|
|
 |
|
Calanoida ( Order ) |
|
|
|
Arietelloidea ( Superfamily ) |
|
|
| |
| | | |
| Arietellidae Sars, 1902 ( Arietelloidea ) | | Ref.: | Sars, 1902 (1903) (p.123); Gurney, 1931 (p.85); Wilson, 1932 a (p.136); Rose, 1933 a (p.238); Brodsky, 1950 (1967) (p.393); Bradford, 1969 b (p.502, Table 3); Andronov, 1974 a (p.1005); Campaner, 1977 (p.811, Redef.); Björnberg & al., 1981 (p.650); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.526); Brodsky & al., 1983 (p.143, 145); Campaner, 1984 (p.102, 107, Rem.); Mauchline, 1988 (p.705, 706, 740: cuticular pores); Ohtsuka & Mitsuzumi, 1990 (p.96); Huys & Boxshall, 1991 (p.460); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.105, Redef., diagnosis emended, Genera Key, cladistic analysis); Chihara & Murano, 1997 (p.719); Cuoc & al., 1997 (p.651, phylogeny); Bradford-Grieve & al., 1999 (p.881, 902, 903, 936, 937: Genera Key); Bradford-Grieve, 1999 b (p.27, Def.); Ohtsuka & Huys, 2001 (p.445, 461); Boxshall & Halsey, 2004 (p.13; 49; 62: Def.; p.64: Genera Key); Vives & Shmeleva, 2007 (p.146, part. Genera Key); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, Biol. mol, phylogeny); Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | Definition from Bradford-Grieve (1999 b, p.27) :
Female :
- Body of variable size (0.8-7 mm), relatively robust.
- head and 1st pediger separated or weakly fused, 4th and 5th pedigers fused, sharply or weakly produced, with or without dorsolateral and/or ventrolatetal processes
- Forehead rounded or pointed anteriorly.
- Rostrum produced ventrally with a pair of filaments.
- Urosome relatively short, 4-segmented.
- Genital segment with single paired gonopores and copulatory pores; gonopore(s) located ventrolaterally or ventrally, with or without opercular plate; copulatory pore sharing a common opening with the gonopore or separate with the gonopore located ventromedially or posteriorly, rarely ventrally on the right side; seminal receptacles usually paired, rarely the left is entirely lacking;
- Caudal rami symmetrical or slightly asymmetrical, relatively short with up to 7 setae.
- A1 symmetrical or asymmetrical, longer on left that on right, 16-22 segments, sometimes differing in fusion pattern and armature.
- A2 - basipod 2 (= basis) with an outer edge seta; exopod and endopod separated from basis; basis 2-segmented, segment 1 with 0-1 seta at mid-length, segment 2 with 1-3 inner setae medially and 5 or 6 setae terminally; endopod indistinctly 6-10-segmented.
- Md blade well chitinised, with 3 or 4 sharp teeth; endopod rudimentary, 1-segmented with 1 or 2 setae terminally or completely absent; exopod segment 1 with normal or reduced setae, segment 5 carrying 2 setae, one of which is sometimes vestigial.
- Mx1 inner lobe 1 (arthrite) with 0-6 spines and setae; inner lobe 2 with 1 seta or unarmed; inner lobe 3 represented by 1 seta which may be vestigial or absent; endopod bulbous, 1-segmented, with 0-3 setae or completely incorporated into basis; exopode lobate, with 3 setae; outer lobe 1 with 5-9 setae.
- Mx2 well-developed; inner lobe 1 with 1-3 setae (1 vestigial), inner lobe 2 with 1-2 setae; inner lobes 3 and 4 with 2 setae each, inner lobe 5 with a stout spine, spinulose or bare; endopod 4-segmented with 1, 3, 2, 2 setae respectively.
- Mxp elongate, basipod 1 (:= coxa) with 1 medial and 2 terminal setae; basis with patches of setules or spinules and 2 setae medially; endopod 6-segmented, segment 1 almost fully incorporated into basis with 1, 4, 4, 3 (rarely 2), 3 (rarely 2), 4, segment 6 with 2 outermost terminal setae reduced or not.
- Swimming legs 1-4 distinctly 3-segmented, rarely with endopod segments of P1 incompletely fused. Sta and spine formula as in table).
- P5 variable, not natatory, almos symmetrical; coxa and coupler (sclerite) separate or fused; basis end endopod separate or fused; endopod with 0-4 setae; exopod 1-3-segmented or completely fused with basis, carrying 0-5 spines and setae.
Male :
- Body similar to female.
- Urosome 5-segmented.
- Left A1 geniculate, 16-20-segmented, proximal segments often with a row of setules along posterior margins.
- Mouthparts and P1-P4 similar to female or with slightly different armature elements of A2, endopod segment 2 and mandibular exopod segment 1.
- P5 variable, not natatory, almost symmetrical to strongly asymmetrical; coxa and coupler (sclerite) fused to form a common base or separate; right basis sometimes fused with coxa; right endopod 1-segmented, bulbous or absent; right exopod distinctly or indistinctly 3-segmented, segment 1 and 2 each with 1 seta on the outer margin (rarely segment 1 unarmed), segment 2 with a tuft of setules on inner distal angle of segment 2, segment 3 with 0-3 spines and setae terminally; left endopod 1-2-segmented, unarmed or completely absent; left exopod distinctly 3-segmented, segments 1 and 2 each with 1 seta on the outer margin, segment 3 with 1-3 spines and setae terminally.
Ohtsuka, Boxshall & Roe (1994) redefine the family and establish the phylogenetic relationships between the genera (including 3 new). Type-genus: Arietellus Giesbrecht, 1893 ['1892'']. Now a total of 12 genera: Arietellus, Campaneria, Crassarietellus, Metacalanalis, Metacalanus, Paramisophria, Paraugaptiloides, Paraugaptilus, Pilarella, Protoparamisophria, Sarsarietellus, Scutogerulus. Griceus is considered as incertae sedis and Rhapidophorus as inquirendum. These authors do not retain the sub-families but show the existence of 2 lines.
Taxonomic notes in Boxshall & Halsey (2004, p.64): The Arietellidae was placed in the superfamily Arietelloidea (as Augaptiloidea) by Andronov (1974) and Park (1986). Huys & Boxshall (1991) regarded it as one of the most plesiomorphic of calanoid families, on the basis of the morphology of the female genital system as well as on limb segmentation and setation. The female genital system is highly variable in the Arietellidae but paired gonopores and paired copulatory pores are found within the family. Some arietellids exhibit a tendency towards asymmetry related to their habit of swimming close to the bottom on one side (see Fosshagen, 1968b) or lying on the bottom on one side (Ohtsuka & Mitsuzumi, 1990). This asymmetry is reflected particularly in the female reproductive system and in the length and form of the female antennules.
