|
|
 |
|
Calanoida ( Order ) |
|
|
|
Arietelloidea ( Superfamily ) |
|
|
| |
| | | |
| Heterorhabdidae Sars, 1902 ( Arietelloidea ) | | Syn.: | Heterochätina Giesbrecht, 1892 (p.63);
Heterochaetidae Sars,1900 (p.79);
Heterorhabdinae : Esterly, 1905 (p.181) | | Ref.: | Sars, 1902 (1903) (p.73, 117); Gurney, 1931 a (p.84); Rose, 1933 a (p.198); Brodsky, 1950 (1967) (p.82, 341); Mazza, 1967 (p.190); Heptner, 1971 (p.146); 1972 a (p.54, Rev., Genera Key.); Grice, 1973 (p.943, Rem.); Andronov, 1974 a (p.1005); Razouls, 1982 (p.460); Bowman & Abele, 1982 (p.9); Brodsky & al., 1983 (p.143, 145); Zheng Zhong & al., 1984 (1989) (p.248); Mauchline, 1988 (p.708: cuticular pores); Huys & Boxshall, 1991 (p.463); Razouls, 1993 (p.307); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.816); Ohtsuka & al., 1997 (p.577, Phylogeny); Bradford-Grieve & al., 1999 (p.883, 902, 903, 942, 943: Key of Genera); Bradford-Grieve,1999 b (p.70, Def., Rem.); Park, 2000 (p.5, Rem., p.9: Genera & Sug-Genera Keys); Ohtsuka & Huys, 2001 (p.461); Boxshall & Halsey, 2004 (p.14; 49; 124: Def.; p.126: Geneta Key); Vives & Shmeleva, 2007 (p.292, part. Genera Key); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig. 2, 3, 4, molecular biology, phylogeny); Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | The family Heterorhabdidae was revised by Heptner (1992) who judges the used characters artificial, notably for the genera Hemirhabdus and Disseta. The species are regrouped on the basis of the structure of the mouth parts related to the type of food regime, evolving from filter-feeding mode (Disseta) to carnivorous sucking mode (Heterorhabdus). The author creates so the two new genera: Neorhabdus and Microdisseta. A comparitive examination of the Md, Mx1, Mx2 and Mxp guides the author to contemplate an evolution in the family. The genus Heterorhabdus is divided into two subgenera by Brodsky (1950): Paraheterorhabdus (type: H. robustus) and Euheterorhabdus (types: H. papilliger et H. norvegicus).
Grice (1973) adds to this family a new genus Alrhabdus following the discovery of a female (A. johrdae), which represents affinities with both the Heterorhabdidae and the Augaptilidae, but the author, in the absence of the knowledge of the male, does not create a specific family. He suggests that various species of hyperbenthic calanoids are in a comparable situation. Park (2000, p.1, 8, 141) excludes the genus from the Heterorhabdidae family as well as Microdisseta, maintained here for memory, and confirms the genus Paraheterorhabdus of Brodsky (1950).
Taxonomic notes after Boxshall & Halsey (2004, p.126) :
Park noted three synapomorphies for the heterorhabdidae within the Arietelloidea : the asymmetry of the caudal rami, with the left ramus usually longer than the right and fused to the anal somite, the marked elongation and lack of ornamentation on caudal seta IV on the left caudal ramus, and the presence of a large olumose inner lobe on the basis of the male right P5. Parks (2000) decision to exclude Microdisseta from the family Heterorhabdidae was based on the absence of these three synapomorphies. Park did not suggest an alternative placement for Microdisseta and his proposal is not adopted since several important characters were omitted from the phylogenetic analysis. The possibility that the failure to express these character states inn Microdisseta, with its very small body size, might be secondary.
The Heterorhabdidae was included in the superfamily Arietelloidea (as Augaptiloidea) by Andronov (1974) and Park (1986).
Biology notes after Boxshall & Halsey (2004, p.127) :
All genera are mainly distributed at mesopelagic and bathypelagic depths, with only a few species of Heterorhabdus and Heterostylites occurring in the epipelagic. For example, the smallest species of Heterorhabdus, H. papilliger is common an dis distributed in epipelagic to bathypelagic depths whereas larger congeners are less abundant and tend to be restricted to only mesopelagic and bathypelagic depths (Soh, 1998).
The carnivorous genera exhibit profound modifications in the labrum, mandibles and maxillae. The labrum with its large gland openings and the tubular, hypodermic needle-like ventralmost tooth on the mandible together form a mechanism that allows the labral gland secretion, possibly a venom, to be injected into the prey. Also in the carnivorous genera the maxilla is modified as a raptorial organ by the elongation of the praecoxal part and by the highly specialized setation elements on the basis and first endopodal segment.
Bioluminescence has been confirmed in species of Heterorhabdus, Hemirhabdus, Heterostylites, Neorhabdus, Disseta an dit is probable that all members of the Heterorhabdidae have a luminous capability (Herring,1988). The luminous glands can be numerous, a total of 36 in Heterorhabdus papilliger, for example, and they are located on the cephalothorax, urosome and limbs (after Giesbrecht, 1895a).
8 G: Microdisseta, Disseta, Hemirhabdus, Heterorhabdus, Heterostylites, Mesorhabdus, Neorhabdus, Paraheterorhabdus.
Definition after Bradford-Grieve (1999, p.70) Female :
- Body usually slender, prosome oval.
- Cephalosome and pediger 1 separated, pedigers 4 and 5 usually fused.
- Posterolateral corners of last thoracic segment rounded.
- Forehead rounded.
- Rostral filaments slender.
- Urosome elongate, 4-segmented.
- Genital segment with ventral process sometimes protruding greatly and sometimes occupying the whole length of segment.
- Caudal rami usually asymmetrical (largest on left) with varying degrees of fusion between them and anal segment, with 7 setae, one of the terminal setae often slightly or extraordinarly elongate on left side.
- A1 symmetrical, 25-segmented.
- A2 basipod 2 with 2 outer edge setae; exopod and endopod separated from basipod; endopod 2-segmented, segment 1 with 2 distally placed setae, segment 2 with 6-8 inner setae medially and 6-7 setae terminally; exopod 7-9-segmented, prpximal 2 segments sometimes without setae.
- Md blade usually asymmetrically developed on each side with 3-5 (or more) long, sharp teeth : in Heterorhabdus, Hemirhabdus and Neorhabdus one tooth is set far apart from remaining teeth; endopod 2-segmented, segment 1 with 2-4 setae, segment 2 with 6-9 setae terminally; exopod segments 1-4 with 1 setae each, segment 5 carrying 2 setae.
- Mx1 trends to have inner lobes 1-3 and endopod reduced; inner lone 1 with 5-15 spines and setae (most reduced setation in Hemirhabdus, least reduced in Disseta); inner lobe 2 small or absent with 0-2 setae; inner lobe 3 small or absent with 0-2 setae; basipod with 1-4 setae; endopod small, 1-segmented, with 4-111 setae; exopod large, with 5-11 setae (least reduced setation in Disseta, Mesorhabdus); outer lobe 1 with 3-8 setae.
- Mx2 usually modified as a grasping, piercing limb (least modified in Disseta and most modified in Hemirhabdus); lobes 1 and 2 with 3-5 and 0-3 reduced setae respectively; lobes 3 and 4 with 2-3 and 2-3 usually small setae respectively; lobe 5 with 1-3 setae 1 or 2 of which may be greatly enlarged and spine-like; endopod segment 1 usually with an enlarged spine-like seta; the remainder of the endopod with greatly reduced setae.
