|
|
 |
| Oncaeidae Giesbrecht, 1893 ["1892"] ( Cyclopoida ) * | | Syn.: | Oncäidae part.Giesbrecht,1892 (p.81); Oncaeidae part.: Esterly, 1905 (p.215); van Breemen, 1908 a (p.184); Sars, 1918 (p.190); Rose, 1933 a (p.295); Mori, 1937 (1964) (p.119); Humes & Stock, 1973 (p.6); Heron, 1977 (p.38, Genera key); Boxshall, 1977 a (p.109, Def., Rem.); Heron & Damkaer, 1978 (p.3, Genera key); Björnberg & al., 1981 (p.665); Razouls, 1982 (p.665); Bowman & Abele, 1982 (p.12); Heron & al., 1984 (p.449); Zheng Zhong & al., 1984 (1989) (p.262, Genera key); Sazhina, 1985 (p.93); Boxshall & Huys, 1988/89 (p.164, 172); Huys & Boxshall, 1991 (p.465); Razouls,1993 (p.313); Heron & Bradford-Grieve, 1995 (p.11, 45, Rev., Genera key); Chihara & Murano, 1997 (p.978, Pl.220); Bradford-Grieve & al., 1999 (p.887, 969, Genera Key) | | Ref.: | Huys & Böttger-Schnack, 1996/97 (p.243, 246, Redef.); Heron & Frost, 2000 (p.1016, Genera Key); Boxshall & Halsey, 2004 (p.17; 613: Def.; p.615: Genera key. Rem. p.1: For the authors the order Poecilostomatoida is questionable); Di Capua & Boxshall, 2008 (p.1413: Key to species females in the Gulf of Naples); Vives & Shmeleva, 2010 (p.258, Rem., Genera key from Iberica Fauna); Böttger-Schnack & Machida, 1011 (p.111, DNA sequences, phylogeny); Böttger-Schnack & Schnack, 2013 (p.265, species groups: Def.); Böttger-Schnack & Schnack, 2015 (p.1, Table 1, 3, 5, morphological characters for identification); Hirai & al., 2020 (p.1, Fig.4: metabarcoding, Fig.8: OTUs distribution patterns, Fig.9: phylogenetic analysis) | | Rem.: |
Wickstead (1962, p.547, 548, food & feeding).
Boxshall & Halsley (2004, p. 615) acknowledge the genus Triconia, in spite of the arguments given by Heron & Frost (2000). The genus Pachos must be removed from this family (incertae sedis). 7 G.: Archioncaea, Conaea, Epicalymma, Monothula, Oncaea, Spinoncaea, Triconia.
After Böttger-Schnack (pers. comm., 2009), publication year of Giesbrecht´s monography on the pelagic copepods of the Gulf of Naples should be cited as Giesbrecht 1893 ["1892"] (for an explanation, see the note of Ruth Böttger-Schnack in the family site of Oncaeidae in the WoRMS database).
Key to genera after Boxshall & Halsey (2004, p.615):
1 - P5 bearing 3 setae; legs P1 to P4 with 2-segmented endopods ..........Archioncaea.
1' - P5 with at most 2 setae, or leg further reduced; legs P1 to P4 typically with 3-segmented endopods ............ 2.
2 - Caudal ramus with conspicuous dorsal expansion surrounding insertion of dorsal seta ....... 6.
2' - Caudal ramus without expansion on dorsal surface .......... 3.
3 - Female carrying single dorsal egg sac; 3rd endopodal segment of legs P2 to P4 lacking outer margin spine ........Monothula.
3' - Female carrying paired dorsal egg sacs; 3rd endopodal segment of legs P2 to P4 typically ** with outer margin spine ....... 4.
4 - Caudal ramus with seta III modified into strong spine ..........Spinoncaea.
4' - Caudal ramus with seta III setiform ......... 5.
5 - Adult females without mid-dorsal projection on 2nd pedigerous somite; endopod of leg P4 without distal conical process between apical and outer margin spines ............ Oncaea *.
5' - Adult females with or without mid-dorsal projection on posterior margin of 2nd pedigerous somite; endopod of leg P4 with distal conical process located between apical and outer margin spines .......... Triconia *.
6 - 3rd exopodal segment of legs P1 to P3 with II, III, III spines on outer margin ............ Epicalymma.
6' - 3rd exopodal segment of legs P1 to P3 with III, II, II spines on outer margin .......... Conaea.
