|
|
 |
|
Calanoida ( Ordre ) |
|
|
|
Clausocalanoidea ( Superfamille ) |
|
|
| |
| | | |
| Euchaetidae Giesbrecht, 1892 ( Clausocalanoidea ) | | Syn.: | Euchaetina Giesbrecht, 1892 (p.55);
Euchaetinae : Esterly, 1905 (p.156) | | Ref.: | Sars, 1902 (1903) (p.36); 1925 (p.97); Gurney, 1931 a (p.84); Rose, 1933 a (p.112); Sewell, 1929 (p.12, 132); 1947 (p.110); Brodsky, 1950 (1967) (p.81, 196); Mazza, 1967 (p.144); Andronov, 1974 a (p.1005); Park, 1978 (p.199, Rem. p.201-202 concerning Euchaeta and Paraeuchaeta); Razouls, 1982 (p.234); Bowman & Abele, 1982 (p.9); Bradford & al., 1983 (p.13); Brodsky & al., 1983 (p.144, 147); Zheng Zhong & al., 1984 (1989) (p.235); Sazhina, 1985 (p.113: N); Mauchline, 1987 (p.731, 734); Fontaine, 1988 (p.27, 30, Rem.); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.310); Mazzocchi & al., 1995 (p.176: Rem.); Park, 1995 (p.5, Redef., 95); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.797); Mauchline, 1999 (n°182, p.5: species Key); Bradford-Grieve & al., 1999 (p.880, 903; 923, Genera Key; Valdiviella includes in this family); Ohtsuka & Huys, 2001 (p.445, 461); Boxshall & Halsey, 2004 (p.118, Déf., Genera Key); Mulyadi, 2004 (p.67); Vives & Shmeleva, 2007 (p.644, Key G.); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, Biol. mol, phylogeny) ; Jeong & al., 2011 (p.117, species key: p.130); Laakmann & al., 2012 (p.535, Rem.: p.543, phylogeny); Laakmann & al., 2019 (p.330, fig. 2, 3, Table A, phylogenetic relationships); Hirai & al., 2020 (p.1, Fig.4: metabarcoding, Fig.8: OTUs distribution patterns)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | Le nombre de genres inclus dans cette famille diffère selon les auteurs, la discussion portant sur la validité de Paraeuchaeta.
La distinction des genres Euchaeta et Paraeuchaeta na pas été admis par tous les auteurs. Vervoort (1957) et Fontaine (1967) ne considèrent que le seul genre Euchaeta alors que Sewell (1947) et Tanaka (1958) reconnaissent lexistence des deux genres. Pour ces deux derniers auteurs le genre Paraeuchaeta se distingue du genre Euchaeta par les critères suivants :
Caudal rami : Les soies allongées sont toujours coudées et minces.
Mx1 : le nombre de soies au premier lobe externe varie dune espèce à lautre. Sewell (1947) divise le genre en 4 groupes suivant le nombre de soies.
Groupe 1 : 9 soies.
Groupe 2 : 7 soies.
Groupe 3 : 6 soies ; Groupe 4 : 5 soies.
Mx2 : Les soies sont garnies de courtes spinules (tandis que chez Euchaeta elles possèdent de longues spinules).
Mxp : Les longues soies spiniformes de lendopodite se terminent en crochet arrondi. Elles ne se prolongent pas comme chez Euchaeta par une petite pointe flagelliforme. Le bord interne se termine après une série de dents fines par 3 dents plus fortes.
P1 : La majorité des Paraeuchaeta possède une petite épine externe dans le creux de la concavité située au niveau de la fusion du premier er deuxième segment de lexopodite. Cette épine est absente chez Euchaeta.
P2 : La longueur de lépine externe du deuxième segment de lexopodite présente des variations. Tanaka (1958) en déduit 3 groupes :
Groupe 1 : lépine marginale externe dépasse lextrêmité de la première épine externe du troisième segment (distal) (à ce groupe appartient P. californica, P. soibogae).
