|
|
 |
|
Calanoida ( Order ) |
|
|
|
Clausocalanoidea ( Superfamily ) |
|
|
| |
| | | |
| Euchaetidae Giesbrecht, 1892 ( Clausocalanoidea ) | | Syn.: | Euchaetina Giesbrecht, 1892 (p.55);
Euchaetinae : Esterly, 1905 (p.156) | | Ref.: | Sars, 1902 (1903) (p.36); 1925 (p.97); Gurney, 1931 a (p.84); Rose, 1933 a (p.112); Sewell, 1929 (p.12, 132); 1947 (p.110); Brodsky, 1950 (1967) (p.81, 196); Mazza, 1967 (p.144); Andronov, 1974 a (p.1005); Park, 1978 (p.199, Rem. p.201-202 concerning Euchaeta and Paraeuchaeta); Razouls, 1982 (p.234); Bowman & Abele, 1982 (p.9); Bradford & al., 1983 (p.13); Brodsky & al., 1983 (p.144, 147); Zheng Zhong & al., 1984 (1989) (p.235); Sazhina, 1985 (p.113: N); Mauchline, 1987 (p.731, 734); Fontaine, 1988 (p.27, 30, Rem.); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.310); Mazzocchi & al., 1995 (p.176: Rem.); Park, 1995 (p.5, Redef., 95); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.797); Mauchline, 1999 (n°182, p.5: species Key); Bradford-Grieve & al., 1999 (p.880, 903; 923, Genera Key; Valdiviella includes in this family); Ohtsuka & Huys, 2001 (p.445, 461); Boxshall & Halsey, 2004 (p.118, Déf., Genera Key); Mulyadi, 2004 (p.67); Vives & Shmeleva, 2007 (p.644, Key G.); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, Biol. mol, phylogeny) ; Jeong & al., 2011 (p.117, species key: p.130); Laakmann & al., 2012 (p.535, Rem.: p.543, phylogeny); Laakmann & al., 2019 (p.330, fig. 2, 3, Table A, phylogenetic relationships); Hirai & al., 2020 (p.1, Fig.4: metabarcoding, Fig.8: OTUs distribution patterns)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | The number of genera included in this family differs according to the authors, the discussion dealing with the validity of Paraeuchaeta.
Park (1995) redefines the family and confirms the existence of the two genera.
2 G.: Euchaeta Philippi, 1843, Paraeuchaeta A. Scott, 1909.
Definition from Boxshall & Halsey (2004, p.118) :
- Cephalosome and pedigerous somite 1 partially or completely fused, pedigers 4 and 5 fused.
- Forehead pointed (rostrum with a single point).
- Nauplius eye present.
- Urosome female 4-segmented.
- Genital segment female projecting ventrally ; genital apparatus comprising common genital aperture located medially on ventral surface of genital double-somite ; copulatory ore contained within median genital aperture.
- Urosome male 5-segmented.
- Single genital male aperture located ventrolaterally at posterior rim of genital somite on left side.
- Caudal rami short, , seta VI, internal seta generally very long (accessory caudal seta), with up to 6 setae.
- A1 23 or 24-segmented in female ; segmental homologies (based on Paraeuchaeta norvegica (Boeck) : segment 1 (I) free, segment 2 (II-IV) triple, segments 3 (V) to 7 (IV) separate, segments 8 and 9 (X-XI) fused, segments 10 (XII) to 24 (XXVI) separate, apical (25th) segment double (VVVII-XXVIII). Aesthetascs present on segments 5, 8, 11, 13, 18, 24. Very long setae present on segments 3 (V), 7 (IX), 9 (XI) and 14 (XVI), forming 3-dimensional array.
- Male A1 non geniculate ; segmented fusions as in female except segments 8/9 and 10 ofthe fused to form triple segment (X-XII). Aesthetascs present on all segments except XI, XXII and XXV ; 2 aesthetascs present on segments II and III on E. marina (Prestandrea).
- Female A2 biramous, with separate or partly fused coxa and basis ; coxa with 1 seta, basis with 1 or 2 setae ; endopod 2-segmented ; compound distal segment bilobed,setation formula 2, 6, 7 ; exopod 7-segme,ted, segmental fusions indeterminate except for IX-X, setation formula 0, 1, 1, 1, 1, 1, 4. Endopod about 2/3 as long as exopod.
- Male A2 with reduced setation on coxa, basis and 1st endopodal segment.