* One exception : Oncaea insolita Heron & Frost, 300, also has a small conical process (addition to the version from Boxshall & Halsey, 2004, after R. Boettger-Schnack (30/11/2015, pers. comm.).
** However, the genus Oncaea includes also a number of species (16 species at present) lacking outer margin spines. (addition to the version from Boxshall & Halsey, 2004, after R. Boettger-Schnack, 30/11/2015, pers. comm.) | | * We thank Ruth Böttger-Schnack (Universität Bremen) for having forwarded to us a copy of the families Corycaeidae, Oncaeidae et Sapphirinidae from the Fauna of Japan by Chihara et Murano (1997), as well as the English translation concerning the Oncaeidae family performed by Hiroshi Itoh. |  Issued from : Boxshall & Halsey in The Ray Society, 2004, Part II. [p.613]. Armature formula of swimming legs P1 to P4. Nota: Inner seta on basis of P1 present or absent. Setation often modified by loss of elements from both rami. Endopod of P4 armature 0-1; 0-2; I, II, 2 or with terminal armature reduced to 0, II, I, or 0, II, 1, or 0, I, 1. P2 and P3 usually with conical process between distal spines on endopod; process present on P4 in Triconia. - P5 laterally located on somite; protopod incorporated into somite, usually represented by seta on surface of somite but absent in Archioncaea; exopod 1-segmented typically armed with 2 setae0, or reduced to single seta plus a spinule, on surface, or absent; exopod bearing 3 setae in Archioncaea. - P6 represented by genital opercula; armed with 1 seta in female, unarmed in male. - Egg sacs typically paired, multiseriate, single egg sac carried in Monothula. Remarks: See separation of genera and species groups showing leg armature type 1 (Table 4) in Böttger-Schnack & Schnack (2013, p.4). |
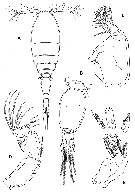 Issued from : Boxshall & Halsey in The Ray Society, 2004, Part II. [p.614, Fig.199). A, Triconia borealis (as Oncaea borealis) female; B, Oncaea media f minor male urosome; C, Oncaea venusta Mx2; D, A2; E, Md.
After Sars, 1918: A; Huys & Boxshall, 1991: B-E. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.5, Table 1].. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.10, Table 2]. In several cases some additional, secondary characters are required for a final identification of species. For example, the length to width ratio of the caudal ramus can be used to separate species within the genus Oncaea s. str. or the length of the outer basal seta on P5 can separate the five described species belonging to the dentipes-subgroup of Triconia. |
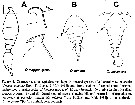 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.7, Fig.4]. |
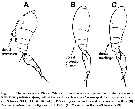 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.7, Table 5]. |
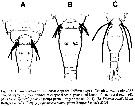 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.8, Fig.6]. Nota: Reed in (A) Oncaea serrulata and no serrata. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.8, Fig.7]. |
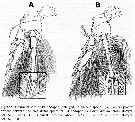 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2015. [p.9, Fig.8]. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.9, Table 2]. Main morphological characters of females used for separating genera and species groups of Oncaeidae |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.10, Table 2 (Continued)]. Main morphological characters of females used for separating genera and species groups of Oncaeidae |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.11, Table 3]. Morphological characters of oncaeid genera and of species groups within Oncaea s.l. Nota: Differences in labral structure have been found to be very important, separating e.g. the compacta-group, the olsoni-group, and the ovalis-group from closely related groups or genera. The distinct formation of a distal conical process on endopodite 3 of P4 is typically found in the genus Triconia (but see the discussion of O. insolita Heron & Frost). The number of elements on the endopodite 2 of A2 and/or their ornamentation separate the tregoubovi-group from the prendeli-group and are unique in the tenuimana-group. A very elogate P5 exopod segment is typically found only in the notopus-group. The only other group sharing this character, the longipes-group differs in leg armature. In summary, based on the characters shown in Table 3, group separation is uniquivocal for all taxa sharing leg armature type 1, even for some very closely related genera and groups, except for the monotypic curvarta-group, which is rather similar to the tregoubovi-group and the olsoni-group in the Table. However, separation is obvious when considering the proportional lengths of the urosomites in