Key to genera (adapted from Ohtsuka, Boxshall & Roe, 1994) :
1 4 distal setae present on exopod of Mx1 ; rostrum bilobed, plate-like, without paired rostral filaments
Griceus.
1 3 setae present on exopod of Mx1 ; rostrum with paired rostral filaments
. 2.
2 P1 with 1 outer spine on 3rd exopodal segment
. 3.
2 P1 with 2 outer spines on 3rd exopodal segment
.. 5 .
3 - Mx1 with 5 spines and 1 process on praecoxal arthrite ; caudal seta II well developed ; female genital double-somite with paired genital system, each copilatory pore opening inside common genital slit shared with gonopore
4.
3 Mx1 with 0/2 elements on praecoxal arthrite ; caudal seta II reduced ; female genital double-somite with separate copulatory pore(s) located anteromedially to gonopore(s)
Metacalanus.
4 Mx2 with 1 seta on distal praecoxal endite ; female A1 22-segmented
Scutogerulus.
4 Mx2 with 2 setae on distal praecoxal endite ; female A1 21-segmented
Pilarella.
5 Mx1 with 6 setae on coxal epipodite
6.
5 Mx1 with 8 or 9 setae on coxal epipodite
7.
6 Mxp with long proximal seta on 5th endopodal segment and well developed outermost seta on 6th segment ; male left A1 with segments XXI and XXII fused ; endopod of male right P5 lacking
Crassarietellus.
6 Mxp with short proximal seta on 5th endopodal segment and reduced outermost seta on 6th segment ; male left A1 with segments XXI and XXII separate ; endopod of maleright P5 present
Campaneria.
7 Proximal seta on 4thand 5th segments of Mxp vestigial
. Arietellus.
7 Proximal seta on 4th and 5th segments of Mxp normally developed
8.
8 Apical segment of A2 exopod unarmed
Paraugaptilus
.Apical segment of A2 exopod with 3 setae
. 9.
9 P4 with inner coxal seta ; 2nd endopodal segment of A2 with 2 inner setae at midlength
.. Paraugaptiloides9 P4 without inner coxal seta ; 2nd endopodal segment of A2 with 3 inner setae at midlength
10.
10 - A1 segments XXV and XXVI separated ; Mxp with vestigial outermost seta on 6th endopodal segment ; inner (endopodal) process of female P5 with 4 setae
. Sarsarietellus.
10 - A1 segments XXV and XXVI fused ; Mxp with normally developed outermost seta on 6th endopodal segment ; inner (endopodal) process of female P5 with 0/2 setae
Paramisophria. | 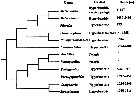 issued from : S. Ohtsuka, S. Nishida & R.J. Machida in J. Nat. Hist., 2005, 39 (27). [p.2510, Fig.14]. Cladogram of Arietellid genera. Main habitat and depth range provided only for hyperbenthic genera, exvluding two truly pelagic genera, Arietellus and Paraugaptilus. The phylogenetic relationship among arietellid genera two clades are recognized: the Metacalanus-group and the -group. |
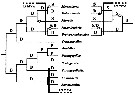 issued from : S. Ohtsuka, S. Nishida & R.J. Machida in J. Nat. Hist., 2005, 39 (27). [p.2512, Fig.15]. Cladogram depicting main habitats and colonization routes for arietellid genera and their presumed ancestrors. D: deep-water hyperbenthic: S: shallow-water hyperbenthic (including cavernicolous and epipelagic); P: deep-water planktonic. In the Metacalanus-Metacalanis-Pilarella-clade there are two possibilities for colonization routes (right: habitat transition from deep-sea to shallow-water hyperbenthos; left: habitat shift from deep-sea to shallow-water hyperbenthos occurred independently in Metacalanus and Pilarella. (o) Habitat shift from deep-sae hyperbenthic to shallow-sea hyperbenthos; (black o) habitat shift from shallow-water hyperbenthos to deep-sea hyperbenthos. |
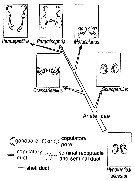
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.161, Fig.38]. Evolutionary trends in the structures of the female genital systems of the arietellid genera. A: Fusion of copulatory pores to form single pore, and anterolateral migration of both gonopores. B: Posterior migration of both gonopores and copulatory pores, and separation of copulatory pore from gonopore. C: Anterolateral migration of gonopores, and separation of copulatory pore from gonopore and their asymmetrical arrangement and enlargement. D: Lateral migration of both gonopores and copulatory pores, and separation of copulatory pore from gonopore. E: Lateral migration of both gonopores and copulatory pores. Pg: Paramisophria giselae; Pj: Paramisophria japonica; Pc: Paramisophria cluthae; Sa: Sarsarietellus abyssalis; Ap: Arietellus plumifer; M1: Metacalanus sp.1; M2: Metacalanus sp.2; Ps: Paraugaptilus similis; Ch: Crassarietellus huysi; Sp: Scutogerulus pelophilus. g : gonopore; c: copulatory pore. |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.163, Fig.39]. Schematic illustration of segmental fusion patterns and armature of female antennules of the arietellid genera. A: Crassarietellus; B: Sarsarietellus; C: Scutogerulus; D: Paramisophria; E: Arietellus; F: Paraugaptilus; G: Metacalanus; H: Pilarella. The number indicates the number of setae on each segment. Asterisk indicates segment without aesthetasc. |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.164, Fig.40]. Schematic illustration of segmental fusion patterns and armature of male antennules of the arietellid genera. A: Campaneria; B: Crassarietellus; C: Paramisophria; D: Arietellus; E: Paraugaptiloides; F: Paraugaptilus; G: Metacalanus. The number indicates the number of setae on each segment. Asterisk indicates a segment with no aesthetasc. |
