|
|
 |
|
Calanoida ( Order ) |
|
|
|
Metridinidae ( Family ) |
|
|
|
Pleuromamma ( Genus ) |
|
|
| |
Pleuromamma borealis (F. Dahl, 1893) (F,M) | |
| | | | | | | Syn.: | Pleuromma borealis F. Dahl,1893;
Pleuromamma gracilis : Esterly, 1905 (p.175, fig.F, Rem.F,M); 1912; Farran, 1926 (part.);
Pleuromamma gracilis esterlyi T. Scott, 1912 (1913) (p.535, figs.F) | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.110); Farran, 1929 (p.209, 262, figs.F, Rem.); Steuer, 1932 a (p.39, 52, 67, 78, figs.F,M); Rose, 1933 a (p.182, figs.F); Massuti Alzamora, 1942 (p.91, figs.F); Farran, 1948 f (n°17, p.3, figs.F,M); Brodsky, 1950 (1967) (p.309, figs.F,M); Grice, 1962 (p.218, figs.F); Giron-Reguer, 1963 (p.33, figs.F); Paiva, 1963 (p.56); Chen & Zhang, 1965 (p.70, figs.F,M); Vervoort, 1965 (p.111, Rem.); Vilela, 1968 (p.24, figs.F,M); Corral Estrada, 1970 (p.176); Arcos, 1975 (p.18, figs.F,M); Dawson & Knatz, 1980 (p.5, figs.F,M); Björnberg & al., 1981 (p.642, figs.F); Gardner & Szabo, 1982 (p.336, figs.F,M); Zheng & al., 1982 (p.52, figs.F); Ferrari, 1984 a (p.167, figs.F); Kim & al., 1993 (p.270); J.S. Park & Mauchline, 1994 (p.108 & suiv., fig.F); Bradford-Grieve & al., 1999 (p.884, 949, figs.F,M); Bradford-Grieve,1999 b (p.120, figs.F,M, Rem., figs.179, 192); G. Harding, 2004 (p.31, figs.F,M); Vives & Shmeleva, 2007 (p.377, figs.F,M, Rem.); Cheng F. & al., 2013 (p.119, molecular biology, GenBank) | 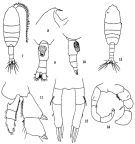 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.23, 7-14]. Female (from E China Sea): 7, habitus (dorsal); 8, forehead (lateral); 9, urosome (ventral); 10, idem (lateral right side); 11, endopod of right P2 (posterior); 12, P5 (posterior). Male: 13, habitus (dorsal); 14, P5 (posterior).
|
 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.48, fig.1]. Female (from Canarias Is.): 1, P5.
|
 issued from : F. Giron-Reguer in Thèse Fac. Sc. Paris, 1963 [Fig.13]. Female (from Alboran Sea): P5.
|
 issued from : F. Giron-Reguer in Thèse Fac. Sc. Paris, 1963 [Fig.12]. Female: thoracic segments and urosome (ventral view).
|
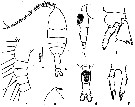 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.52, Fig.29]. Female: a, habitus (dorsal); b, forehead (lateral); c-d, urosome (lateral and ventral, respectively); e, proximal segments of A1; f, basipodite segment 2, exopodal segment 1 and endopod of P2; g, P5. Scale bar in mm.
|
 issued from : C.O. Esterly in Univ. Calif. Publs. Zool., 1905, 2 (4). [p.175, Fig.33, c]. As Pleuromamma gracilis. Female (from San Diego Region): c, P5.
|
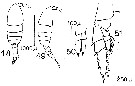 issued from : D.F.R. Arcos in Gayana, Zool., 1975, 32. [Lam.V, Figs.48-51]. Female (from Bahia de Concepcion, Chile): 48-49, habitus (dorsal and lateral, respectively) [ Juveniles ? ]; 50, P5; 51, P2.
|
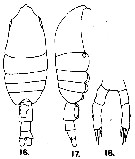 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.217, Pl.22, Figs.16-18]. Female (from equatorial Pacific): 16-17, habitus (dorsal and lateral, respectively); 18, P5.
|
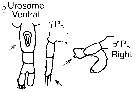 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.31]. Female & Male. Male: Right P5 with s short knobbed process (arrowed) on the exopodal segment 2.
|
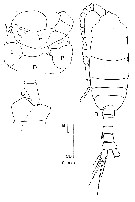 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.120, Fig.81]. Female (from 41°41S, 167°03'E): B, habitus (dorsal); C, basipod 2 and endopod segment 1 of left P2; D, P5. Nota: L = left leg; R = right leg. Female characteristics: Pigment spot on the right side - Very similar to P. gracilis- Genital boss flattened in lateral view. - P5 3-segmented with 2 free segments, terminated by 3 long spines, the outer the longest. Male characteristics: Pigment spot on right side.