addition, showing relatively short genital double-somite and very long postgenital somites in O. curvata compared with the tregoubovi-group and the olsoni-group. Additional information is also required for grouping the new morphospecies Oncae sp. A-MedSea, for which the complete set of morphological characters has not yet been examined. The species appears to be unique by the nature and position of the genital apertures of the female, located extremely laterally on the genital double-somite. At present only one species of Triconia ( T. gonopleura) has been found to show a similar character, but the absence of a conical process on the distal endopod segment of P4 in sp. A-MedSea clearly points that the two species belong to different groups. The speciose genus Triconiaconifera- vs. similis-subgroup). However, a further subgrouping may be necessary, as it has been found by Böttger-Schnack & Machida (2011) that a minuta-subgroup can be split from the similis-subgroup by the proportional spine lengths on endopodite 3 of P2, and by genetic evidence. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.12, Table 3 (Continued)]. Morphological characters of oncaeid genera and of species groups within Oncaea s.l. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.13, Table 3 (Continued)]. Morphological characters of oncaeid genera and of species groups within Oncaea s.l. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.14, Table 4]. Leg armature of Oncaeidar typical leg armature (Type 1) found in the majority of species*. Nota: Two oncaeid genera, Oncaea s.str., and Triconia, eight species groups of Oncaea s.l. and one new morphospecies of uncertain taxonomic status ( Oncaea sp.A-MedSea) share leg armature type 1 (Table 3), i.e. the typical spine and setal count on the swimming legs (cf. Table 4). In total 73 species, corresponding to 2/3 of all described oncaeids and one of the new morphospecies, share this ''typical'' leg armature. For an unequivocal classification of these specues groups and genera, other morphological characters have to be considered in addition, most of which are presented in Table 3. Deviations from this typical leg armature are provided in Table 5 and 6, listing only those parts of the legs, where modified spine counts can be found. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.15, Table 5]. Leg armature of Oncaeidae variation of spine count on endopods and exopods (Type 2 to 8). |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.16, Table 5 (Continued)]. Leg armature of Oncaeidae variation of spine count on endopods and exopods (Type 2 to 8). |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.17, Table 6]. Leg armature of Oncaeidae modifications only on endopods (Type 10 to 14), only distal segments listed). |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.18, Table 6 (Continued)]. Leg armature of Oncaeidae modifications only on endopods (Type 10 to 14), only distal segments listed). |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.8, Table 1 C]. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.7, Table 1 B]. |
 Issued from : R. Böttger-Schnack & D. Schnack in J. Nat. Hist., 2013, (4-7). [p.8, Table 1 B]. | | | | | (1) Archioncaea Böttger-Schnack & Huys, 1997 | |
| | Ref.: | Böttger-Schnack & Huys, 1997 (p.80); Boxshall & Halsey, 2004 (p.613, 615); Böttger-Schnack & Schnack, 2013 Table 1, 5, (p.18, Rem. | | Rem.: | Total: 1 sp.
For Böttger-Schnack & Schnack (2013, p.18) the genus has a unique leg armature and combines primitive and advanced characters, which separates this taxon from all other known oncaeids. Most remarkable is the trisetose P5 exopod. On all four swimming legs, the middle and distal segments of the endopods are completely fused and the leg armature (Table 5, type 4) is characterized by the absence of an outer subdistal spine on the fused endopodal segments of P3 and P4 and showing only 2 outer spines on exopodite 3 of P2. | | | | | (2) Conaea Giesbrecht, 1891 | |
| | Syn.: | Conäa Giesbrecht, 1891; 1892 (p.82, 605) | | Ref.: | A. Scott, 1909 (p.243); van Breemen, 1908 a (p.191); Rose, 1933 a (p.303); Davis, 1949 (p.77); Heron, 1977 (p.38, 86, spp. Key); Boxshall, 1977 a (p.145); Razouls, 1982 (p.678); Malt, 1982 (p.191, 192, Rem.); Heron & al., 1984 (p.449); Razouls, 1993 (p.313); Heron & Bradford-Grieve, 1995 (p.45, Déf., Rem.); Huys & Böttger-Schnack, 1996/97 (p.245, 246, Rem.); Böttger-Schnack & Huys, 1998 (p.369); Bradford-Grieve & al., 1999 (p.970, spp. key); Heron & Frost, 2000 (p.1060, spp. key); Boxshall & Halsey, 2004 (p.613, 615); Vives & Shmeleva, 2010 (p.260, Rem.); Böttger-Schnack & Schnack, 2013 (p.19, Rem.) | | Rem.: | type: Conäa rapax Giesbrecht,1891. 4 spp.