- Mxp basipod 1 with 3-7 setae the proximal one of which is greatly enlarged and spine-like in Heterorhabdus; basipod 2 slender with 3 small setae medially; endopod 6-segmented, segment 1 almost fully incorporated into basis with 2 setae, segments 2-5 with 3, 3, 2-3, 2,-3 setae respectively, segment 6 with 3-4 setae the outermost 1-2 of which are reduced.
- Swimming legs P1-P4 distinctly 3-segmented, exopods of legs P2-P4 flattened and leaf-like.
Diagnosis after T. Park (2000, p.5) :
1- Left caudal ramus normally fused to anal segment, longer than right ramus.
2- 4th marginal seta (counted from lateral) of left caudal ramus naked and greatly elongated.
3- Basis of male right P5 with a large, plumose inner lobe, which in some taxa is extremely large;
The Heterorhabdidae can be distinguished not only from the other families of the Arietelloidea but also from all other calanoid families by the above three characters. These 3 characters are highly pronounced in all species examined except for Paraheterorhabdus (Antirhabdus) compactus, in which the left caudal ramus is only slightly longer than the right and separated from the anal segment, and the 4th marginal seta of the left caudal ramus is longer than the other marginal setae but not greatly elongated. These character states of this species are believed to be reductive rather than primitive because the species shows an advanced stage of evolution in all other morphological features.
All the genera, subgenera, and species-groups of the Heterorhabdidae studied by Park (2000) are circumglobal in distribution, occurring widely in all three oceans and the pattern is very similar to that of the bathypelagic genus Paraeuchaeta and supports the hypothesis (Park, 1994), that endemic species with a limited geographic range are found exclusively in highly productive waters, where they are usually abundant. They are believed to be eutropgic. Species with a wide range of distribution are generally rare and are believed to be oligotrophic. The typical endemic species in each eutrophic area are as follows: Heterorhabdus norvegicus in the northern Atlantic; H. guineanensis in the eastern tropical Atlantic; Paraheterorhabdus longispinus, H. fistulosus, H. tanneri in the northern Pacific; Heterostyles echinatus, H. quadrilobus, H. prolixus, H. americanus, H. abyssalis in coastal waters of the eastern Pacific; H. farrani, H. pustulifer, H. spinosus, H. paraspinosus, H. austrinus in the Southern Ocean.
The body lenths are the bigger for the three genera Hemirhabdus, Neorhabdus and Disseta, respectively, followed by Mesorhabdus, Heterostylites, Paraheterorhabdus (Paraheterorhabdus), Heterorghabdus, respectively, lastly with sizes very small that are characteristic from the the preceding genera Paraheterorahbdus (Antirhabdus) and especially Microdisseta with body lengths less than 1 mm millimeter (± 0,740).
For Park (2000, p.6), the Heterorhabdidae seems to be most closely related to the Lucicuttidae according to the morphology of Mx2, in which 1 of the setae on the 5th and 6th lobes modified into a saberlike or falciform spine. The specializations in the Md, Mx2 and Mxp, serve as key characters not only among the genera of the Heterorhabdidae but also in distinguising some of the heterorhabdid genera from the other families of the Arietelloidea. | 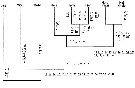 issued from : S. Ohtsuka, H.Y. Soh & S. Nishida in J. Crustacean Biol., 1997, 17 (4). [p.589, Fig.8]. Cladogramm showing phylogenetic relationships between genera of the family Heterorhabdidae generated with PAUP when all characters are set unordered. The numbers corresopnd to characters in Table 3 (following figure). Underline = multistate character; * = reversal; cross = convergence. Diss = Disseta; Micr = Microdisseta; Meso = Mesorhabdus; Hets = Heterostylites; Hemi = Hemirhabdus; Neo = Neorhabdus; Hete (Para) = Heterorhabdus (Paraheterorhabdus); Hete (Hete) = Heterorhabdus (Heterorhabdus). |
 issued from : S. Ohtsuka, H.Y. Soh & S. Nishida in J. Crustacean Biol., 1997, 17 (4). [p.587]. Characters used in the cladistic analysis (in the matrix) for genera on the family Heterorhabdidae. Codes 0-4 refer to transformation series of multistate characters. o = plesiomorphic state, 1 = advanced (apomorphic) state, 2-4 = further advanced states, 9 = missing data. |
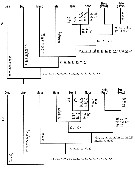 issued from : S. Ohtsuka, H.Y. Soh & S. Nishida in J. Crustacean Biol., 1997, 17 (4). [p.590, Fig.9]. Two cladograms showing phylogenic relationships between genera of the family Heterorhabdidae generated with PAUP when all characters are set-ordered. The numbers, symbols, and abbreviated generic names as in Fig. 8 and Table 3 (above). For the authors gut content analyses partly support Heptner's (1972) conclusion on the feeding habits of the family. The phylogenetic relationships among the genera of the family generated in the present study are almost identical with the noncladistic evolutionary tree of Hepner (1972), which was not based on detailed observations of the mandibular ventralmost teeth and the posterior surface of the labrum. The particle-feeder Disseta is the first to diverge from the main lineage, while the carnivores Hemirhabdus-Neorhabdus-Heterorhabdus are the terminal branch. Microdisseta is the second off-shoot in the evolution of the family. Mesorhabdus and Heterostylites are intermediate between these suspension feeders and the carnivores. The relative position of Heterostylites in the cladograms is equivocal. The specialization of the mandibular ventralmost tooth and the supposed paralytic-substance-releasing gland openings on the labrum are shared by Heterorhabdus and the Hemirhabdus-Neorhabdus clade. The evolutionnary transformation from particle feeders to pure carnivores has been accomplished by a combination of reductions or losses and specializations of several elements on the mouthpart appendages and the conversion of a lateral pair of labral secretory glands into a venom- or anaesthetic-realising system. Only in Heterorhabdus the relative elongation of the maxillary (Mx2) praecoxa is added to these transformations (length of Mx2 praecoxa in relation to body length female = 19.7). The elongation supposedly guarantees effective capture of prey animals (see in Ohtsuka, 1991). The remarkable elongation of the Mx2 praecoxa is found separately in Heterorhabdus and Heterostylites (ratio length of Mx2 praecoxa in relation to body length female = 14.4 in the latter) is not involved in the cladistic analysis. The lateral pair of large gland openings (type-1) on the posterior surface of labrum of Hemirhabdus-Neorhabdus-Heterorhabdus seem not be novel structures, but a conversion of two outer pores of two pairs of openings of ancestral type-3 glands into type-1 gland openings. In Disseta, Mesorhabdus, and Heterostylites, which lack type-1 glands, there are two pairs of openings of type-3 glands of almost equal size, while in Hemirhabdus, Neorhabdus, and Heterorhabdus, which possess type-1 glands, there is only one pair of openings of type-3 glands. This interpretation is supported by the fact that both type-1 and type-3 glands have one-paired gland cells and a common opening (see in Nishida & Ohtsuka, 1996); the conversion may be accompanied by an increase in the size of the gland system and the lateral movement of the position of the openings. In the Md, the reductions or losses of dorsal