- Differs from P. gracilis by exopod segment 1 of left P5 bearing a shorter appendage without a knob; distal segment of right leg shorter and wider.
Nota: The identity of the Southwest Pacific males is not always clear, therefore there is a possibility that P. gracilis, P. piseki, P. borealis have been confused when they have not been dissected.
| | | | | Compl. Ref.: | | | Massuti Alzamora, 1942 (p.91); Lysholm & al., 1945 (p.32); Sewell, 1948 (p.503, 508, 514, 547, 549); C.B. Wilson, 1950 (p.289); Grice & Hart, 1962 (p.287, table 4: abundance); Duran, 1963 (p.20, Rem.); Giron-Reguer, 1963 (p.33); Grice, 1963 a (p.496); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Björnberg, 1963 (p.51, Rem.); Unterüberbacher, 1964 (p.27); De Decker & Mombeck, 1964 (p.13); Grice & Hulsemann, 1965 (p.224); Mazza, 1966 (p.71); 1967 (p.329); Fleminger, 1967 a (tabl.1); De Decker, 1968 (p.45); Shih & al., 1971 (p.42); Timonin, 1971 (p.281, trophic group); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 b (p.549, Rem.); Björnberg, 1973 (p.338, 388); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Vives & al., 1975 (p.46, tab.II, III, IV, XII); Peterson & Miller, 1976 (p.14, Table 1, 3, abundance vs interannual variations); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Timonin & Voronina, 1977 (p.292); Tranter, 1977 (p.596, 602); Peterson & Miller, 1977 (p.717, Table 1, seasonal occurrence); Carter, 1977 (1978) (p.36); Vives, 1982 (p.293); Kovalev & Shmeleva, 1982 (p.84); Dessier, 1983 (p.89, Tableau 1, 2, Rem., %); Tremblay & Anderson, 1984 (p.5); De Decker, 1984 (p.316, 359: chart); Sameoto, 1984 (p.767, vertical migration); Roe, 1984 (p.358); Guangshan & Honglin, 1984 (p.118, tab.); Longhurst, 1985 (tab.2); Greze & al., 1985 (p.9); Brenning, 1985 a (p.28, Table 2); Wishner & Allison, 1986 (tab.2); Brenning, 1986 (p.11, spatial distribution, T-S diagram, Rem.); Saraswathy, 1986 (p.189); Madhupratap & Haridas, 1986 (p.105, tab.1); Lozano Soldevilla & al., 1988 (p.59); Dessier, 1988 (tabl.1); Timonin, 1990 (p.479); Yoo, 1991 (tab.1); Ashjian & Wishner, 1993 (p.483, abundance, species group distributions); Seguin & al., 1993 (p.23, Table 2: abundance, %), p.26, Rem.); Morales C.E. & al., 1993 (p.185); Landry & al., 1994 (p.55, abundance, grazing); Landry & al., 1994 a (p.73, grazing, gut evacuation); Hays & al., 1994 (tab.1); Genin & al., 1994 (p.941, fig.5, patchiness); Shih & Young, 1995 (p.70); Errhif & al., 1997 (p.422); Timonin, 1997 (p.83, Rem.); Gilabert & Moreno, 1998 (tab.1, 2); Suarez-Morales & Gasca, 1998 a (p.110); Reid & Hunt, 1998 (p.310, figs.2, 3, Rem.); Lapernat, 2000 (tabl. 3, 4); Razouls & al., 2000 (p.343, Appendix); Haury & al., 2000 (p.69, Table 1); d'Elbée, 2001 (tabl. 1); Rebstock, 2001 (tab.2, 4); 2002 (p.71, Table 3, 5, 6, Fig.2, climatic variability); Holmes, 2001 (p.20); Sameoto & al., 2002 (p.13); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, figs.5, 6); Labat & al., 2002 (p.741, tab.3); CPR, 2004 (p.59, fig.176); Lo & al., 2004 (p.89, tab.1); Hunt, 2004 (p.1, 74, Table 4.7); Zuo & al., 2006 (p.163: tab.1); Lavaniegos & Jiménez-Pérez, 2006 (p.152, tab.2, 3, Rem.); Mackas & al., 2006 (L22S07, Table 2); Khelifi-Touhami & al., 2007 (p.327, Table 1); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); C.