Groupe 2 : lépine externe atteint presque lextrémité de la première épine externe du troisième segment (P. barbata,P. malayensis, P. bisinuata, P. weberi, P. confusa, P. calva, P. aequatorialis).
Groupe 3: l'épine dépasse la base mais n'atteint pas l'extrémité de l'épine marginale externe proximale (à ce groupe appartient P. sarsi, P. hanseni, P. comosa, P. propinqua, P. tonsa).
P5 male : Le troisième segment de lexopodite de la patte gauche est transformée en un doigt mobile toujours court et tronqué tandis quil est transformé en processus spiniforme chez Euchaeta.
Park (1995) redéfinit la famille et confirme l'existence des deux genres.
2 G.: Euchaeta, Paraeuchaeta.
After Madhupratap & al. (1996), la famille des Euchaetidae représente de 4 à 15,7 % des copepodes selon la saison dans la couche de mélange des eaux océaniques de la région ouest de l'Inde (Mer Arabe), en usant un filet type Multiple Plankton Closing Net à 200 µm de vide de maille (mesh aperture). | 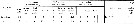 issued from : E.L. Markhaseva & F.D. Ferrari in Invert. Zool., 2005, 2 (2). [p.162, Table 4] Setation of oral parts in females Euchaetidae (Clausocalanoidea) and ancestral condition of setation. |
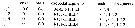 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.118]. Armature formula of swimming legs P1 to P4. Nota : - P5 absent in female. - Male P5 large, well developed, asymmetrical ; right leg with 2-segmented exopod and styliform ; 1-segmented endopod. Distal exopodal segment elongate, often styliform. Left leg with 2-segmented exopod and small distal process on elongate basis representing endopod. Distal exopodal segment with complex armature typically consisting of serrated lamella, ridged digitiform process, hirsute lobe and movable finger ornamented with row of setules near tip ; movable finger drawn out to form long spiniform process in some species. - Eggs retained in mass on ventral side of urosome . Nauplii lecihotrophic. |
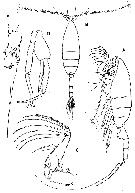 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.119, Fig. 23]. Euchaetidae. A, Paraeuchaeta norvegica (as Euchaeta norvegica) habitus female (lateral); B, habitus male (dorsal); C, female Mxp; D, Paraeuchaeta glacialis (as Euchaeta glacialis) male P5; R, Euchaeta pubera male P5 (distal part). [Sars, 1902: A-D; Park, 1975 a: E]. | | | | | (1) Euchaeta Philippi, 1843 | |
| | Syn.: | Euchirus Dana,1846; Euchäta : Giesbrecht, 1892 (p.55, 245); Euchaete : With, 1915 (p.157, clé spp.).