For Böttger-Schnack & Schnack (2013, p.19) within Conaea s.L. two subgroups can be distinguished (Table 3): the type species of the genus, C. rapax, (defined as subgroup Conaea s.str.) differs from all other Conaea species (difined as expressa-subgroup) by a different endopodal spine count on P2 (Table 5, type 3), and is also separated by details of the lateral elements on enddopodite 2 of A2, especially the lack of the typical Conaea-like spinulose ornamentation of antennary elements and a very small seta IV. Further differences are found in the proportional lengths of the urosomites. | | Remarks on dimensions and sex ratio: | | The size ratio (male : female) is 0.886 (n = 3; SD = 0.0111) or 89 %, if we consider the ratio for each species. | | | | | (3) Epicalymma Heron, 1977 | |
| | Syn.: | Oncaea : Malt, 1982 (part., p.192) | | Ref.: | Heron, 1977 (p.82); Razouls, 1982 (p.682); Heron & al., 1984 (p.449, 479, spp. key); Razouls, 1993 (p.313); Heron & Bradford-Grieve, 1995 (p.45, Rem.); Huys & Böttger-Schnack, 1996/97 (p.245, 246, Rem.); Heron & Frost, 2000 (p.1058, spp. key); Boxshall & Halsey, 2004 (p.613, 615); Vives & Shmeleva, 2010 (p.262, Rem., spp. key); Böttger-Schnack & Schnack, 2013 (Table 1, 3, 5, p.19, Rem.) | | Rem.: | Malt (1982, p.185) regards this genus as a synonym of Oncaea, contrary to Heron & al. (1984) and Heron & Bradford-Grieve (1995).
Type: Epicalymma schmitti. 7 spp.:
For Böttger-Schnack & Schnack, (2013, p.19) within the genus, one species, E. umbonata, may deserve a separate group status, based on morphological grounds (labral structure, antennary and maxillipedal elements) as discussed by Böttger-Schnack (2009), but not shown in detail in the present study. hence, the species has been subsumed under Epicalymma S.l. (in Table 3, p.12) to denote the potential existence of two lineages within this genus. | | Remarks on dimensions and sex ratio: | | If we consider ratios species by species, the mean female size is 0,559 mm (n= 6; S= 0,105; Cv= 0,188) and the mean male size is 0,483 mm (n= 3; S= 0,097; Cv= 0,201). The size ratio (M/F) is 0,948 ou 94,8 % (n= 3; S= 0,161; Cv= 0,17).
The size ratio (male : female) is 0.926 (n = 5; SD = 0.1173), if we consider the ratio for each species. | | | | | (4) Monothula Böttger-Schnack & Huys, 2001 | |
| | Ref.: | Böttger-Schnack & Huys, 2001 (p.468); Boxshall & Halsey, 2004 (p.613, 615); Vives & Shmeleva, 2010 (p.266, Rem..); Böttger-Schnack & Schnack, 2013 (Table 1, 3, 6, p.19, Rem.) | | Rem.: | type: Oncaea subtilis Giesbrecht,1892. Total: 1 sp. | | | | | Myspictosum Kazatchenko & Andreev, 1977 | | Ref.: | Razouls, 1982 (p.683); Malt, 1982 (p.192, Rem.) | | Rem.: | Cf. Oncaea | | | | (5) Oncaea Philippi, 1843 | |
| | Syn.: | Antaria Dana, 1846; Conäa Giesbrecht, 1891; Oncäa : Giesbrecht, 1892 (p.81, 590); Myspictosum Kazatchenko & Andreev, 1977 (p.47) | | Ref.: | Sars, 1900 (part., p.106); Wheeler, 1901 (p.190); Esterly, 1905 (p.215); van Breemen, 1908 a (p.184); Farran, 1908 b (p.95, spp. key of Females); A. Scott, 1909 (p.241); Sars, 1918 (p.190); Pesta, 1920 (p.650 spp. key); Wilson, 1932 a (p.350, spp. key); Rose, 1933 a (p.295, spp. nkey of Females); Mori, 1937 (1964) (p.119); Dakin & Colefax, 1940 (p.116, spp. key); Davis, 1949 (p.76); Shmeleva, 1968 (p.1791, clé spp.); 1969 (p.3, 25, spp. key); Ferrari, 1975 (p.217, 230, clé spp.); Heron, 1977 (p.40, spp. key); Razouls, 1982 (p.665); Gardner & Szabo, 1982 (p.113); Malt, 1982 (p.192, Rev.); 1982 a (p.140, key. N, juv.); 1983 a (p.4, spp. key); 1983 b (p.449, polymorphism); Heron & al., 1984 (p.461, clé spp., Rem.); Zheng Zhong & al., 1984 (1989) (p.262, spp. key); Böttger-Schnack & al., 1989 (p.1089, Rem.: biology); Huys & Boxshall, 1991 (p.465); Heron & Bradford-Grieve, 1995 (p.13, Déf., spp. key of Females, Rem.); Chihara & Murano, 1997 (p.979, Pl.221); Böttger-Schnack & Huys, 1998 (p.369, Rem.: phylogeny); Bradford-Grieve & al., 1999 (p.970, spp. key); Heron & Frost, 2000 (p.1017, spp. key, Rem.: For the authors the genus Triconia is questionable); Böttger-Schnack, 2001 (p.28, Déf.); Boxshall & Halsey, 2004 (p.613, 615); Vives & Shmeleva, 2010 (p.269, Rem., spp. key from Iberica Fauna); Böttger-Schnack & Machida, 1011 (p.111, DNA sequences, phylogeny); Böttger-Schnack, 2013 (Table 1: sub)groups, Table 3, 5, 6: subgroups identifications, p.18: Rem.) . | | Rem.: | Malt (1982, p.191, 192) regards the 2 genera Conaea and Epicalymma as synonyms of Oncaea, whereas Heron & al. (1984) and Heron & Bradford-Grieve (1995) believe that they should be maintained.
Type: Oncaea venusta Philippi,1843. Total: 71 spp. + 1 unidentified.:
After Ohtsuka & Nishida (2017, p.582), Oncaea abd related genera are associated with discarded appendicularian houses, feeding on plankters retained on the appendicularian feeding nets (Ohtsuka & Kubo, 1991; Ohtsuka & &l., 1993; 1996a). | | | | | (6) Spinoncaea Böttger-Schnack, 2003 | |
| | Ref.: | Böttger-Schnack, 2003 (p.189, Def.); Boxshall & Halsey, 2004 (p.613, 615); Huys & Böttger-Schnack, 2007 (p.121, Table 3); Vives & Shmeleva, 2010 (p.328, Rem., spp. key) | | Rem.: | type: Oncaea ivlevi. Total: 3 spp.
For Böttger-Schnack & Schnack (2013, p.19) the genus displays intrageneric variation in the spine count on the exopod segment of P2 (Table 5, type 6). Similar variation has also been found in the longipes-group and the zernovi -group (Table 5: type 5 and type 7a, respectively). In allthese cases, however, all the other morphological characters of the species, including details of ornamentation, were similar (unlike the situation within Conaea s.l.), indicating that they are closelt related sibling/sister species within the same group. Hence, differences in exopodal spine counts on the distal segment of P2 alone are not regaeded as an indication for defining separate subgroups within any group defined in the present study. | | Remarks on dimensions and sex ratio: | | The size ratio (male : female) is 0.968 (n = 3; SD = 0.0418) if we consider the ratio for each species. | | | | | (7) Triconia Böttger-Schnack, 1999 | |
| | Ref.: | Böttger-Schnack, 1999 (p.41, Def.); Boxshall & Halsey, 2004 (p.613, 615); Vives & Shmeleva, 2010 (p.336, Rem., spp. key from Iberica Fauna); Böttger-Schnack & Machida, 1011 (p.111, DNA sequences, phylogeny); Böttger-Schnack & Schnack, 2013 (table 1, 3, 4, identification, p.1, 186: Rem.) | | Rem.: | type: Oncaea conifera Giesbrecht,1891. Total: 26 spp. + 1 indet.
Generic name derived from the Latin tres, tria, meaning three and conus, meaning cone, and refers to the presence of a conical projection on the distal endopodal segments of the last three pairs of swimming legs.
Böttger-Schnack (1999) established the genus Triconia which is composed of all oncaeids having a large conical process on the distal margin of the endopod of swimming legs 2-4.
This genus was subdivided into 3 subgroups: similis, conifera, dendipes.
After reexamination, the similis sub-group includes 2 groups, which can be distinguished by the proportional spine lengths on the endopods of swimming P2: 1- containingT. minuta, T. umerus, T. gonopleura, T. parasimilis, present a relatively short distal endopodal spine: 2- including T. similis, T. hawii, T. recta, T. denticulata, with a relatively long distal endopodal spine. | | | | | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 22, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