 issued from :C. Cuoc, D. Defaye, M. Brunet, R. Notonier & J. Mazza in Mar. Biol., 1997, 129. [p.13, Fig.6]. Diagrammatic representation of main evolutionary trends in organization of female genital area in calanoïds for Arietellidae (modified after Huys & Boxshall, 1991 and Ohtsuka & al., 1994). |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.165, Fig.41]. Schematic illustration of fusion patterns and armature of antennary exopods of the arietellid genera. A, Hypothetical calanoid ancestor; B, Crassarietellus; C, Paramisophria giselae; D, Campaneria, Paraugaptiloides, Paramisophria japonica, Sarsarietellus, Scutogerulus; E, Metacalanus; F, Arietellus; G, Paraugaptilus similis, male; H, P. similis, female. Solid and dotted lines indicating complete separation between segments, and incomplete fusion or suture between segments, respectively. |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.168, Fig.42]. Schematic comparison of patterns of segmentation and setation of female P5 in some arietellids. The arrows indicate possible derivation of setation and segmentation patterns and are not indicative of ancestor-descendant relationships between taxa. Ap: Aeietellus pavoninus; As: Arietellus sp.; Am: Arietellus mohri; Ps: Paraugaptilus similis; Sa: Sarsarietellus abyssalis; Pj: Paramisophria japonica; Pi: Paramisophria itoi; Pp: Paramisophria platysoma; Pg: Paramisophria giselae; Pr: Paramisophria reducta; Ch: Crassarietellus huysi; M1: Metacalanus sp.1; M2: Metacalanus sp.2; Ma: Metacalanus acutioperculum; Pl: Pilarella longicornis. C = coxa; B (in C. huysi) = basis; Is = intercoxal sclerite; Ex = exopod; En = endopod. A-D (in S. abyssalis): setae on endopod; a-f: spines on exopod. |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.169, Fig.43]. Schematic comparison of segmentation and setation of male P5 in the Arietellidae. The arrows indicate possible derivations of setation and segmentation patterns and are not indicative of ancestor-descendant relationships between taxa. Ch: Crassarietellus huysi; Pp: Paramisophria platysoma; Pm: Paraugaptiloides magnus; Cl: Campaneria latipes; M1: Metacalanus sp.1; Ap: Arietellus plumifer; Pb: Paraugaptilus buchani. C = coxa; B = basis; Is = intercoxal sclerite; Ex = exopod; En = endopod. a-f, k: elements on exopod. Setae and spines are not distinguished here. |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.170]. Key to genera of the family Arietellidae (first part.) |
 issued from : S. Ohtsuka, G.A. Boxshall & H.S.J. Roe in Bull. nat. Hist. Mus. Lond. (Zool.), 1994, 60 (2). [p.171]. Key to genera of the family Arietellidae (continued second part.) |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.62]. Armature of legs on the pedigerous somites. Nota: Swimming legs P1 to P4, with 3-segmented rami, except, very rarely, with endopodal segments of P1 incompletely fused. Inner seta on basis of P1 situated on anterior surface of basis and passing across face of endopod. Serarion sometimes reduced. - Female P5 variable but never natatory; almost symmetrical; coxae and intercoxal sclerite separate or fused; basis and endopod separate or fused; endopod primitively 1-segmented with up to 4 setae; exopod 1 to 3-segmented ot completely fused to basis, bearing 0 to 5 outer and terminal elements (setation formula above given for Sarsarietellus abyssalis Sars. - Male P5 variable, from almost symmetrical to strongly asymmetrical; coxae and intercoxal sclerite separate or fused to form common base. Right leg with basis sometimes fused to coxa; endopod 1-segmented lobate or absent; exopod distinctly or indistinctly 3-qzgmented; 1st segment typically with outer margin element, 2nd segment with outer element and inner patch of setules, 3rd segment with 0 to 3 elements. Left leg endopod 1 or 2-segmented, unarmed or completely absent; exopod distinctly or indistinctly 3-segmented, 1st and 2nd segments each with outer element, 3rd segment with 1 to 3 element terminally. - Eggs contained in paired, multiseriate sacs. |
 Issued from : J.M. Bradford-Grieve in NIWA Biodiv. Memoir 111, 1999. [p.27]. Armature of the swimming legs P1 to P4. (spines: Roman numerals; setae (Arabic numerals). | | | | | Arietellinae Brodsky, 1950 | | Ref.: | Brodsky, 1950 (1967) (p.394); Brodsky & al., 1983 (p.143) | | Rem.: | Brodsky crée cette sous-famille pour y inclure Arietellus, Paraugaptilus, Paramisophria et Scottula et considère Phyllopus comme le type d'une autre sous-famille (Phyllopinae ). Campaner (1977, p.811) considère celle-ci comme une famille : Phyllopidae (emend. Bowman & Abele, 1982, p.9 = Phyllopodidae = Nullosetigeridae ). Dans ces conditions la sous-famille établie par Brodsky n'a plus de raison d'être. | | | | (1) Arietellus Giesbrecht, 1892 | |
| | Ref.: | Giesbrecht, 1892 (p.66, 415); Giesbrecht & Schmeil, 1898 (p.124); Esterly, 1905 (p.189); Farran, 1908 b (p.81); Van Breemen, 1908 a (p.139, 238, 239); A. Scott, 1909 (p.142); Wolfenden, 1911 (p.329); Sewell, 1932 (p.329); Rose, 1933 a (p.238, spp. Key); Tanaka, 1964 c (p.231); Owre & Foyo, 1967 (p.90, spp. Key); Brodsky, 1950 (1967) (p.394, spp. Key); Razouls, 1982 (p.526); Gardner & Szabo, 1982 (p.395, Rem.); Mauchline, 1988 (p.705, 740); Razouls,1993 (p.306); Ohtsuka & al., 1994 (p.126, Redef.); Chihara Murano, 1997 (p.720); Mauchline, 1998 (p.71: F; p.74: M); Bradford-Grieve & al., 1999 (p.937: spp. key); Bradford-Grieve,1999 b (p.28, Def.); Boxshall & Halsey, 2004 (p.65); Vives & Shmeleva, 2007 (p.147, spp. key) | | Rem.: | type: Arietellus setosus Giesbrecht, 1893 [1892]. Total: 15 spp. + 1 undetermined.
Definition after Bradford-Grieve (1999 b, p.28) :
- As in the family with the additional characters:
- Body relatively robust.
- Cephalosome and pediger 1 separated or weakly fused, pedigers 4 and 5 fused.
- Forehead rounded or pointed anteriorly
- Posterior metasome with a pair of blunt or pointed lateral processes symmetrical of asymmetrical, strongly or weakly produced backwards.
- Rostrum produced ventrally with a pair of thin filaments.
Urosome female relatively short, 4-segmented.
- Urosome male 5-segmented.