? Paraeuchaeta A. Scott, 1909 (p.69) | | Ref.: | Claus, 1863 (p.183); Giesbrecht & Schmeil, 1898 (p.37, clé spp.); Sars, 1902 (1903) (p.36); Esterly, 1905 (p.156, Key spp.); van Breemen, 1908 a (p.48, Key spp.); A. Scott, 1909 (p.64); Wolfenden, 1911 (p.293); Sars, 1925 (p.103); Sewell, 1929 (p.146); Wilson, 1932 a (p.62); Rose, 1933 a (p.112, clé spp.); Farran, 1936 a (p.92: Rem.); Mori, 1937 (1964) (p.42, Key spp.); Sewell, 1947 (p.110, Rev.); Davis, 1949 (p.33); Brodsky, 1950 (1967) (p.197, Key spp.); Vervoort, 1957 (p.74 & follow., Rev.); Tanaka, 1958 (p.327); Gaudy, 1963 b (p.13, Key spp.); Vervoort, 1963 b (p.157, Rem.); Owre & Foyo, 1967 (p.51, Rem., Key spp.); Tanaka & Omori, 1968 (p.219); Owre & Foyo, 1967 (p.51, clé spp.); Tanaka, 1973 (p.147); Bradford, 1974 (p.159); Park, 1975 c (p.1, clé spp.); 1978 (p.201, Rev., Key spp.); Séret, 1979 (p.88); Bradford, 1981 (p.398, Rem.); Razouls, 1982 (p.234); Gardner & Szabo, 1982 (p.259); Bradford & al., 1983 (p.13, Redef., p.18); Zheng Zhong & al., 1984 (1989) (p.236, Key spp.); Mauchline, 1988 (p.731: cuticular pores); Fontaine, 1988 (p.27, 30, Rem.: E. antarctica Group); Razouls, 1993 (p.310); Park, 1995 (p.13, 95, Redef., Key spp.); Chihara & Murano, 1997 (p.797); Mauchline, 1998 (p.92: F; p.93: M); 1999 (CIEM, n°182, p.5, Key spp.); Bradford-Grieve & al., 1999 (p.924, Key spp.); Boxshall & Halsey, 2004 (p.120); Mulyadi, 2004 (p.67); Vives & Shmeleva, 2007 (p.645, spp. Key) | | Rem.: | type: Cyclops marinus Prestandrea,1833. Le genre redéfini par Bradford & al. (1983, p.18) ne renferme que 4 spp.: E. indica , E. marina , E. marinella , E. rimana. Fontaine (1988) discute la validité de ce genre fondé sur l'espèce type P. norvegica Boeck,1872. De nombreuses espèces présentent des caractères intermédiaires entre les deux espèces types : E. marina et P. norvegica . Des groupes d'espèces ont été tentés comme E. marina et E. antarctica . Suivant les auteurs les espèces seront réparties, postérieurement aux descripteurs anciens, dans un genre ( With,1915; Vervoort,1957; Fontaine,1967; Bradford, 1972; 1974; Park,1975; Fontaine, 1988 ), ou deux genres ( A. Scott,1909; Sars, 1925; Rose, 1933 a; Sewell, 1947; Brodsky,1950; Tanaka,1958; Heptner,1971; Yamanaka, 1976; Bradford & al., 1983; Park, 1995 (p.13).
Park (1995) redéfinit le genre et établit 3 groupes: 'marina', 'concinna', 'acuta', plus 1 indépendante (E. spinosa), comportant un total de 16 spp. analysées (*).
Au total 16 spp. ont été recensées mais certaines nécessitent une description complémentaire: | | Remarques sur les dimensions et le sex-ratio: | | Le rapport des tailles male/femelle est de 0,944 (n = 13; Sd = 0.045), si l'on ne prend en compte que les rapports de chacune des espèces considérées présentant les mesures maximales et minimales de la taille des males et femelles. | 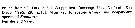 1 a: 1 a: Basis of female Mxp with long spinules in addition to short spinules along proximal half of medial margin (Fig.2-b). Exopod of male left P5 with poorly sclerotized lobes; serrated lamella of male left P5 exopod reaching or extending beyond tuft of stiff hairs (Fig.3-l)...... 2.