teeth and of daggerlike spinules have occurred throughout the evolution of the family. Heterostylites and Heterorhabdus may have independently developped the most advanced condition, the complete absence of these elements. The specialization of the ventralmost tooth and its isolation from the remaining teeth are evidently associated with the acquisition of the type-1 glands, and facilitate its protrusion from between the labrum and paragnath to inject a paralytic substance into prey (see in Nishida & Ohtsuka, 1996). The complete tubular structure of the ventralmost tooth of Heterorhabdus seems to be much effective in injecting a paralytic substance into prey than the semitube of Hemirhabdus and the groove of Neorhabdus. To avoid leaking the labral paralytic substance, the postrior side of the labrum presumably functions as an operculum to close the groove of Neorhabdus and the semitube of Hemirhabdus during injection of the substance into prey via the ventralmost mandibular tooth. The central teeth also appear to have become relatively elongate and slender in an evolutionary spep from Disseta to Mesorhabdus. In the Mx2, the transformations in the family can be explained by both developments and reductions of elements. Two setae on the basis and the first endopod segment are highly specialized into a grasping organ in the carnivores, with heavy sclerotization of the setae with hooked tips and an inner row of stout spinules. Reductions of setae on the second to fourth endopod segments are exhibited by all carnivores. Similar specializations and reductions of the Mx2 elements have convergently occurred also in the deep-sea predatory family Hyperbionychidae (see in Ohtsuka & al., 1993). |
 Issued from : E.T. Park & F.D. Ferrari in A selection from Smithsonian at the Poles Contributions to International Polar year. I. Krupnik, M.A. Lang and S.E. Miller, eds., Publs. by Smithsonian Institution Scholarly Press, Washington DC., 2009. [p.163, Table 8] Endemic species of several species of Heterorhabdidae found in four different areas of interest. A''+'' indicates presence. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.124]. Armature formula of swimming legs P1 to P5 Nota: Female P5 relatively unmodified, with 3-segmented rami; inner seta on 2nd exopodal segment strongly developed and directed medially. - Male P5 asymmetrical; biramous, typically with serrate coxa and basis, and 3-segmented rami; right leg typically with large medial process on basis. 1st exopodal segment with outer spine, 2nd with rounded or spinous inner process plus distal spine, 3rd typically forming curved claw armed with inner seta proximally and long subapical seta. Left leg with or without small medial process on basis; endopod formula 0-0; 0-1; 2,2,2, or with seta on middle segment absent. 1st and 2nd exopodal segmets each with outer spine, 3rd segment drawn out into apical claw, armed with inner and outer spines proximal to base of claw. - Eggs release into water. |
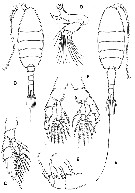 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.125, Fig. 25]. Heterorhabdidae. A, heterorhabdus robustus habitus female; B, Md: C, P5; D, habitus male; E, Hemirhabdus latus (as Macrorhabdus latus) female Mx2; F, Mesorhabdus gracilis male P5. [Sars, 1924]. |
 Issued from J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.71]. Spine and seta formula of swimming legs P1 to P5. - P5 female natatory, symmetrical, similar to other swimming legs but smaller; inner edge seta on exopod segment 2 ismodified into a long slender spine articulated with its segment; exopod segment 2 posterior distal border expanded into a toothed flange. - P5 male with a 3-segmented endopod and exopod on left and right; right basipod 2 usually with inner extensions of variable shape and usually fringed by hairs or spinules, sometimes with an inner extension on the left; right exopod segment 2 with an inner expansion of variable shape but usually in the form of a thick spine; left exopod segment 3 often but not always termunated by an elongate spine. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9]. Key of genera and subgenera of the family Heterorhabdidae. |
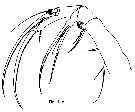
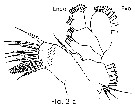 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 154, Fig.2 a]. Mx1 of Disseta palumbii female. |
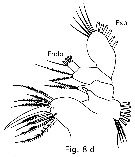 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 160, Fig.8 d]. Mx1 of Mesorhabdus angustus female. |
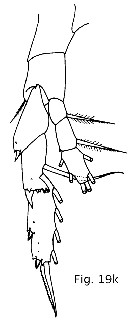 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 &171, Fig.19 k]. P5 of Heterostylites major female. |
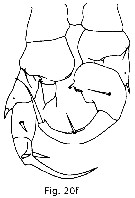 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 172, Fig. 20 f]. P5 (with endopods omitted, posterior view) of Heterostylites major male. Nota: female as 19 k. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 183, Fig.31 b]. Mx2 of Hemirhabdus grimaldii female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 187, Fig.35 b]. Left Mx2 (asp: anterior spine; psp = posterior spine; tsp = terminal spine) of Neorhabdus latus female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 196, Fig.44 d]. Mx2 of Paraheterorhabdus (Paraheterorhabdus) robustus female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 196, Fig. 44 a, b]. Md of Paraheterorhabdus (Paraheterorhabdus) robustus female. a: left mandible (posterior view); b: masticatory edge of right mandible (posterior view). |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 209, Fig.57 a, b]. Fig. 57 a, b: Md of Paraheterorhabdus (Antirhabdus) compactus female. a: left mandible (posterior view); b, masticatory edge of right mandible (posterior view). |
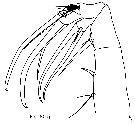 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.9 & 212, Fig.60 g]. Left Mx2 (posterior view) of Heterorhabdus spinifrons female. |
 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.268, Fig.116]. Morphological characters states in the genera Heterorhabdidae. |
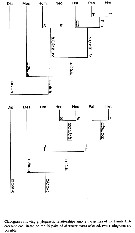 Issued from : T. Park in Bull. Scripps Inst. Oceanogr., Univ. California, San Diego, 2000, 31 [p.269, Fig.116 ]. Phylogenetic relationships among the genera of Heterorhabdidae. | | | | | Alrhabdus Grice, 1973 | | Ref.: | Grice, 1973 (p.943, 946); Razouls, 1982 (p.476); 1993 (p.307); Bradford-Grieve,1999 b (p.71, Déf.); Park, 2000 (p.1, 8, 141) | | Rem.: | Boxshall & Halsey (2004 (p.68, 126) transfer this genus to the Augaptilidae, however with some doubt as long as the male has not been encountered. | | | | (1) Disseta Giesbrecht, 1889 | |
| | Ref.: | Giesbrecht, 1892 (p.63, 369); Giesbrecht & Schmeil, 1898 (p.112); Esterly, 1906 a (p.71); A. Scott, 1909 (p.133); Wolfenden, 1911 (p.313); Sars, 1925 (p.221); Rose, 1929 (p.34, Rem.); Sewell, 1932 (p.308); Rose, 1933 a (p.198); Farran, 1948 d (n°15, p.3); Brodsky, 1950 (1967) (p.342, clé spp.); Tanaka, 1964 (p.32); Heptner, 1972 a (p.56, spp. Key); Razouls, 1982 (p.461); Mauchline, 1988 (p.706); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.816); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve,1999 b (p.71, Def.); Park, 2000 (p.11, Rem., spp. key); Boxshall & Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.293) | | Rem.: | type: Disseta palumbii Giesbrecht, 1899. Total: 3 spp.
The species of this genus appear to feed mainly on particles by suspension feeding.