- Genital segment female longer than wide ; a pair of gonopores ventrolaterally and copulatory pore ventromedially ; seminal receptacle relatively large, bulbous, located laterally.
- Anal segment large, operculum not developed.
- Caudal rami symmetrical, longer than wide, divergent or not, with all 6 terminal setae well developed.
- A1 female symmetrical, distinctly 20-segmented ; posterior margins of segments 1-10 fringed with long setules.
- Left A1 male 19-segmented, geniculate (segments 21 and 22 fused).
- A2: Endopodal segment 2 unarmed (in male with 1 long and 1 short seta medially), segment 2 with 2 inner setae (reduced in some species), and 5 setae and a setule terminally; exopod indistinctly 7- or 8-segmented.
- Md: blade lacking a tuft of hairs at midlength; 3 cusped teeth on cutting edge, dorsalmost of which is bifid at tip; mandibular palp with endopod absent; exopod segments of female with reduced or normal seta, outer seta of segment 5 vestigial but in the male exopod segment 1 has normally developed setae.
- Mx1: inner lobe 1 with 5 spines and 1 process; inner lobe 2 with 1 relatively short, thick seta fringed with long setules; endopod rudimentary, almost fused to basipod 2 or 1-segmented, bulbous, with 1 terminal seta;
- P5 female reduced ; basipod 1 and the plate between basipod 1 on each side fused to form a common transverse plate ; basipod 1 and 2 fused or separate ; right basipod 2 longer than left basipod 2 ; endopod fused with basipod 2, represented by a small knob with 1-3 setae terminally, vestigial in some species ; exopod 1-segmented, bulbous, carrying 1 terminal spine or almost fused to bnasipod 2, unarmed.
- P5 male with basipod 1 and coupler form a common plate. Right basipods 1 and 2 incompletely fused ; basipod 2 inner seta remarkably or normally elongate ; endopod 1-segmented and unarmed ; exopod indistinctly 3-segmented with distal 2 segments incompletely fused ; exopod segment 2 with a stout process on the inner angle ; exopod segment 3 spatulate, with 0-2 vestigial elements. Left endopod indistinctly 2-segmented or 1-segme,ted, unarmed ; exopod 3-segmented, segment 2 expanded medially, segment 3 incompletely fused with the preceding segment, bearing 2 terminal spines, with or without an outer minute spinules. | | Remarks on dimensions and sex ratio: | | The mean female size is 5.076 mm (n = 25; SD = 0.9935), and for the males 4.827 (n = 16; SD = 0.8135). The size ratio (Male: Female) is 0,951. The sex ratio (F:M) is 1.555. | | | | | (2) Campaneria Ohtsuka, Boxshall & Roe, 1994 | |
| | Ref.: | Ohtsuka & al., 1994 (p.119); Mauchline, 1998 (p.74: M); Bradford-Grieve,1999 b (p.34, Def., Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.64) | | Rem.: | Type: Campaneria latipes Ohtsuka, Boxshall & Roe, 1994. 1 sp.99
Definition after Bradford-Grieve (1999 b, p.34) :
Male:
- As in the family definition.
- Anal segment short, operculum not developed.
- Caudal rami symmetrical, longer than wide, outer lateral and posterodorsal seta minute.
- left A1 male reaching almost to end of urosome, 20-segmented.
- A2 endopodal segment 1 with 1 inner seta; segment 2 with 3 inner subterminal setae and 5 terminal setae; exopod indistinctly 8-segmented.
- Md blade with a tuft of setules, palp with rudimentary, 1-segmented endopod with 2 setae, exopodal segment 1 seta not reduced, outer seta on exopodal segment 5 relatively long.
- Mx1 inner lobe 1 (= arthrite) with 5 spines (3 of which weakly serrate medially) and a process; inner lobe 2 with 1 long seta; basis with 1 vestigial seta; endopod bulbous, 1-segmented with 2 setae; exopod with 3 setae; outer lobe 1 with 6 setae;
- Mx2 lobes 1-4 with 2 setae each, lobe 5 with 1 stout spine with 3 rows of spinules proximally.
- Mxp endopodal setal formula 1, 4, 4, 3, 3, 4; endopodal segment 4 with a non-reduced innermost seta, segment 5 with innermost seta shorter than segment 4, segment 6 with 2 large setae, 1 small seta and 1 vestigial seta.
- P1 with 2 outer spines on exopodal segment 3;
- P4 lacking an inner seta on coxa.
- P5 coxa and the common plate between them fused; coxa and basis separate; right endopod 1-segmented, bulbous, exopod indistinctly 3-segmented, distal 2 segments almost fused, expanded medially, with a rounded process medially and 2 setules and 1 prominence terminally; left endopod indistinctly 2-segmented, unarmed. Exopod 2-segmented, distal segment curved outwards near its tip, with 3 setae terminally and 1 seta medially.
- Female unknown.
For Bradford-Grieve, this genus is based on the male described as Scutogerulus pelophilus Bradford, 1969. It was separated by Ohtsuka & al., 1994) because the male does not have shield-shaped appendages on terminal setae of Mx2 and Mxp and other characters. | | | | | (3) Crassarietellus Ohtsuka, Boxshall & Roe, 1994 | |
| | Ref.: | Ohtsuka & al., 1994 (p.109); Mauchline, 1998 (p.71: F); Bradford-Grieve,1999 b (p.34, Def.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.64) | | Rem.: | type: Crassarietellus huysi. 1 sp. + 1 indet. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.865 mm (n =2; SD = 0.021), and for males 2.84 mm (n = 2; SD 0.0056). The size ratio (male: female) is 0.735, but the species male does not correspond to the female. | | | | (0) Griceus Ferrari & Markhaseva, 2000 | |
| | Ref.: | Ferrari & Markhaseva, 2000 (p.77); Bradford-Grieve, 2004 (p.287) | | Rem.: | After Boxshall & Halsey, 2004 (p.64, Rem.: Genus incertae sedis). 1 sp. | | | | (4) Metacalanalis Ohtsuka, Nishida & Machida, 2005 | |
| | Ref.: | Ohtsuka & al., 2005 (p.2484 | | Rem.: | Type: Metacalanalis hakuhoae. 1 sp.
