdalena), G. of California (rare in Bahia de los Angeles, after Lavaniegos & al., 2012), S Chile, E Medit. (W Egyptian coast in Zakaria & al. (2016) | | | | N: | 101 (Arct.: 11; N Pacif.: 90; Medit.: 1) | | | | Lg.: | | | (22) F: 2,1-1,63; M: 1,46; (26) F: 1,61; M: 1,52-1,43; (59) F: 1,9-1,7; (280) F: 2,1-1,48; (290) F: 1,9-1,7; M: 1,6-1,35; (866) F: 1,3-2,1; M: 1,2-1,6; {F: 1,30-2,10; M: 1,20-1,60}
Chiba S. & al., 2015 (p.971, Table 1: Total length female (June-July) = 1.4 mm [optimal SST (°C) = 11.1]. | | | | Rem.: | coastal ± brackish.
For Willey (1920, p.10-11) McMurrich off the entrance to Esparanza inlet (west coast of Vancouver Island) identified Centopages hamatus, whilst noting differences. The female has the ventral recurved hook of C. hamatus in front of the genital pore but the remaining setulose armature of the genital segment is distinctive. The other principal character is afforded by the strong unguiform process on the inner side of the exopod 2 of P5; in C. hamatus this process is smooth and less than half the length of the exppod 3; in the prsent species the process is at least two-thirds the length of exopod 3 and is denticulated along its outer edge. The relative dimensions of anal segment and caudal rami in the female are: anal segment 6, caudal ramus length 11, width of caudal ramus 3. The spinules on the genital segment of the female include a pair of anteroventral groups arranged in a comb-like row right and left of the swollen base of the hook; a right lateral comb-like group of strong spinules near the middle of the segment and a left postero-lateral group of much smaller spinules near the posterior end of the segment; also right and left dorso-lateral tufts in the anterior region; the segment itself and the groups of spinules are asymmetrical.
Hirakawa (1986) reports the presence of this species near southern Chile (Ensenada Baja, a lagoon at the head of Aysén Fjord) as the result of transport by tanker from Japan (Cf. in Carlton, 1987, fig.1: map).
Madhupratap & al. (1996, Table 2) point to the occurrence of the resting eggs in this species in The Inland Sea of Japan by Kasahara & al. (1974).
After Barton & al. (2013, Table 1) the feeding mode is a mixing of 'ambush' and 'cruising' behaviour (see definition in Centropages typicus). | | | Last update : 11/01/2021 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed January 02, 2026] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