Diagnosis after Bradford-Grieve (1999 b, p.71) :
- As for the family definition.
- Female genital segment usually with a greatly protruding ventral process.
- A1 exceeding the body in length.
- A2 with subequal rami.
- Masticatory edge of Md without a large space between some teeth; there are 4 wide ventral teeth, and remaining dorsal teeth small.
- Mx1 in the form of a fan with long plumose setae; distally exopod and endopod arranged together at one level; outer lobe 1 with 9 setae, exopod with 10 setae, endopod with 11-12 setae.
- Mx2 with distal part greatly reduced, setae long, plumose, and of almost equal size.
- Mxp basipod 1 shorter than basipod 2 by 1/5 of its length and without large naked curved spines; endopod segments 1 and 2 not more than 1.5 times as long as wide.
- Male P5 with right exopod segment 2 with its inner edge swollen and with an inner edge protuberance.
Diagnosis after T. Park (2000, p.11) :
- All principal marginal setae of caudal rami except for 4th on left ramus armed with small spines in addition to normal setules.
- 1st segment of A1 with 1+3+3+3 setae/aesthetes.
- In A2, all 8 exopodal segments each with a well-developed inner marginal seta.
- In Md, masticatory edge with a basal spine, a group of 6-8 small, contiguous teeth, and 3 or 4 large, spiniform teeth: basis and 1st endopodal segment each with 4 setae, and 2nd endopodal segment with 8 terminal and 2 anterior appendicular setae.
- Mx1 with 2nd and 3rd inner lobes bearing setae; exopod reaching distal end of endopod.
- In Mx2, 5th and 6th lobes each with 1 spine, which is shorter than the long, normal setae of the lobe.
- Coxa of Mxp with 4 lobes bearing setae;
- Basis of P1 with outer seta;
- 3rd endopodal segment of P2 with 8 setae.
In female P5, outer spinule of 2nd rxopodal segment flanked by 2 large spiniform processes of the segment;
- In male left , geniculated A1, segments 19-21 fused as 1st post-geniculate complex with 1 proximal and 2 distal setae, which appear to belong to 19th and 21st segment, respectively.
) In male P5, 2nd segment of left endopod without seta; 2nd segment of right exopod with spiniform or toothlike medial projection.
For Park (2000, p.13), the genus Disseta is the most primitive genus in the Heterorhabdidae and can be distinguished from the other genera of the family by such primitive features as the inner marginal setae on all exopodal segments of A2, a large number of teeth and setae on Md, a relatively short endopod of Mx1, relatively unspecialized setae and spines of Mx2, a large number of setae on the coxa of Mxp, and an outer seta on the basis of P1.
For Park (2000) Disseta is a bathypelagic genus. | | Remarks on dimensions and sex ratio: | | The mean female size is 8.295 mm (n = 6; SD = 1.7147) and the mean male size is 7.698 mm (n = 6; SD = 1.7321). The size ratio (Male : Female) is 0.929 (n = 3; SD = 0.0617). The sex ratio (Female : Male) is 1. | | | | | (2) Hemirhabdus Wolfenden, 1911 | |
| | Syn.: | Macrorhabdus Sars, 1920 c (p.11) | | Ref.: | Wolfenden, 1911 (p.308, Def.); Sars, 1925 (p.230); Sewell, 1932 (p.302, Rem.); Rose, 1933 a (p.205); Farran, 1948 d (n°15, p.3); Tanaka, 1964 a (p.26); Heptner, 1972 a (p.58, Rem.); Razouls, 1982 (p.477); Mauchline, 1988 (p.708); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.820); Mauchline, 1998 (p.70 : F; p.74: M); Bradford-Grieve,1999 b (p.76, Def.); Park, 2000 (p.51, 52: Key spp., Rem.: p.143); Boxshall & Halsey, 2004 (p.126); Vives & Shmeleva, 2007 (p.295) | | Rem.: | type: Heterochaeta grimaldii Richard, 1893 = Hemirhabdus grimaldii.
After Park (2000, p.143), the genus is bathypelagic.
Diagnosis after Bradford-Grieve (1999b, p.76) :
As for the family with additional characteristics :
- Left Md with 4 teeth, right with 5 teeth ; with ventral capped teeth separated from the remaining tooth by a gap.
- Mx1 outer lobe 1 with 4 setae, endopod with 6 setaze.
- Mx2 lobe 1 with 4 fine setae, lobe 2 atrophied, lobes 3 and 4 with 2 setae each, lobes 5 and 6 each with a strong claw-like spine clophed on its concave surface with short, widely spaced spinules.
- Mxp first basipod 1.5 times shorter tann basipod 2.
Diagnosis after Park (2000, p.51) :
- 1st segment of A1 with 1+3+3+3 setae/aesthetes.
- 1st and 2nd exopodal segments of A2 each with a small inner marginal seta.
- Basis and 1st endopodal segment of Md with 1 and 2 setae, respectively; 2nd endopodal segment without appendicular setae; masticatory edge with a basal spine followed by 3 teeth in left Md and 4 teeth in right Md, which are separated by an extremely wide gap from last, ventralmost tooth. Posteriorly ventralmost tooth with longitudinal ridges resembling a reinforcement rib.
- In Mx1, 3rd inner lobe missing; endopod 1-segmented, relatively well-developed; exopod elongate, accupying at distal end of appendage.
- In Mx2, 2nd lobe missing, 5th and 6th each with a large, coarely serrated saberlike spine in addition to 2 small setae; endopod with 2 long setae.
- Coxa of Mxp with a middle seta and 2 or 3 distal setae.
) 3rd endopodal segment of P2 with 7 setae.
- 3rd exopodal segment of P3 broad, with small terminal spine.
- In male P5, 2nd endopodal segment in broad legs with an inner marginal seta; 2nd exopodal segment of right leg with toothlike or fingerlike medial projection close to proximal end
The genus Hemirhabdus can be distinguished from Disseta, Mesorhabdus, Heterostyles by 1- Body massive; 2- masticatory edge of Md with ventralmost tooth separated from the other teeth by an extremely wide gap; 3- Mx2 with 2nd lobe missing, 3rd and 4th lobes with only small and unarmed setae, and endopod with only 2 setae; 4- Ledt A1 male with first 2 segments only partially fused and last 5 (21st to 25th) segments all separate.
Hemirhabdus shares with Heterostylites: 1- Mx1 with 3rd inner lobe missing; 2- masticatory edge of Md without a group of small, contiguous teeth next to basal spine; 3- coxa of Mxp with only 2 distal lobes bearing 1 seta on the 1st and 2or 3 setae on 2nd; 4- 3rd endopodal segment of P2 with 7 setae. | | Remarks on dimensions and sex ratio: | | The mean female size is 10.233 mm (n = 4; SD = 2.0230) and the mean male size is 10.585 mm (n = 4; SD = 1.6406). The size ratio (Male : Female) is about 1 . Le sex ratio (Female : Male) is 1. | | | | | (3) Heterorhabdus Giesbrecht, 1898 | |
| | Syn.: | Heterochaeta Claus, 1863 (p.180); Dahl, 1894 (p.77, Key spp.); Sars, 1900 (p.79); Heterochäta : Giesbrecht, 1892 (p.64, 372); Alloiorhabdus (part.) Wolfenden, 1911 (p.303); Vervoort, 1951 (p.140, Rem.)