For Ohtsuka & al. (2005, p.2485) the genus is similar to the genera Metacalanus Cleve, 1901 and Pilarella Alvarez, 1985, and has mixed features of these two genera. Metacalanis is distinguishable from Metacalanus and Pilarella by the following features: (1) caudal rami about 1.2 times as long as wide; (2) left A1 reaching at most to the posterior margin of pediger 3: (3) A2 showing the most primitive condition in the family Arietellidae, with an indistinctly 3-segmented endopod and an indistinctly 10-segmented exopod with setal formula 0, 0, 1, 1, 1, 1, 1, 0, 2; (4) praecoxal arthrite of Mx1 with 1 short and 4 long spines; (5) coxal epipodite of Mx1 with 6 setae; (6) basal spine of Mx2 with a short row of minute spinules medially; (7) P5 female with a unisegmented exopod bearing 2 lateral and 2 terminal spines. Asymmetry of P1-P3 is unique to the new genus. | | | | (5) Metacalanus Cleve, 1901 | |
| | Syn.: | Scottula (part.) Sars, 1902 | | Ref.: | Cleve, 1901 (p.43); A. Scott, 1909 (p.146); Sewell, 1932 (p.329); Tanaka, 1964 c (p.236); Razouls, 1982 (p.531); Alvarez, 1985 (p.190, 194, Rem.); Ohtsuka, 1985 (p.305); Mauchline, 1988 (p.706); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.138, Redef., diagnosis emended); Chihara & Murano, 1997 (p.719); Mauchline, 1998 (p.73: F; p.74: M); Bradford-Grieve,1999 b (p.35, Déf.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.64); Vives & Shmeleva, 2007 (p.155) | | Rem.: | type: Metacalanus aurivilli . 4 spp.+ 2 unidentified.
Definition after Bradford-Grieve (1999 b, p.35) :
- As in the family with the additional characters:
- Body small, compact.
- Prosome oval in dorsal view, not produced frontally.
- Cephalosome and pediger segment 1 separate or weakly fused.
- Posterior corners of last thoracic segment produced to form a ventrolateral lobe, without dorsolateral processes.
- Urosome short, less than 193 the length the length of prosome.
- Genital segment wider than long, with a ventrolatrtal pair of gonopores or only yje right gonopore (left reduced) located posteriorly ; paired copulatory pores small, located near the inner corner of genital aperture (in the case of the reduction of the left gonopore, only the right copulatory pore is present).
- Anal operculum either developed, triangular or not.
- Caudal rami symmetrical, longer than wide, with the dorsal lateral seta reduced or lacking ; the outer distal seta relatively small.
- Female A1 asymmetrical, left longer than right, different in fusion pattern and armature.
- Male left A1 16-segmented.
- A2 endopodal segment 1 with 1 inner seta, segment 2 with 2 setae medially, and 5 setae terminally ; exopod indistinctly 7-segmented.
- Mandibular blade lacking a tuft of setulrs, 4 teeth on cutting edge, dorsalmost of which is trifid at its tip ; palp with endopod almost fused with basipod 2 represented by a small knob with 1 or 2 setae terminally ; exopod segment 1 not reduced, segment 5 with outer setae relatively long.
- Mx1 inner lobe 1 with 0-2 spines ; endopod absent or 1-segmented, bulbous with 1 seta ; outer lobe 1 with 5 setae.
- Mx2 lobe 1 with 1 seta and 1 rudimentory element ; lobe 5 spine with 2 rows of minute spinules proximally ; endopod setae with a row of spinules along inner margin.
- Mxp setal formula on endopod segments 1-6 : 1, 4, 4, 3, 3, 4 ; the innerlost seta on endopod segments 4 and 5 not reduced ; only the distal most setae on these segments long.
- P1 exopod segment 3 with 1 outer spine.
- Female P5 basipod 1 separated from the common connecting plate ; endopod represented by 1 seta or completely absent ; exopod and basipod 2 frused or separate ; exopod either 1-segmented with 1-3 spines or represented by a small knob bearing 1 seta.
- Male P5 basipod 1 and common plate fused ; basipods 1 and 2 separate ; endopod absent ; exopod 3-segmented, segment 3 with a large seta almost fused with the segment.
After Ohtsuka & al. (1994, p.138) the species seems to be epipelagic in subtropical waters in the Indo-West Pacific, some are hyperbenthic in shallow waters in temperate and subtropical regions, or marine cave-dwellers. | | Remarks on dimensions and sex ratio: | | The mean female size is 0,871 mm (n = 10; SD = 0.2153), and for the males 0.754 mm (n = 7; SD = 0.1573). The size ratio (Male: Female) is 0,866. | | | | | (6) Paramisophria T. Scott, 1897 | |
| | Syn.: | Parapseudocyclops Campaner,1977 (p.812) | | Ref.: | T. Scott, 1897 f (p.147); Giesbrecht & Schmeil,1898 (p.123); Sars, 1902 (1903) (p.127); van Breemen, 1908 a (p.140); Rose, 1933 a (p.242); Brodsky, 1950 (1967) (p.394); Krishnaswamy, 1959 (p.546, Rem.: emend.); Tanaka, 1964 c (p.242); Razouls, 1982 (p.530); McKinnon & Kimmerer, 1985 (p.85, spp. Key); Ohtsuka, 1985 (p.296, 298, Rem.); Ohtsuka & Mitsuzumi, 1990 (p.97); Huys & Boxshall, 1991 (p.49, 325, 436); Ohtsuka & al.,1991 (p.793, 800, Rev.); Ohtsuka & al., 1993 (p.57, 65, Rem.: emend.); Ohtsuka & al., 1994 (p.158, figs., Rem.); Chihara & Murano, 1997 (p.719); Mauchline, 1998 (p.68); Bradford-Grieve,1999 b (p.36, Def.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.65); Vives & Shmeleva, 2007 (p.156, spp. Key); Lim & Min, 2014 (p.539, morphological phylogeny). | | Rem.: | type: Paramisophria cluthae . Lim & Min (2014, p.524) report 19 species world-wide distribution in temperate and tropical waters. 20 spp. + 2 unidentified.
Definition after Bradord-Grieve (1999, p.36) :
As in the family with the additional characters :
- head and pediger 1 separate, pedigers 4 and 5 fused, with or without dorsolateral processes.
- Urosome 4-segmented in female.
- Urosome 5-segmented in male.
- Female genital segment with a pair of anterolateral gonopores and a single ventromedial or ventrolateral copulatory pore.
- Anal operculum not well developed , nor pointed posteriorly.
- Outer proximal caudal setae vestigial, remaining setae well developed.
- A1 female 20-22-segmented, extending at most to distal border of cephalosome ; left A1 longer than right.