-Y. Lee & al., 2009 (p.151, Tab.2); Galbraith, 2009 (pers. comm.); Lidvanov & al., 2010 (p.356, Table 3); Takahashi & al., 2010 (p.317, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Mazzocchi & Di Capua, 2010 (p.426); Pepin & al., 2011 (p.273, Table 2, seasonal abundance); Tutasi & al., 2011 (p.791, Table 1, 3, fig.7, abundance distribution vs La Niña event, Rem. p.796, fig.11); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Lidvanov & al., 2013 (p.290, Table 2, % composition); Zaafa & al., 2014 (p.67, Table I, occurrence); Zakaria & al., 2016 (p.1, Table 1, Rem.); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones) | | | | NZ: | 19 | | |
|
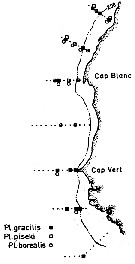
Distribution map of Pleuromamma borealis by geographical zones
|
| | | | | | | | | | | | | | | | | |  issued from : M. Saraswathy in Mahasagar-Bull. Nat. Inst. Oceanogr., 1986, 19 (3). [p.190, Fig.2A]. issued from : M. Saraswathy in Mahasagar-Bull. Nat. Inst. Oceanogr., 1986, 19 (3). [p.190, Fig.2A].
Distribution of P. borealis in the Indian Ocean (upper water column of 200 m).
Triangle: South West Moonsoon season: April 16 to October 15 (white triangle = negative stations; black = animals collected). Circle: North East Moonsoon season: October 16 to April 15 (white circle = negative stations; black = numbers of ianimals collected).
Nota: This species appears to be more or less restricted to the area off the coast of Africa, with very few scattered occurrences towards the southern regions also south of Java. |
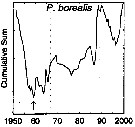 issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 t]. issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 t].
Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California (31°-35°N, 117°-122°W.
Cumulative sums of nonseasonal anomalies from the long-term means of copepod abundance from years 1950 to 2000.
A negative slope indicates a period of below-average anomalies; a positive slope indicates a period of above-average anomalies. Abrupt changes in slope indicate step changes. Step changes are marked with arrow (downward -pointing for decreases).
The October 1966 cruise (prior to the increase in sampling depth), March 1976 cruise (prior to the 1976-77 climatic regime shift), and October 1988 cruise (prior to the hypothesized 1989 climatic regime shift) are marked with vertical lines. |
 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.8, Fig.5]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.8, Fig.5].
Spatial distribution for Pleuromamma borealis, P. piseki and P. gracilis from 8° S - 26° N; 16°- 20° W. |
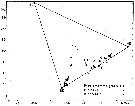 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.10, Fig.7]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.10, Fig.7].
T-S Diagram for Pleuromamma boreals, P. piseki and P. gracilis from 8° S - 26° N; 16°- 20° W.
SO: Southern Surface Water (S °/oo: 34,50; T°C: 29,0); ND: Northern Water of the Surface Layer (S °/oo: 37,5; T°C: 21,0); SD: Southern Deep Water of the surface layer (S °/oo: 35,33; T°C: 13,4). See commentary in Temora stylifera and Brenning (1985 a, p.6). |
 Issued from : A. Dessier in Oceanol. Acta, 1983, 6 (1). [p.101, Fig.8 C]. Issued from : A. Dessier in Oceanol. Acta, 1983, 6 (1). [p.101, Fig.8 C].
Geographic distribution of Pleuromamma borealis along the transect Noumea (New Caledonia) to Panama.
black square: more than 18 organisms; black point inside square: 1 to 18 organisms.