- Female Md biramous ; coxa with well developed gnathobase and distal palp consisting of basis, armed with up to 2 setae , proximal seta typically claw-like ; 2-segmented endopod and indistinctly 5-segmented exopod ; endopodal segments 1 and 2 with 0/1 and up to 10 setae ; exopodal setation formula 1, 1, 1, 1, 2.
- Male Md gnathobase atrophied ; palp with reduced setation on basis.
- Female Mx1 with well developed praecoxal arthrite bearing (inner lobe 1) about 13 elements ; coxa with endite bearing 2 setae and with 5 to 9 setae on epipodite ; basis lacking outer seta, with proximal endite defined bearing 1 seta ; distal group of 3 setae representing distal endite ; endopod 2-segmented, proximal segment double, setation formula 5, 3 ; exopod 1-segmented, armed with up to 11 setae.
- Male Mx1 with atrophied praecoxal arthrite, coxal and basal endites and endopod ; exopod epipodite as in female.
Female Mx2 indistinctly segmented ; setation formula of praecoxal and coxal endites 3, 3, 3, 3 ; basis and endopod fused or partly separate ; basal endite with 3 elements, endopod retaining up to 6 elements.
- Male Mx2 very reduced, forming minute setose lobe.
- Female Mxp 7-segmented ; rotation at syncoxa-basis joint directing distal part of limb anteriorly ; syncoxa with endite setation formula 1, 2, 3, 3 ; basis with 3 setae, plus ½ setae on incorporated 1st endopodal segment ; free endopod 5-segmented, segmental setation formula 4, 2, 2, 2+1, 4 in E. rimana Bradford, or apparently reduced.
- Male Mxp smaller than in female ; syncoxal setation reduced.
- Swimming legs P1 to P4 biramous ; typically with 3-segmented rami, except P1 with 1-segmented endopod and P2 with 1 or 2-segmented endopod.
- 1st and 2 nd exopodal segments of P1 often partly or completely fused, particularly in females ; inner seta on basis of P1 situated on anterior surface of basis and passing across face of endopod.
Biology notes after Boxshall & Halsey (2004, p.120): Adult males are scarce and sexual dimorphism is very marked with the non-feeding adult males having atrophied mouthparts (see Boxshall, Yen & Strickler, 1997). The three dimensional array of long setae on the antennules of the adult female and copepodid stages functions for mechanoreception and plays an role in prey location in these predatory copepods (see Yen, 1988; Yen & Nicoll, 1990; Yen, Sanderson, Strickler & Okubo, 1991). This setal array is lost at the moult to the adult stage in males. | 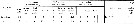 issued from : E.L. Markhaseva & F.D. Ferrari in Invert. Zool., 2005, 2 (2). [p.162, Table 4] Setation of oral parts in females Euchaetidae (Clausocalanoidea) and ancestral condition of setation. |
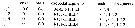 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.118]. Armature formula of swimming legs P1 to P4. Nota : - P5 absent in female. - Male P5 large, well developed, asymmetrical ; right leg with 2-segmented exopod and styliform ; 1-segmented endopod. Distal exopodal segment elongate, often styliform. Left leg with 2-segmented exopod and small distal process on elongate basis representing endopod. Distal exopodal segment with complex armature typically consisting of serrated lamella, ridged digitiform process, hirsute lobe and movable finger ornamented with row of setules near tip ; movable finger drawn out to form long spiniform process in some species. - Eggs retained in mass on ventral side of urosome . Nauplii lecihotrophic. |
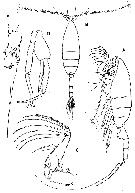 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.119, Fig. 23]. Euchaetidae. A, Paraeuchaeta norvegica (as Euchaeta norvegica) habitus female (lateral); B, habitus male (dorsal); C, female Mxp; D, Paraeuchaeta glacialis (as Euchaeta glacialis) male P5; R, Euchaeta pubera male P5 (distal part). [Sars, 1902: A-D; Park, 1975 a: E]. | | | | | (1) Euchaeta Philippi, 1843 | |
| | Syn.: | Euchirus Dana,1846; Euchäta : Giesbrecht, 1892 (p.55, 245); Euchaete : With, 1915 (p.157, clé spp.).