- Left male A1 geniculate, 19-21-segmented.
- A2 exopod indistinctly 8-9-segmented ; endopod 2-segmented, segment 1 with an inner medial seta, segment 2 with 3 inner setae at midlength and 5 setae and 1 minute seta terminally.
- Mandibular blade lacking or having a small tuft of setules medially ; with 3 teeth, the dorsalmost of which is bifid at the tip ; endopod fused with basipod 2 or consisting of 1 rudimentary segment ; exopod segment 1 with a well-developed seta, outer seta on segment 5 relatiively long.
- Mx1 inner lobe 1 with 4-5 spines plus a process ; inner lobe 2 with or without a seta ; endopod 1-segmented, bulbous, bearing 2-3 setae ; outer lobe 1 with 8 setae.
- Mx2 lobe 1 with 1-2 setae and a vestigial element ; lobe 2 with 2 setae ; lobe 5 bearing 1 stout spine without spinules along the outer margin.
- Mxp endopod setal formula : 1, 4, 4, 3, 3, 4 ; innermost seta on endopod segments 4 and 5 not rudimentary.
- Endopod segment 3 of P2 and P3 with 7 or 8 setae ; of P4 with 6 or 7 setae.
- P5 female with basipod 1 and the common plate fused or separate ; endopod completely or incompletely fused with basipod 2 , with 0-2 terminal setae ; basipod 2 and exopod fused or separate, exopod segment 1 fused or completely or incompletely separated from segment 2, segment 2 and segment 3 completely or incompletely fused , exopod with 2 terminal and 2-3 lateral spines ;
- P5 male with the right exopod 3-segmented, the distal 2 segments completely or incompletely separate, segment 2 expanded ; left basipod 2 with or without a 1-segmented, rudimentary endopod ; left exopod 3-segmented, segment 3 with 1 long spinulose process terminally.
After Ohtsuka & al. (1994, p.135) the genus is mainly distributed in the near-bottom communities on the continental shelf, but also colonizes marine caves.
Jaume, Cartes & Boxshall (2000) describe three new species of Paramisophria from the Mediterranean and consider that invasion from the shallow water hyperbenthic zone is the most likely colonisation route of anchialine caves by species of this genus. | | Remarks on dimensions and sex ratio: | | The mean female size is 1.708 mm (n = 20; SD = 0.7322), and 1.381 mm for the males (n = 24; SD = 0.3223). The size ratio (Male: Female) is 0.809. The sex ratio (F: M) = 0.82. | | | | | Parapseudocyclops Campaner, 1977 | |
| | Ref.: | Campaner, 1977 (p.812); Razouls, 1982 (p.531); Campaner, 1984 (p.107, Rem.); Ohtsuka & al., 1991 (p.793, 800, Rem.) | | Rem.: | Cf. Paramisophria | | | | (7) Paraugaptiloides Ohtsuka, Boxshall & Roe, 1994 | |
| | Ref.: | Ohtsuka & al., 1994 (p.120); Mauchline, 1998 (p.74: M); Bradford-Grieve,1999 b (p.37, Def.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.65); Ohtsuka & al., 2005 (p.2497: emend part.) | | Rem.: | Type: Paraugaptiloides magnus (Bradford,1974). Total: 2 spp.
Definition after Bradford-Grieve (1999, p.37), male only , female unknown:
As in the family definition with additional characters :
- Body relatively large ;
- Cephalosome and pediger 1 separate, pediges 4 and 5 fused.
- Prosome produced posteriorly with a small dorsolateral prominence and bluntly produced lateral lobe on each side.
- Lateral flap of cephalosome developed to cover bases of mouthparts.
- Caudal rami symmetrical with setae normally developed.
- Male left A1 19-segmented, fringed with setules along the anterior margin of segment 1 only.
- A2 endopod endopod segment 1 without an inner seta, segment 2 with an inner seta at midlength and 5+1 setae terminally ; exopod indistinctly 8-segmented, setal formula : 0, 1, 1, 1, 1, 1, 0, 3.
- Mandibular palp with the endopod rudimentary, 1-segmented, with 2 setae ; exopod segment 1 seta not reduced, exopod segment 5 with a relatively long outer seta.
- Mx1 inner lobe 1 with 5 spines and 1 process ; inner lobe 2 with 1 long seta ; basispod 1 withot seta ; endopod 1-segmented with 2 setae ; outer lobe 1 with 8 setae.
- Mx2 lobe 1 with 2 setae ; lobe 5 with 1 with 1 spine with 2 rows of spinules.
- Mxp endopod setal formula : 1, 4, 4, 3, 3, 4 ; the innermost seta on segments 4 and 5 not vestigial, seta on segment 6 reduced.
- P1 with 2 outer spines on exopod segment 3.
- P4 with a vestigial element on inner distal corner of basipod 1.
- P5 basipod 1 fused with common plate ; basipods 1 and 2 separate on left leg and incompletely fused on right. Right endopod 1-segmented, rudimentary, unarmed ; exopod segment 2 expanded inwards, almost completely separated from segment 3 which is triangular, tapering distally, with a minute outer and 1 terminal setule. Left endopod 2-segmented, unarmed ; exopod 3-segmented, distal 2 segments completely separate, segment 2 expanded inwards, segment 3 with 2 stout processes directed laterally.
For Ohtsuka & al. (1994, p.126) male P5 exhibits a more primitive state than Paraugaptilus in: (1) 2-segmented left endopod; (2) both exopods 3-segmented. The right 3rd exopodal segment of Paraugaptiloides is certainly movable with well developed muscles originating in the preceding segment, while the counterpart of Paraugaptilus is almost fused with the preceding segment and has reduced musculature, it is probably not movable. In addition, the 2nd segment of the left endopod in Paraugaptiloides is likely to be movable as indicated by the presence of a muscle extending between 1st and 2nd segments.
Emended part only after Ohtsuka & al; (2005, p.2497):
male left geniculate A1 fringed with long setules posteriorly on compound segments I-IV only or segments I-IV to V; segment I with 1 or 2 setae. 2nd endopodal segment of A2 with 2 or 3 inner setae. Md endopod 1-segmented with 2 terminal setae of unequal length, or absent.P4 with or without vestigial element on inner distal corner of coxa. Male P5 with basis and coxa of right leg separate or incompletely fused; right endopod unisegmented, rudimentary or well-developed; 2nd exopodal segment of right leg expanded inwards or extremely elongated, 3rd segment triangular, tapering distally, or lamellar; left endopod 2-segmented, with 2nd segment originating from inner base of 1st segment; 2nd exopodal segment of left leg expanded inwards or outwards, 3rd segment with 2 or 3 heavily chitinized, long processes. | | Remarks on dimensions and sex ratio: | | The mean male size is 4.930 mm (n = 3; SD = 0.2981.