Nota: Sampling by net (mesh aperture = 330 µ) between depth 5 and 10 m, 34 hours after sunset. From November 1977 to September 1981 (90 % of samples obtained since 1979). |
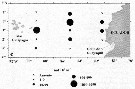 Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.798, Fig.7 c] Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.798, Fig.7 c]
Geographic distribution of Pleuromamma borealis in September and October 2001, associated with the weak La Niña event of 2001. |
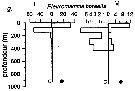 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.9 g]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.9 g].
Verical distribution of Pleuromamma borealis at an eutrophic site (off Mauritanian coast: 20°32 'N, 18°36' W) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
| | | | Loc: | | | sub-Antarct. (SW & SE Pacif., Indian), South Africa (E & W), Namibia, Congo, G. of Guinea, Brazil, off Amazon, Cape Verde Is., off Mauritania-NW Cape Verde Is., off Morocco-Mauritania, Canary Is., off Madeira, Lisboa, off W Cabo Finisterre, Bay of Biscay, NW Atlant., W Ireland, off SW Ireland, Ibero-moroccan Bay, off Bermuda: Station S (32°10N, 64°30W), off Cape Hatteras, Sargasso Sea, Nova Scotia, off SE Nova Scotia, Labrador (coast), off E Newfoundland, S Iceland, Ireland (S, W & NE), Faroe Is., North Sea, off W Tangier, Medit. (Alboran Sea, Gulf of Annaba, El Kala shelf, Baleares, Ligurian Sea, Tyrrhenian Sea, Strait of Messina, W Egyptian coast), Natal, Indian, S Indian (subtropical convergence), W Australia, Philippines, China Seas (Yellow Sea, East China Sea, South China Sea), Taiwan (N: Mienhua Canyon, NW), S Korea, Pacif. (E & W equatorial), New Caledonia, Tasman Sea, New Zealand (W, S: sub-Antarctic), Oregon (off Newport), California, Santa Monica Bsin, Sixtymile Bank, W BaJa California, off W Guatemala, Pacif. (equatorial), W Colombia, Galapagos-Ecuador, Peru, N Chile | | | | N: | 113 | | | | Lg.: | | | (16) F: 1,8; (22) F: 2,46-1,67; M: 2,13-1,47; (35) F: 2,34-2,28; (47) F: 2,5-2; (73) F: 1,98-1,93; (101) F: 2,01-1,83; ?(142) F: 2-1; M: 2-1; (180) F: 2-1,8; (199) F: 1,9-1,67; M: 1,75-1,44; (237) F: 1,9-2,0; (290) F: 1,85; M: 1,85; (327) F: 2,22-1,95; M: 1,69-1,67; (432) F: 2,45-2,25; (909) F: 2,3; M: 1,72-1,8; (1023) F: 1,92; (1108) F: 1,92-2,4; M: 1,63-1,83; {F: 1,67-2,50; M: 1,44-2,13}
The mean female size is 2.051 mm (n = 26; SD = 0.2483), and the mean male size is 1.726 mm (n = 11; SD = 0.1888). The size ratio (male : female) is 0.86 (n = 6; SD = 0.0851). | | | | Rem.: | epi-mesopelagic. Considered as vetical migrator.
Sampling depth (sub-Antarct.) : 0-600 m. Overall Depth Range in Sargasso Sea: 0-1500 m (Deevey & Brooks, 1977, Station "S");
After Björnberg (1963, p.51) this species was prospering off the Brazilian coast in cooler waters (17.76 to 18.00°C of high salinity.
For Bradford-Grieve (1999 b, p.120) the identity of the males P. borealis and related species is not always clear, therefore is a possibility that P. gracilis, P. piseki and P. borelis have been cofused when they have not ben dissected.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is omnivorous.
After Cheng F. & al. (2013, p.125) the intraspecific genetic divergence revealed geographical differentiation. The biggest genetic divergence observed was between specimens sampled in westerly Antarctic areas and the Prydz Bay.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 4 corresponding to small filter feeding herbivorous and mixed feeding omnivorous (mostly broadcasters). | | | Last update : 28/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed January 05, 2026] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