Nota: Heterochaeta preoccuped name, therefore renamed Heterorhabdus by Giesbrecht & Schmeil (1898). | | Ref.: | Giesbrecht & Schmeil, 1898 (p.113, spp. Key); Sars, 1902 (1903) (p.117); Esterly, 1905 (p.182, clé spp.); van Breemen, 1908 a (p.117, spp. Key); A. Scott, 1909 (p.130); Wolfenden, 1911 (p.302); Sewell, 1932 (p.298); Wilson, 1932 a (p.131, spp. Key); Rose, 1933 a (p.199, spp. Key); Mori, 1937 (1964) (p.73); Sewell, 1947 (p.178, Rem.); Farran, 1948 e (n°16, p.3); Davis, 1949 (p.56); Brodsky, 1950 (1967) (p.344, spp. Key); Tanaka, 1964 a (p.1); Bradford, 1971 a (p.120, 135, spp. Key); Heptner, 1972 a (p.56, 59, spp. Key); Razouls,1982 (p.464); Gardner & Szabo, 1982 (p.363); Mauchline, 1988 (p.706); Zheng Zhong & al., 1984 (1989) (p.248); Chihara & Murano, 1997 (p.817); Mauchline, 1998 (p.70: F; p.74: M); Barthélémy, 1999 a (p.29); Bradford-Grieve & al., 1999 (p.943, 944: spp. Key); Bradford-Grieve,1999 b (p.76, Def.); Park, 2000 (p.89, Redef., Key of 4 Groups, Rem.: p.144); Boxshall & Halsey, 2004 (p.126); Vives & Shmeleva, 2007 (p.297, spp. key) | | Rem.: | type: Heterochaeta spinifrons Claus, 1863.
For Vervoort, 1951 (p.140) the genus Alloiorhabdus is based exclusively on the number of setae on the various segments of the swimming legs. It appears, however, that wolfendens observations are inaccurate as no differences in this respect exists between Heterorhabdus stricto sensu and the proposed genus Alloiorhabdus. The structure of the oral appendages in these genera too does not justify such a distinction.
Brodsky , 1950 (1967, p.345) establishes the 2 subgenera : Heterorhabdus (type: H. robustus) et Euheterorhabdus (types : H. papilliger & H. norvegicus).
Park establishes 4 groups ("spinifrons", "papilliger", "fistulosus", "abyssalis".
Type: Heterochaeta spinifrons Claus, 1863. Total: 31 spp. (+ 4 doubtful).
The species of this genus appear typical carnivores preying upon copepodids, polychaetes, and othe rmacrozooplanktoners (see litt. in Ohtsuka & al., 1997).
Diagnosis after Bradford-Grieve (1999 b, p.76) :
As for the family definition with the additional characteristics :
- Left Md with 3 teeth, right with 4 teeth.
- Mx1 is drawn out, the axis of exopod is of equal height to the remaining part which is aligned with exopod ; endopod reduced with 1-5 setae and sometimes atrophied.
- Mx2 lobe 1 with 2 setae, lobe 2 atrophied and replaced by 1 seta or spine, lobe 3 with 2 setae, lobe 4 with 3 long claw-like setae, the proximal parts of which have a partial comb of secondary spines or teeth, the distal part is smooth and sometimes flexed relative to proximal part.
- Mxp basipod 1 1.5 times shorter than basipod 2, proximal seta is often thickened, stronger, and noticeably longer than its segment (subgenus Heterorhabdus see Brodsky, 1950 as Euheterorhabdus) or the proximal seta is thin, plumose, and not longer than its segment (subgenus Paraheterorhabdus Brodsky, 1950).
- In some species P1 basipod 2 with a relatively large posteriorly-directed hook, visible in lateral view of whole copepod.
Diagnosis after Park (2000, p.89) :
- Dorsally, forehead with a midanterior tubercular process, which extends ventrad inyo tostrum.
- Sternite of the 1st pedigerous somite with a spiny papilla.
- Left caudal ramus fused with anal somite and distinctly longer than right.
- In 1st segment of A1, 1st lobe with a small seta, 2nd with 1 long and 1 very short setae and 1 aesthete, 3rd with 1 small and 2 medium-sized setae, 4th with 1 medium-sized and 1 long setae and 1 aesthete.
- Left geniculate A1 of male with segments 22 and 23 fused.
- In A2, first 2 exopodal segments without setae, last exopodal segment with a normally developed inner marginal seta, inner and outer lobes of 2nd endopodal segment with 8 (marginal) + 1 (posterior appendicular) and 6+1 setae, respectively.
- In Md, basis with 1 seta, 1st endopodal segment with 1 very small and 2 well-developed setae, 2nd with 8 terminal and 1 anterior appendicular setae; masticatory edge with basal spine followed by 2 teeth in left side and by 3 teeth in right side; ventralmost tooth separated from remaining teeth by a wide, flat gap. Ventralmost teeth of both Md symmetrical, large, falciform, each extending proximad, in the form of a reinforcement rib attached to the tooth, along ventral margin of mandibular blade to a distance about equal to length of tooth protruding beyond masticatory edge.
- Mx1 wit exopod greatly elongated and 2nd inner lobe (= coxal endite) bearing a relatively short seta.
- In Mx2, 1st segment greatly elongated; 1st lobe small, papillate, with 2 unequal setae; 2nd represented by a short, stout spine attached directly to segment; 3rd represented by a small seta and a medium-sized stout spine, both arising directly from segment; 4th elongate, with a small posterior subterminal spine and 2 long, saberlike terminal spines of similar length; 5th lobe elongate, with a relatively short saberlike spine and a long falcate spine; 6th with a very small anterior seta and a long falcate terminal spine serrated with short spinules along proximal section of medial margin; endopd small, with 7 relatively short setae;
- Coxa odf Mxp with middle marginal spine developed into a long, saberlike spine reaching to distal end of basis; anterior distal spine large and stout and 2 posterior distal spines greatly reduced in size.
3rd exopodal segment of P3 broad, with small terminal spine.
For Park (2000, p.90), the most important characters shared by all species of this genus are a spiny papilla of the sternite of the 1st pedigerous somite and a saberlike of falciform spine on the coxa of Mxp. Also important for identification of this genus are Mx1, which has a small endopod bearing 3 or fewer setae and an elongate exopod, and Mx2 in which the 5th lobe has 2 large, saberlike or falciform spines. The male P5 are characteristic in having elongate exopods and the terminal spine of the left exopod greatly elongated and completely fused with the segment.
After Bode & al. (2018, p.75) the species of this genus are abundant in the lower epipelagic and upper mesopelagic zone, these specimens may inject venom into their prey via a mandible strong tooth (Nishida & Ohtsuka, 1996; Ohtsuka & al., 1997; Wishner & al., 2008). | | Remarks on dimensions and sex ratio: | | The mean female size is 3.249 mm (n = 62; SD = 2.6382) and the mean male size is 2.792 mm (n = 62; SD = 0.6757). The size ratio (Male : Female) is 0.859 or if we consider the mean size for each species, we obtain 0.959 (n = 31; SD = 0.0512). The sex ratio est 1.
For the first group (''spinifrons''): the mean female size is 2.881 mm (n = 14; SD = 6860) and the mean male size is 2.719 mm (n = 14; SD = 0.6035). The size ratio (male : female) is 0.944. The sex ratio is 1.