1 b: Basis of female Mxp with short spinules only along proximal half of medial margin (Fig.12-i). Exopod of male left P5 without poorly sclerotized lones (Fig.12-p-. Serrated lamella of male left P5 far short of reaching tuft of stiff hairs....... 3.2 a: Two endopodal setae of female Mx2 armed with both long and short spinules (Fig.2-a). Third exopodal segment of male left P5 with a poorly sclerotized lobe next to tuft of stiff hairs (Fig.3-m)....... marina species Group. 2 b: One endopodal seta of female Mx2 aemed with both long and short spinules (Fig.7-h). Third exopodal segment of male left P5 without a poorly sclerotized lobe next to tuft of stiff hairs (Fig.8-g)....... concinna species Group. 3 a: First endopodal segment of female Mx1 with 1 seta. Both exopods of male P5 similar in length (Fig.12-n)....... acuta species Group. 3 b: First endopodal segment of female Mx1 with 4 setae (Fig.18-g). Left exopod male P5 much shorter than right exopod (Fig.18-l)....... E. spinosa. | | | | | (2) Paraeuchaeta A. Scott, 1909 | |
| | Syn.: | Pareuchaeta : Sars, 1925 (p.104, 111); Rose, 1933 a (p.117, Key spp.); Jespersen, 1934 (p.70); Farran, 1936 a (p.92: Rem.); Sewell, 1947 (p.110, 120); Brodsky, 1950 (1967) (p.203, Key spp.); Vervoort, 1957 (p.75 & suiv., Rem.); Tanaka, 1958 (p.327, 333); Heptner, 1968 (p.680); 1971 (p.76, Rem.: 4 Groups); Zvereva, 1976 (p.70); Heptner, 1987 (p.1177, 1188, Rem.); Mauchline, 1988 (p.731: cuticular pores); Park, 1994 (p.317: Biogéo); Mauchline, 1998 (p.92: F; p.93: M) | | Ref.: | A. Scott, 1909 (p.69); Brady, 1918 (p.20); Sewell, 1929 (p.154); Wilson, 1932 a (p.65); Sewell, 1947 (p.110, 120: 4 Groupes); Bradford, 1981 (p.398, Rem.); Razouls, 1982 (p.264); Bradford & al., 1983 (p.20, Redef.); Razouls, 1993 (p.310); Park, 1995 (p.29, Redef., Key spp.); Chihara & Murano, 1997 (p.798, Key spp.); Bradford-Grieve & al., 1999 (p.924, 925: Key spp.); Boxshall & Halsey, 2004 (p.120); Vives & Shmeleva, 2007 (p.656, spp. Key) | | Rem.: | Park (1995) définit 6 groupes d'espèces: 'malayensis' (p.30, 31, 97), 'pavloskii' (p.30, 66, 98), 'norvegica' (p.30, 71, 98), 'glacialis' (p.31, 81, 98), 'hebes' (p.31, 83, 99), 'antarctica' (p.30, 87, 99), plus 3 espèces indépendantes (p.30). A partir des critères définis par Park (1995) 66 spp. appartiennent à ce genre (*), 15 spp. semblent devoir y être référées (**), les espèces restantes nécessitent une description complémentaire.
Type species: Paraeuchaeta norvegica (Boeck, 1872).
Au total 88 spp. sont recensées (plus 3 indéterminées): | | Remarques sur les dimensions et le sex-ratio: | | Le rapport des tailles male/femelle is 0,8777 (n = 44; SD = 0,0946), si l'on ne considère que les espèces présentant les deux sexes. |  Issued from : J.H. Wickstead in Proc. Zool. Soc. London, 1962, 139 (4). [p.563, Fig.2]. Drawings from a specimen of a female Paraeuchaeta sp. to show the trap formed by the extended first and second maxillipeds [= Mx2 or maxilla, and Mxp or maxilliped]. A: Frontal aspect; B: Lateral aspect. Example of maily carnivorous forms. |
 malayensis malayensis species Group. 1 - Appendicular setae geniculated (See Fig.19-g in P. malayensis). 2 - Genital flange linguiform, lying along length of genital field and produced distally into a distinct posterior lobe (see Fig.19-d in P. malayensis). 3 - First inner lobe of Mx1 with 1+9+3 setae (see Fig.20-a in P. malayensis). Supralabrum pointing obliquely forward (See Fig.19-b in P. malayensis). |
pavlovskii species Group. 1 - Appendicular setae geniculated (See Fig.19-g in P. malayensis). 2 - Genital flange linguiform, lying along length of genital field and produced distally into a distinct posterior lobe (see Fig.19-d in P. malayensis). 3 - First inner lobe of Mx1 with 1+9+2 setae (see Fig.63-f in P. pavlovskii). Supralabrum pointing ventrad (Fig.63-a in P. pavlovskii). |