? Paraeuchaeta A. Scott, 1909 (p.69) | | Ref.: | Claus, 1863 (p.183); Giesbrecht & Schmeil, 1898 (p.37, clé spp.); Sars, 1902 (1903) (p.36); Esterly, 1905 (p.156, Key spp.); van Breemen, 1908 a (p.48, Key spp.); A. Scott, 1909 (p.64); Wolfenden, 1911 (p.293); Sars, 1925 (p.103); Sewell, 1929 (p.146); Wilson, 1932 a (p.62); Rose, 1933 a (p.112, clé spp.); Farran, 1936 a (p.92: Rem.); Mori, 1937 (1964) (p.42, Key spp.); Sewell, 1947 (p.110, Rev.); Davis, 1949 (p.33); Brodsky, 1950 (1967) (p.197, Key spp.); Vervoort, 1957 (p.74 & follow., Rev.); Tanaka, 1958 (p.327); Gaudy, 1963 b (p.13, Key spp.); Vervoort, 1963 b (p.157, Rem.); Owre & Foyo, 1967 (p.51, Rem., Key spp.); Tanaka & Omori, 1968 (p.219); Owre & Foyo, 1967 (p.51, clé spp.); Tanaka, 1973 (p.147); Bradford, 1974 (p.159); Park, 1975 c (p.1, clé spp.); 1978 (p.201, Rev., Key spp.); Séret, 1979 (p.88); Bradford, 1981 (p.398, Rem.); Razouls, 1982 (p.234); Gardner & Szabo, 1982 (p.259); Bradford & al., 1983 (p.13, Redef., p.18); Zheng Zhong & al., 1984 (1989) (p.236, Key spp.); Mauchline, 1988 (p.731: cuticular pores); Fontaine, 1988 (p.27, 30, Rem.: E. antarctica Group); Razouls, 1993 (p.310); Park, 1995 (p.13, 95, Redef., Key spp.); Chihara & Murano, 1997 (p.797); Mauchline, 1998 (p.92: F; p.93: M); 1999 (CIEM, n°182, p.5, Key spp.); Bradford-Grieve & al., 1999 (p.924, Key spp.); Boxshall & Halsey, 2004 (p.120); Mulyadi, 2004 (p.67); Vives & Shmeleva, 2007 (p.645, spp. Key) | | Rem.: | type: Cyclops marinus Prestandrea,1833. The genus redefined by Bradford & al. (1983, p.18) holds only 4 spp.: E. indica , E. marina , E. marinella , E. rimana. Fontaine (1988) discusses the validity of this genus founded on the type species P. norvegica Boeck,1872. Numerous species show characters, intermediate between the two type species: E. marina and P. norvegica. Species groups have been tempted as E. marina et E. antarctica . According to the authors the species would be distributed, subsequently to ancient descriptors, in one genus ( With,1915; Vervoort,1957; Fontaine,1967; Bradford, 1972; 1974; Park,1975; Fontaine, 1988 ), or two genera ( A. Scott,1909; Sars, 1925; Rose, 1933 a; Sewell, 1947; Brodsky,1950; Tanaka,1958; Heptner,1971; Yamanaka, 1976; Bradford & al., 1983; Park, 1995 (p.13).
Park (1995) redefines the genus and establishes 3 groups: 'marina', 'concinna', 'acuta', plus 1 independent (E. spinosa), comprising a total of 16 analysed (*) species.
In total, 16 species have been inventoried but certain need a complementary description.
Diagnosis after Bradford & al. (1983, p.18):
- Distal edge of Mx1 inner lobe 1 (arthrite) with at least 7 spines, posterior surface with up to 4 setae (total number of spines and setae on lobe = 11); inner lobe 2 or 3 absent, remaining lobe with 1 seta; basipod 2 with 3 setae, endopod with 4 setae, exopod with 11 setae, outer lobe with 5 setae.
- Females with 2 of the 6 apical setae of Mx2 covered with long spines in addition to short spinules .
- Male left P5 with thin-skinned lobe at base of long terminal spiniform process of exopod segment 3; digitiform process always spine-like.
Diagnosis after Park (1995, p.13) :
Appendicular caudal setae greatly developed, straight, and much thicker and longer than distal marginal setae of caudal ramus ;
Distal exopodal segment in both right and left P5 of male tapering into a long spine.
Type species: Euchaeta marina (Prestandrea, 1833).
Diagnosis after Boxshall & Halsey (2004, p.120) :
Mx2 female with 2 of the 6 apical setae ornamented with long setules in addition to short spinules.