For the second group (''papilliger''): the mean female size is 2.084 mm (n = 10; SD = 0.3832) and the mean male size is 2.047 mm (n = 10; SD = 0.3970). The size ratio (male : female) is 0.982. The sex ratio is 1.
For the third group (''fistulosus''): the mean female size is 3.278 mm (n = 4; SD = 0.6592) and the mean male size is 3.308 mm (n = 4; SD = 0.7100). The size ratio (male : female) is 1.009. The sex ratio is 1.
For the fourth group (''abyssalis''): the mean female size is 3.152 mm (n = 34; SD = 0.7383) and the mean male size is 2.980 mm (n = 34; SD = 0.6095).The size ratio (male : female) is 0.946. The sex ratio is 1.
| | | | | (4) Heterostylites Sars, 1920 | |
| | Ref.: | Sars, 1920 c (p.11); 1925 (p.237); Sewell, 1932 (p.300); Rose, 1933 a (p.207); Farran, 1948 d (n°15, p.3); Davis, 1949 (p.60); Brodsky, 1950 (1967) (p.357); Tanaka, 1964 a (p.23); Heptner, 1972 b (p.58); Razouls, 1982 (p.473); Gardner & Szabo, 1982 (p.377); Mauchline, 1988 (p.707); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.816); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve & al., 1999 (p.943, 945: spp. Key); Bradford-Grieve,1999 b (p.88, Déf.); Park, 2000 (p.37, 39: spp. Key, Rem.: p.143); Boxshall Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.312, spp. Key) | | Rem.: | Type: Heterochaeta longicornis Giesbrecht, 1889. Total: 6 spp.
The species of this genus appear to feed mainly on particles by suspension feeding, additionally feeding on zooplankters, such as copepods (see in Harding, 1974 and Ohtsuka & al., 1997). For Park (2000, p.143), This genus is meso-bathypelagic, occurring widely in the world's oceans. Of the 6 species found by Park (2000), H. major and H. longicornis have the widest ranges, occurring throughout the Pacific, Atlantic, and Indian Oceans, except for Arctic and Antarctic waters. The most common of the six species found were the the two Pacific species, H. longioperculis and H. echinatus, each representing 30 % of the specimens found for the genus, followed by H. longicornis and H. major accounting for 20 % and 16 %, respectively
Diagnosis after Bradford-Grieve (1999 b, p.88) :
- As for the family definition.
- Genital segment with a protruding ventral process with a large genital valve.
- A1 much longer than the body.
- Masticatory edge of Md with 3 teeth on the left and 4 teeth on the right; without a wide space between 1 tooth and the rest.
- Mx1 with 1 thick plumose spine on the proximal border of inner lobe 1 some distance from the terminal broup of finer spines; endopod with 5 setae.
- Mx2 lobes 5 and 6 each with 1 smooth, thickened hook-like seta; lobe 4 is the largest; the distal part of the limb is reduced, without large spines.
- Mxp basipod 1 is 1.5 times shorter than basipod 2.
- Female P5 with a complex crest of fused spines near the base of outer spine of exopod segment 2.
- Male P5 right exopod segment 2 inflated on inner border but without a definitr process.
Diagnosis after Park (2000, p.37) :
- None of marginal setae of caudal rami armed with conspicuous spines in addition to normal setules.
- 1st segment of A1 with 4 lobes bearing 1, 3, 2, 2 setae/aesthetes.
- In exopod of A2, first 2 exopodal segments without setae.
- In Md, masticatory edge with a basal spine and 3 or 4 long spiniform teeth, without a group of short contiguous teeth next to basal spine; basis with a long seta; 1st endopodal segment with 2 setae; 2nd endopodal segment with only 8 terminal setae, witout appendicular setae.
- mx1 with 3rd inner lobe missing; exopod greatly elongated, making up distal half of appendage.
- In Mx2, 5th and 6th lobes each with a large saberlike spine; 1 of 3 setae on 4th lobe reansformed into a long spine.
- Coxa of Mxp with 1 middle and 3 distal marginal setae ( 1 anterior and 2 posterior; first posterior distal seta greatly elongated).
- Basis of P1 without outer seta.
- 3rd endopodal segment of P2 with 7 setae.
- Anteriorly, 2nd exopodal segment of female P5 with distal margin extended distad into a serrated lappet.
- Left geniculated A1 of male with 2 setae belonging to 20th segment. Both right and left endopods of male pair of legs with inner marginal seta on 2nd segment. 2nd exopodal segment of right P5 much wider than long, with short medial projection normally divided distally into 2 processes (a spiny and a plumose process).
For Park (2000, p.38), the genus Heterostylites differs from Mesorhabdus and Disseta by: 1 - 1st segment of A1 with 1+3+2+2 setae/aesthetes, instead of 1+3+3+3 setae/aesthetes. 2 - first 2 exopodal segments of A2 witout setae. 3 - masticatory edge of Md without a group of short, contiguous teeth next to basal spine and 2nd endopodal segment with only 8 terminal setae; 4 - Mx1 with 3rd inner lobe missing; exopod greatly elongated, making up distal half of appendage. 5 - In Mx2, 1 or 3 setae on 4th lobe transformed into a long spine. 6 - coxa of Mxp with 4 setae, 3 on distalmost lobe and 1 seta on penultimate lobe; 1st pposterior seta of distalmost lobe much longer than the others. 7 - in basis of P1, outer seta missing as in Mesorhabdus. 8 - in P2, 3rd endopodal segment with 7 setae, instead of 8. 9 - anteriorly, distal margin of 2nd exopodal segment of female P5 extending distad into a serrated lappet; 10 - in male P5, 2nd endopodal segment in both legs with an inner marginal seta. 11 - in right P5 male, 2nd endopodal segment greatly enlarged, with a small but hugkly specialized medial projection.
The females of Heterostylites can be distinguished from those of the other calanoid copepods by a unique feature of P5: the serrated distal lappet of the 2nd exopodal segment. The male can be distinguished from those of the other heterorhabdid genera by the greatly enlarged 2nd exopodal segment of the right P5. | | Remarks on dimensions and sex ratio: | | The mean female size is 4.133 mm (n = 12; SD = 1.0460) and the mean male size is 3.948 mm (n = 12; SD = 0.8596). The size ratio (Male : Female) is 0.955. The sex ratio (Female : Male) is 1. | | | | | (5) Mesorhabdus Sars, 1905 | |
| | Ref.: | Sars, 1905 c (p.9); A. Scott, 1909 (p.132); van Breemen, 1908 a (p.126); Wolfenden, 1911 (p.312); Sars, 1925 (p.233); Sewell, 1932 (p.308); Rose, 1933 a (p.206); Farran, 1948 d (n°15, p.3); Tanaka, 1964 a (p.29); Heptner, 1972 a (p.57, Key spp.); Razouls, 1982 (p.474); Mauchline, 1988 (p.707); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.817); Mauchline, 1998 (p.70: F; p.74: M); Bradford-Grieve,1999 b (p.90, Rem.); Park, 2000 (p.23, Key spp., Rem.: p.143); Boxshall & Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.317, Key spp.) | | Rem.: | type: Mesorhabdus annectens (= Heterorhabdus brevicaudatus). Total: 5 spp.
After park (2000, p.143), Mesorhabdus is a meso-bathypelagic genus.
Diagnosis after Bradford-Grieve (1999 b, p.90) :
- As for the family definition.