The females are unknown at this date (2014/2015). | | | | (8) Paraugaptilus Wolfenden, 1904 | |
| | Ref.: | Wolfenden, 1904 (p.123); Van Breemen, 1908 a (p.138); A. Scott, 1909 (p.144); Wolfenden, 1911 (p.353); Sars, 1925 (p.335); Sewell, 1932 (p.329); Rose, 1933 a (p.241); Tanaka, 1964 c (p.234); Brodsky, 1950 (1967) (p.397); Deevey, 1973 (p.247, Rev, spp. Key); Razouls, 1982 (p.529); Campaner, 1984 (p.107); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.143, 158, 160, Redef., diagnosis emended); Chihara & Murano, 1997 (p.721); Mauchline, 1998 (p.71: F; p.74: M); Bradford-Grieve,1999 b (p.38, Def.); Boxshall & Halsey, 2004 (p.65); Vives & Shmeleva, 2007 (p.166); Renz & Markhaseva, 2015 (p.96, Table 4, Fig.3, biogeography) | | Rem.: | type: Paraugaptilus buchani. Total: 6 spp.
For Deevey (1973,p.257) the different species of Paraugaptilus have closely similar appendages. P5 females are apparently indistinguishable; only P. similis has somewhat different, slightly asymmetrical. The male P5 differ primarily in the shape and relative lengts of the two pincerlike spines on the last segment of the left leg. The mouthparts are very similar. The species are also all of about the same size ± 3 mm. The records indicate taht all apparently live within the upper 1,000 m. The scarcity of records must be due in part to the fact that the intermediate depths have been insufficiently sampled, but in addition the different species may have relatively restricted ranges. members of the genus are also rare; however, despite the paucity of numbers, the genus has a wide geographical distribution, having been recorded from 23°S in the Indian Ocean, from 1°30'N to 60°N in the Atlantic, and from about 54°N in the Pacific to 10°N, and in Indonesian waters.
After Ohtsuka & al. (1994, p.145 the genus is mainly distributed within the upper 1000 m, in particular, between 500 and 1000 m depths and appears to be meso- and bathypelagic.
Definition after Bradford-Grieve (1999, p.38)
:
As in the family definition with additional characters :
- Bofy relatively large.
- Cephalosome and pediger 1 separated or weakly fused, pedigers 4 and 5 fused.
- Last pediger segment with a short prominence or curved process dorsally and weakly developed lateral lobes.
- Genital segment with a pair of anteroventral gonopores, copulatory pores asymmetrically located posteroventrally, each copulatory duct heavily chitinised ; seminal receptacle relatively small.
- - Caudal rami symmetrical, longer than wide, with setae normally developed.
- A1 female symmetrical or slightly asymmetrical in ornamentation of terminal segments.
- Male left A1 19-segmented.
- A2 endopod segment 1 without an inner seta, segment 2 relatively shorter in male than in female with 1-2 setae medially in male, in female bearing 1 medial seta and 5+1 vestigial setae terminally ; exopod indistinctly 6-segmented, segment 6 rudimentary, unarmed in female, indistinctly 6-7-segmented in male.
- Mandibular blade with a tuft of setules ; 3 teeth on cutting edge, dorsalmost of which is bifid at tip ; palp endopod absent ; exopod segment 1 with a vestigial seta in female, (but with a well-developed seta in the male), segment 5 with a vestigial outer seta.
- Mx1 inner lobe 1 with 5 spines ; inner lobe 2 without seta ; endopod absent ; outer lobe 1 with 8 setae.
- Mx2 lobe 1 with 1 seta and 1 rudimentary element, lobe 2 with 1 seta, lobe 5 spine bipinnate ; endopodal setae with triangular spinules along inner margin.
- Mxp setal formula of the endopod : 1, 4, 4, 3, 3, 4.
- P1 exopod segment 3 with 2 outer spines.
- P4 with a minute inner seta on basipod 1 ; inner seta on basipod 2 ;
- Female P5 rudimentary, represented by a plate with a proximal seta and terminal or subterminal seta ;
- Male P5 with bothh basipod 1 fused with common plate ; basipods 1 and 2 separated on left leg and incompletely fused on right. Right endopod 1-segmented, rudimentary, unarmed ; exopod segment 2 expanded inwards, almost completely fused with segment 3 to form a compound segment, tapering distally, carrying a proximal seta and subterminal setule along the outer margin. Left endopod 1-segmented, unarmed ; exopod 3-segmented, last 2 segments almost fused, segment 2swollen medially, segment 3 with 2 stout, long, outwardly-directed processes terminally.
| | Remarks on dimensions and sex ratio: | | The mean female size is 3.220 mm (n = 10; SD = 0.3060), and for the males 3.120 mm (n = 10; SD = 0.0595). The size ratio (Male: Female) is 0.969. The sex ratio (F: M) = 0.833. | | | | (9) Pilarella Alvarez, 1985 | |
| | Ref.: | Alvarez, 1985 (p.189); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.156, Redef., diagnosis emended); Mauchline, 1998 (p.73: F); Bradford-Grieve,1999 b (p.40, déf.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.64) | | Rem.: |
Type: Pilarella longicornis Alvarez, 1985. Total: 1 sp.
Diagnostic characters after M.P.J. Alvarez (1985, p.189) :
Female :
- Cephalon and 1st pedigerous segment not fused, 4th and 5th pedigerous segments fused.
- Rostrum relatively wide, long and with 2 long tapering filaments (very thin distally).
Urosome 4-segmented.
- Anal segment short.
- Caudal rami very long (length about half that of the urosome).
- A1 asymmetrical, both with 21 segments, the left longer, and reaching the caudal rami.
- Mx1 with 5 spiniform setae on its 1st internal lobe; 1 setose seta on the 2nd internal lobe; 2 setae on the endopod; 5 setae on the 1st external lobe and 3 terminal setae on the exopod.
- The other buccal appendages and the P1 to P4 similar to those of the genus Metacalanus.
- P5 symmetrical and 3-segmented; 2nd segment globose with 1 external, short, distal seta and 1, internal, long seta; 3rd segment with 1 short lateral, external seta, 1 short seta and 1 long seta; endopod absent.
