|
|
 |
Fiche d'espèce de Copépode |
|
|
( Ordre ) |
|
|
|
( Famille ) |
|
|
|
( Genre ) |
|
|
| |
| 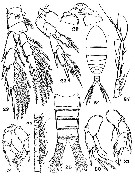 issued from : A.G. Humes & W.L. Smith in Carib. J. Sci., 1974, 14 (3-4). [p.139, Figs.22-31]. Female: 22, P4 (anterior; scale B); 23, P5 and intercoxal plate (anterior; scale B). Male: 24, habitus (dorsal; scale A); 25, urosome (dorsal; scale B);26, A1 (segments 8-18; postero-inner; scale C); 27, A1 (segments 19-24; postero-inner; scale C); 28, P5 (posterior; scale C); 29, intercoxal plate and coxae of P5 (posterior; scale D); 30, right P5 (anterior; scale B); 31, left P5 (anterior; anterior; scale B).
|
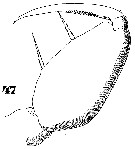 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Fig.47]. As Oncäa mediterranea var. flava. Female: 47, Mxp.
|
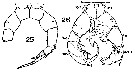 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.265]. As Acartia (Acartiella) keralensis. Female: Proportion of the segments on A1.
| | | | | NZ: | 0 | | |
|
Carte de distribution de par zones géographiques
|
| | | Saisie des informations en cours pour cette espèce | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2025. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 29 août 2025] © copyright 2005-2025 Sorbonne Université, CNRS
|
|
|
|



;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