- Left mandibular blade with 6 teeth, right blade with 7 teeth; teeth are distributed in 2 groups not separated by a space, there are 3 wide teeth on left and 4 wide teeth on right; remaining teeth are small and merge together.
- Mx1 outer lobe 1 and endopod with 5-6 and 3-4 setae respectively; exopod with 5-11 setae.
- Mx2 with 2 proximal endopod segments each with a strong claw-like spine bordered on all sides with fine difficult-to-observe hairs; proximal spine is longer, distal spine is stronger.
- Mxp basipod 1 more than 1.5 times (up to 2 times) shorter than basipod 3.
Diagnosis after T. Park (2000, p.23) :
- 3rd marginal seta of left caudal ramus and 3rd and 4th marginal setae of right caudal ramus armed with small spines in addition to normal setules.
- 1st segment of A1 with 1+3+3+3 setae/aesthetes.
- Masticatory edge with a basal spine, a group of 2 or 3 short, contiguous teeth, and 3 or 4 long spiniform teeth ; 2nd endopodal segment with 8 terminal setae plus 2 appendicular setae.
- Mx1 with 1 of setae on 1st inner lobe developed into a conspicuously large spine; 3rd inner lobe with 1 or 2 setae; exopod extending beyond distal end of endopod by 1/2 its length.
- In Mx2, 5th and 6th lobes each with a large, saber-like spine.
- Coxa of Mxp with 3 lobes bearing setae.
- Basis of P1 without outer seta.
- P2 with 8 setae on 3rd endopodal segment.
- Left A1 male with 0 or 1 seta belonging to 20th segment.
- 2nd endopodal segment of male left P5 without seta. 2nd exopodal segment of male right P5 with a large toothlike medial projection.
For Park (2000, p.24), male's geniculated A1 and P5 are basically the same as those of the Disseta male, but the male can be distinguished from that of Disseta by the medial projection of the 2nd exopod of the right P5, which is relatively long and extends at right angle to the longitudinal axis of the appendage. | | Remarks on dimensions and sex ratio: | | The mean female size is 5.287 mm (n = 10; SD = 1.4503) and the mean male size is 5.326 mm (n = 9; SD = 1.5488). The size ratio (Male : Female) is about 1 (0.948 to 1.038). The sex ratio (Female : Male) is 1. | | | | | (6) Microdisseta Heptner, 1972 | |
| | Ref.: | Heptner, 1972 a (p.60); Razouls, 1982 (p.463); 1993 (p.307); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve,1999 b (p.91, Def. Rem.); Park, 2000 (p.1, 7,141); Boxshall & Halsey, 2004 (p.127); Vives & Shmeleva, 2007 (p.320) | | Rem.: | Type: Disseta minuta Grice & Hulsemann, 1965. Total: 1 sp.
Boxshall & Halsey (2004, p.126) do not follow the opinion of Park on the uncertainty of the place of this genus within this family. Total: 1 species.
The feeding habits of species are still enigmatic, but probably mainly employs suspension feeding on the basis of their similarities in the mouthpart structures to those of Disseta and Heterostylites (see the fine structures of Md and labrums in Ohtsuka & al., 1997).
Diagnosis after Bradford-Grieve (1999 b, p.91) :
- As for the family definition.
- Md with 6 conical, ungrouped teeth of one type.
- The body of Mx1 is drawn out so that the distal part of the exopod only extends as far as the proximal part of the endopod; outer lobe 1 with 6 setae, exopod with 7 setae, endopod with 9 setae.
- Mx2 lobes 1-4 with one type of long plumose setae; lobe 5 with a strong, slightly shorter, crescent-shaped spine.
- Mxp basipod 1 noticeably longer than basipod 2; endopod segment 1 equal to segment 2, twice as long as wide and more than half the length of basipod 2.
After Boxshall & Halsey (2004, p.127) the genus Microdisseta is doistinguished from Disseta by the basal endite of Mx2 with claw-like element plus 1 seta in the former and without claw-like element, armed with 3 setae in the latter. | | Remarks on dimensions and sex ratio: | | The mean female size is 0.740 mm (0.68 to 0.80 mm) and the mean male size is 0.74 mm (0.73 to 0.75 mm). The size ratio (Male : Female) is 1. The sex ratio (Female : Male) is 1. | | | | | (7) Neorhabdus Heptner, 1972 | |
| | Ref.: | Heptner, 1972 a (p.58, Key spp.); Razouls, 1982 (p.476); 1993 (p.307); Bradford-Grieve,1999 b (p.92, Déf., Rem.); Park, 2000 (p.57, 59: Key spp., Rem.: p.143); Boxshall & Halsey, 2004 (p.126); Vives & Shmeleva, 2007 (p.320) | | Rem.: | type: Heterorhabdus latus Sars, 1905. Total: 5 spp.
After Park (2000, p.143), the genus is bathypelagic occurring mainly in the low latitudes. Of the 5 species found by the author, N. latus and N. brevicornis were commonly found throughout the low latitudes of the world's oceans and represented 61 % and 20 %, respectively of the specimens found for the genus. N. falciformis and N. capitaneus were found in all three major oceans but they were very rare, each representing only 8 % of the total found for the genus. N. subcapitaneus was represented by only 3 specimens found in the northwestern Atlantic and southeastern Pacific.
Diagnosis after Bradford-Grieve (1999 b, p.92) :
- As for the family definition.
- Left mandibular blade with 4 teeth, right blade with 5 teeth; ventral capped teeth separated from remaining tooth by a gap.
- Mx1 outer lobe 1 with 3 setae.
- Mx2 with lobes 1-4 developed, with setae, part of which is thickened basally, sometimes they are claw-like, abruptly tapering distally, densely covered in fine hairs; 1-2 of these setae on every lobe is curved in the direction of the distal part of limb; lobes 5 and 6 relatively short, each with a strong claw-like spine with a row of spinules on the concave border.
- Mxp basipods 1 and 2 are more or less equal.
Diagnosis after Park (2000, p.57) :
- Body massive.
- In 1st segment of A1, 1st lobe with 1 seta, 2nd with 2 setae + 1 aesthete, 3rd with 2 setae + 1 or more aesthetes, 4th with 2 setae and 2 or more aesthetes.
- 1st and 2nd exopodal segments of A2 without appendicular setae.
- In Md, basis without seta, 1st endopodal segment with 2 setae, 2nd endopodal segment with 7 terminal setae and without appendicular seta; masticatory edge with a basal spine followed by 3 teeth in left sideb and 4 teeth in right side, which are separated from last, ventralmost tooth by a wide gap.
- 2nd inner lobe of Mx1 with or without a seta, 3rd inner lobe without seta; endopod 1-segmented and small; exopod elongate, occupying at distal end of appendage.
- Mx2, all setae on 1st to 4th lobes modified into spines, either unarmed or armed with short spinules only; 5th and 6th lobes of equal length, each with a large saberlike, serrated spine in addition to a small spine; endopod vestigial , with 2 setae of equal length.
- Coxa of Mxp with 1 middle seta and 2 or 3 distal setae.
- 3rd endopodal segment of P2 with 7 setae.
- In male, P5, 2nd endopodal segment with 1 seta in both legs; 2nd exopodal segment of right leg of P5 with a digitiform medial projection.