norvegica species Group. 1 - Appendicular setae geniculated (See Fig.69-b in P. norvegica). 2 - Genital flange in form of a triangle or conical process and neither elongated along genital field nor produced distally into a lobe (see Fig.69-c in P. norvegica). 3 - In Mx1, 1st inner lobe with 1+9+3 setae, 2nd inner lobe with 1 seta, basis with 4 or 5 setae (See Fig.69-f in P. norvegica). |
P. biloba. 1 - Appendicular setae geniculated (See Fig.77-b in P. norvegica). 2 - Genital flange in form of a triangle or conical process and neither elongated along genital field nor produced distally into a lobe (see Fig.77-c, d in P. biloba). 3 - In Mx1, 1st inner lobe with 1+9+1 setae, 2nd inner lobe without seta, basis with 3 setae (See Fig.77-f in P. biloba). |
P. grandiremis. 1 - Appendicular setae smoothly curved, not geniculated (See Fig.88-a, c in P. grandiremis). 2 - In Mx1, 1st inner lobe with 1+7+1 setae, 2nd inner lobe without setae, and 1st endopodal segment with 2 setae (See Fig.88-h in P. grandiremis). |
P. antarctica species Group. 1 - Appendicular setae smoothly curved, not geniculated (See Fig.84-e in P. antarctica). 2 - In Mx1, 1st inner lobe with 1+9+3 setae, 2nd inner lobe with 2 setae, and 1st endopodal segment with 3 setae or more setae (See Fig.84-h in P. antarctica). 3 - In Mx1, 2nd inner lobe with 2 setae. |
P. bisinuata. 1 - Appendicular setae smoothly curved, not geniculated (See Fig.86-f in P. bisinuata). 2 - In Mx1, 1st inner lobe with 1+9+2 setae, 2nd inner lobe with 1 seta, and 1st endopodal segment with 3 or more setae (See Fig.86-i in P. bisinuata). 3 - In Mx1, 2nd inner lobe with 1 seta. 4 - In Mx1, basis with 3 setae. |
P. hebes species Group. 1 - Appendicular setae smoothly curved, not geniculated (See Fig.80-f in P. elongata). 2 - In Mx1, 1st inner lobe with 1+9+3 setae, 2nd inner lobe with 1 seta, and 1st endopodal segment with 3 or more setae (See Fig.80-g in P. elongata). 3 - In Mx1, 2nd inner lobe with 1 seta. 4 - In Mx1, basis with 4 or 5 setae. 5 - In Mx2, one of endopodal setae armed with long spinules (Fig.80-h in P. elongata). |
P. glacialis species Group. 1 - Appendicular setae smoothly curved, not geniculated (See Fig.79-d in P. tumidula). 2 - In Mx1, 1st inner lobe with 1+9+3 setae, 2nd inner lobe with 1 seta, and 1st endopodal segment with 3 or more setae (See Fig.79-a in P. malayensis). 3 - In Mx1, 2nd inner lobe with 1 seta. 4 - In Mx1, basis with 4 or 5 setae. 5 - In Mx2, none of endopodal setae armed with long spinules (See Fig.20-b in P. malayensis). |
 Issued from : E.T. Park & F.D. Ferrari in A selection from Smithsonian at the Poles Contributions to International Polar year. I. Krupnik, M.A. Lang and S.E. Miller, eds., Publs. by Smithsonian Institution Scholarly Press, Washington DC., 2009. [p.163, Table 8] Endemic species of Paraeuchaeta found in four different areas of interest. A''+'' indicates presence. |
 Issued from : E.T. Park & F.D. Ferrari in A selection from Smithsonian at the Poles Contributions to International Polar year. I. Krupnik, M.A. Lang and S.E. Miller, eds., Publs. by Smithsonian Institution Scholarly Press, Washington DC., 2009. [p.165, Figure 1] Distribution of selected pelagic calanoids Paraeuchaeta of the Southern Oczean and their closest relatives. |
 Issued from : J.M. Bradford in N. Z. J. Mar. Freshw. Res., 1981, 15. [p.393, Fig.2]. Terminal part of exopod P2 of female Paraeuchaeta erebi illustrating code used to describe relative lengths of outer edge spines in both sexes. | | | | | | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 05 mars 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|



;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