Male left P5 with thin-walled lobe at base of long terminal spiniform process of 3rd exopodal segment
| | Remarks on dimensions and sex ratio: | | The mean female size is 3.588 mm (n = 29; SD = 1.2765), and the mean male size is 3.426 mm (n = 26; SD = 1.1899). The size ratio (male: female) is 0.9436 (n = 13; SD = 0.045). The sex ratio (Female: Male) is temporary 1,16, but likely 1. | | | | | (2) Paraeuchaeta A. Scott, 1909 | |
| | Syn.: | Pareuchaeta : Sars, 1925 (p.104, 111); Rose, 1933 a (p.117, Key spp.); Jespersen, 1934 (p.70); Farran, 1936 a (p.92: Rem.); Sewell, 1947 (p.110, 120); Brodsky, 1950 (1967) (p.203, Key spp.); Vervoort, 1957 (p.75 & suiv., Rem.); Tanaka, 1958 (p.327, 333); Heptner, 1968 (p.680); 1971 (p.76, Rem.: 4 Groups); Zvereva, 1976 (p.70); Heptner, 1987 (p.1177, 1188, Rem.); Mauchline, 1988 (p.731: cuticular pores); Park, 1994 (p.317: Biogéo); Mauchline, 1998 (p.92: F; p.93: M) | | Ref.: | A. Scott, 1909 (p.69); Brady, 1918 (p.20); Sewell, 1929 (p.154); Wilson, 1932 a (p.65); Sewell, 1947 (p.110, 120: 4 Groupes); Bradford, 1981 (p.398, Rem.); Razouls, 1982 (p.264); Bradford & al., 1983 (p.20, Redef.); Razouls, 1993 (p.310); Park, 1995 (p.29, Redef., Key spp.); Chihara & Murano, 1997 (p.798, Key spp.); Bradford-Grieve & al., 1999 (p.924, 925: Key spp.); Boxshall & Halsey, 2004 (p.120); Vives & Shmeleva, 2007 (p.656, spp. Key) | | Rem.: | Park (1995) redefines 6 species groups: 'malayensis' (p.30, 31, 97), 'pavloskii' (p.30, 66, 98), 'norvegica' (p.30, 71, 98), 'glacialis' (p.31, 81, 98), 'hebes' (p.31, 83, 99), 'antarctica' (p.30, 87, 99), plus 3 independent species (p.30). Following the criteria defined by Park (1995) 66 spp. belong to this genus (*), 15 spp. seem to have been referred to (**), the remaining species need a complementary description.
Type species: Paraeuchaeta norvegica (Boeck, 1872).
In total 88 spp. are inventoried (plus 3 unidentified).
Diagnosis after Bradford & al. (1983, p.20):
- Females with either : 1 of the 6 apical setae of Mx2 covered with long spines in addition to short spinules, or all 6 apical setae of Mx2 covered with spinules of uniform short length.
- Male left P5 without thin-skinned lobe on exopod segment 3, which may terminate in a long spiniform process, a short point, or be rounded.
Diagnosis after Park (1995, p.13) :
Appendicular caudal sete of female either geniculated or smoothly curved and distinctltly thinner but usually longer than principal marginal setae of caudal ramus.
3rd exopodal segment of male left P5 about as long as but much thinner than 2nd, with a very small, vestigial terminal spine.
Diagnosis after Boxshall & Halsey (2004, p.120) :
Mx2 female either with only 1 apical seta ornamented with long setules and short spinules, or with all 6 setae ornamented with uniformly short spinules.
Male left P5 without thin-walled lobe on 3rd exopodal segment, which may be rounded, pointed or form long spiniform process.
After Bode & al. (2018, p.75), the species of this genus are 'rheotactic' predators; they apply a 'drift and wait' feeding strategy, having extremely strong maxillipeds and feeding appendages, which even allow them to feed on large diapausing calanids (Auel & Hagen, 2002; Laakmann & al., 2009 a, 2009b), thereby taking advantage of a ''trophic shortcut'' from primary production in the surface layer to the deep sea via seasonally vertically migrating copepods (Hagen & al., 1995; Auel, 1999; Laakmann & al., 2009a, 2009b).
| | Remarks on dimensions and sex ratio: | | The mean female size is 7.240 mm (n = 160; SD = 1.9562), and the mean male size is 6.208 mm (n = 102; SD = 1.4919). The size ratio (Male: Female) is 0.878 (n = 44; SD = 0.0946). The sex-ratio (Female: Male) is temporary 1.58, but likely 1. | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 22, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|



;)
{kind=link}
{kind=link}
{kind=link}