The species of this genus had been referred to Hemirhabdus until Hepner (1972a) recognized their differences from the latter and placed them in a new genus, Neorhabdus. However, neorhabdus is most closely related to hemirhabdus and they share common features: 1- dorsally, forehead without midanterior tubercular process extending from rostral base; 2- caudal rami wide; left ramus only a little longer than right; 3- on masticatory edge, basal spine is followede by 3 teeth in left side and 4 in right side, which are separated from last, ventralmost tooth by an extremely wide gap; 4- ventralmost tooth with longitudinal ridges resembling a reinforcement rib; 5- in Mx1, 1st inner lobe elongate, with 9 or fewer setae, 3rd inner lobe without seta; 6- in Mx2, large spines of 5th and 6th lobes well developed, saberlike and conspicuously serrated with large teeth; endopod vestigial, with only 2 setae; 7- coxa of Mxp with 1 middle and 2 or 3 distal setae; 8- in male P5, each endopod with 1 inner seta on 2nd segment; 2nd segment of right exopod with digitiform medial projection.
The differences between the two genera can be distinguished from each other in Mx2, which in Neorhabdus has all setae on the first 4 lobes modified into spines and the 5th and 6th lobes of similar length. In Mx2 of Hemirhabdus the 2nd lobe is missing, none of the setae of the first 3 remaining lobes modified into spines, and the 5th lobe is much larger and longer than the 6th.
Additional differences between the genera are: 1- cephalosome in lateral view with a cervical groove in Hemirhabdus but without it in Neorhabdus; 2- A2 with a small inner marginal seta on 1st and 2nd exopodal segments and 1 posterior appendicular seta on inner and outer lobes of 2nd endopofal segment in Hemirhabdus; they are absent in Neorhabdus; 3- Md with 1 seta on basis and 8 terminal setae on 2nd endopodal segment in Hemirhabdus, but no seta on basis and 7 terminal setae on 2nd endopodal segment in neorhabdus; 4- Mx1 with 9 setae on 1st inner lobe, 1 seta on basis, and 5 long lateral and 3 small medial setae on exopod in Hemirhabdus, but 6 or 7 setae on 1st inner lobe, 2 or 3 setae on basis, and only 3 or 4 long lateral setae on exopod in Neorhabdus: 5- in P1, each outer exopodal spine is attached in a socket formed by spiniform processes of segment in Neorhabdus, but no such sockets in P1 of Hemirhabdus; 6- exopodal terminal spine of P3 is much smaller than that of P4 in Hemirhabdus, but the same spine of P3 is slightly larger than that of P4 in Hemirhabdus, but the same spine of P3 is slightly larger than of P4 in Neorhabdus; 7- in geniculate A1 male, 19-20th segments fused in Hemirhabdus, but 19th-21st segments fused in Neorhabdus. | | Remarks on dimensions and sex ratio: | | The mean female size is 9.140 mm (n = 10; SD = 2.3632) and the mean male size is 8.483 mm (n = 6; SD = 0.005). The size ratio (Male : Female) is 0,972 (n = 4; SD = 0.0662). The sex-ratio (Female : Male) is 1,25. (Probably equal in the genus) | | | | | (8) Paraheterorhabdus Brodsky, 1950 | |
| | Ref.: | Brodsky, 1950 (1967) (p.345); Park, 2000 (p.71, 2 S-G: Antirhabdus & Paraheterorhabdus, spp. Key, Rem.: p.143); Boxshall & Halsey, 2004 (p.126) | | Rem.: | After Park (2000, p.72) Paraheterorhabdus is a valid taxon chearly distinguishable of the genus Heterorhabdus and other genera of the family Heterorhabdidae in morphological details mainly of Mx2 and Mxp.
In Mx2, 1st segment elongate, about as long as the rest of the appendage, first 3 lobes with 2, 1 and 2 relatively small setae/spines, respectively ; 5th highly elongated, with a small seta and a long spine ; 6th lobe with 2 small setae and a long spine ; long spines of 5th and 6th lobes saberlike, slender, gradually tapering into sharp points, finely serrated along medial margins except for distal sections.
Coxa of Mxp with well-developed middle and distal spines ; middle spine elongate ; anterior distal spine thicker but shorter than 1st posterior distal spine ; 2nd posterior distal spine thinnest and shortest.
Paraheterorhabdus compactus (Sars, 1900) differs from the other species of the genus and a new subgenus is erected for it.
Total: 8 spp.
For Park (2000, p.143), the subgenus comprising 6 species seems to be mesopelagic, occurring widely in the world's oceans. P. robustus, P. illgi, P. medianus, P. vipera all have a similar range of distribution, occurring widely in the low and midlatitudes of all three major oceans. P. robustus was the most common, representing 26 % of the specimens found for the subgenus, followed in orfer by P. illgi, P. medianus, P. vipera accounting for 14 %, 10 % and 8 %, respectively. P. longispinus is endemic to the northern Pacific, where it seems to be quite common. P. farrani is distributed exclusively in the Southern Ocean, accounting for26 % of the total found for the subgenus, and one of the samples, taken in the Drake Passage, contaned as many as 61 specimens. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.516 mm (n = 16; SD = 0.9922) and the mean male size is 3.329 mm (n = 16; SD = 0.9397). The size ratio (Male : Female) is 0.952 (n = 8; SD = 0.0688) The sex ratio (Female : Male) is 1. | | | | | Antirhabdus Park, 2000 | | Ref.: | Park, 2000 (p.85, Def.) | | Rem.: | For Park (2000, p.144), the monotypic subgenus Antirhabdus is bathypelagic and circumglobal, occurring commonly throughout the low and midlatitudes of the world's oceans; Park found 266 specimens.
Subgenus diagnosis after Park (2000, p.85) :
- Left caudal ramus only slightly longer than right and fully separated from anal segment ; 4th marginal seta unarmed , longer than other marginal setae, and about as long as urosome. -
- A1 a little short of reaching posterior end of prosome.
- Masticatory edge of left Md with 1 small and 2 long teeth next basal spine and ventralmost tooth similar in size and shape to that of right Md.
- 1st inner lobe of Mx1 with 11 setae.
- In coxa of Mxp, middle marginal spine much shorter than segment and about as long as 1st posterior distal spine.
- Posterior hooklike of P1 basis elongate, tapering into a sharp point and only slightly curved.
Type: Heterochaeta compacta Sars,1900. 1 espèce.
Mean body length: Female : 2,85 mm; male: 1,96 mm. | | | | Paraheterorhabdus Brodsky, 1950 | | Ref.: | Brodsky, 1950 (1967) (p.345, 346); Park, 2000 (p.71, Redef.) | | Rem.: | Subgenus diagnosis after Park (2000, p.72) :
- Left caudal ramus distinctly longer than right and completely fused with anal segment ; 4th marginal seta slightly thicker than other marginal setae and about as long as body.
- A1 extending beyond posterior end of prosome.
- Masticatory edge of left Md with 2 subequal teeth next to basal spine and ventralmost tooth at least twice as long as that of right Md.
- 1st inner lobe of Mx1 with 12 setae.
- In coxa of Mxp, middle spine about as long as segment and longer than distal spines.
- Hooklike posterior process of P1 basis relatively large.
Type: Heterorhabdus robustus Farran, 1908. 7 species.
Mean body length female: 3,516 mm; male: 3,329 mm. | | | | | | | | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed October 18, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|



;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