Male unknown.
Diagnostic characters after S. Ohtsuka, G.A. Boxshall & H.S..J. Roe (p.156) :
Female:
- Cephalosome and 1st pedigerous somite separate; ventrolateral corner of last prosome somite pointed.
- Urosome nearly half as long as prosome.
- Genital double-somite slightly wider than long; entire reproductive system paired, symmetrical; large circular gonopore and small copulatory pore located at outer and inner ends of slit-like aperture, respectively; copulatory duct short, simple; seminal receptacle relatively small, located medial to gonopore.
- Caudal rami slightly asymmetrical, with right ramus narrower and just shorter than left, with setae II and III relatively long.
- A1 asymmetrical, left longer than right and reaching to end of caudal rami; 21-segmented on both sides (segments I to IV fused, segments IX to XII partially fused, segments XXIV to XXVIII fused into compound apical segment).
- A2: 1st endopod segment with 1 mid-margin inner seta, 2nd with 3 setae at midlength and 5 setae terminally; exopod indistinctly 7-segmented.
- Md: Mandibular gnathobase lacking tuft of setules, 4 teeth on cutting edge, dorsalmost of which tricuspid; endopod rudimentary, 1-segmented with 2 setae; seta on 1st exopod segment not reduced; outer seta on 5th segment relatively long.
- Mx1: praecoxal arthrite with 6 elements (5 setae and 1 process); coxal epipodite with 5 setae; coxal endite with 1 long seta; basal seta absent; endopod bearing 2 setae.
- Mx2: 1st praecoxal endite with 2 setae; basal spine with 2 rows of spinules.
- Mxp: setal formula of endopod segments 1, 4, 4, 4, 3, 3, 4; setae a and b on 6th endopod segment relatively well developed.
- P1 with 1 outer spine on 3rd exopod segment.
- P4 with inner seta on coxa.
- P5: coxae separate from reduced intercoxal sclerite; endopod 1-segmented bearing 1 short spine on outer margin qnd 1 short and 1 long spine terminally.
-
For the authors the genital double-somite resembles that of Scutogerulus.
-
Type: Pilarella longicornis Alvarez, 1985. 1sp.
For Ohtsuka & al; (1994, p.158) as Alvarez (1985) has pointed out, the genus Pilarella is very similar to Metacalanus, but can be distinguished from the latter in the structures of A1, Mx1 and caudal rami. The genital double-somite of Pilarella resembles that of Scutogerulus. | | | | | (10) Protoparamisophria Ohtsuka, Nishida & Machida, 2005 | |
| | Ref.: | Ohtsuka & al., 2005 (p.2491) | | Rem.: | Type: Protoparamisophria biforaminis. Total: 1 sp.
For Ohtsuka & al. (2005, p.2492) the genus is similar to Paramisophria, but exhibits more plesiomorphic states both in the female genital system and in some appendages. Paired copulatory pores open at mid-length on the ventral side of the genital double-somite, while in Paramisophria a single copulatory pore is located midway or on either the left or right side. The new genus displays paired elongate seminal receptaclrs, each located between the corresponding copulatory pore and gonopore, with a short copulatory duct. These characteristics of the new genus seem to be relatively plesiomorphic with respect to the hypothetical calanoid ancestor as proposed by Huys & Boxshall (1991).
The exopod of P5 female displays the most primitive state in the family Arietellidae: indistinct ly 3-segmented; terminal segment with 1 outer, 1 subterminal and 2 terminal elements, all of which are separate at base from the segment. In some species of paramisophria the exopod is indistinctly 3-segmented, but the number of elements on the terminal segment is at most 3 elements and 1 process (in P. japonica). In this case, the outer terminal spine of the new genus also has the following plesiomorphies, although some species of paramisophria share some of them: (1) 5 spines and 1 process on the praecoxal arthrite of Mx1; (2) presence of a distinct seta on the coxal endite of Mx1; (3) 3 setae on the Mx1 endopod; (4) 2 setae and 1 vestigial element on the 1st praecoxal endite of Mx2; (5) separation of coxae and intercoxal sclerite in female P5; (6) 2 setae on endopod of female P5. | | | | (0) Rhapidophorus Edwards, 1891 | |
| | Ref.: | C.L. Edwards, 1891 (p.84); Giesbrecht, 1892 (p.423); Wilson, 1932 a (p.543, non M : 'F'); Fosshagen, 1968 a (p.61, 62, Rem.); Campaner, 1977 (p.817, Rem.); Razouls, 1982 (p.531); Campaner, 1984 (p.107, Rem.); Ohtsuka & al., 1991 (p.801, Rem.); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.131, Rem.); Mauchline, 1998 (p.68: M); | | Rem.: | For Fosshagen (1968 a, p.62) the description of the female should enable to know if the genus Paramisophria is not a synonym of this genus. Boxshall & Halsey, 2004 (p.64: Genus inquirendum). Total: 1 sp. | | | | (11) Sarsarietellus Campaner, 1984 | |
| | Syn.: | Scottula (part.) Sars, 1905 c (p.22); 1925 (p.338, figs.F,M) | | Ref.: | Campaner, 1984 (p.107, Rem.); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.155, 158, 160, Redef., diagnosis emended); Mauchline, 1998 (p.68: F; p.76: M); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.65); Ohtsuka & al., 2005 (p.2504, emend. part.) | | Rem.: | type: Scottula abyssalis Sars,1905. Total: 4 spp.
Emended part, issued from Ohtsuka & al. (2005, p.2504):
Female (only known): Cephalosome with paired papillae anterolaterally. Prosomal end with or without weakly or strongly developed dorsolateral and ventrolateral processes. Genital double-somite slightly or hightly asymmetrical; single copulatory pore located medially or on right side. A1 22- or 23-segmented. Exopod of P5 unisegmented, with fusion lines visible or not.
The anterolateral cephalic papillae bear a pore on tip, this organ may have a secretory function. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.705 mm (n = 4, SD = 1.069). Only 1 male is known with a body length of 3.02 mm. The ratio male: female is 0.873 (from the same species) | | | | Scottula Sars, 1902 | | Ref.: | Sars, 1902 (1903) (p.124); Van Breemen, 1908 a (p.142); A. Scott, 1909 (p.146); Rose,1933 a (p.243); Tanaka, 1964 c (p.242); Brodsky, 1950 (1967) (p.394); Razouls, 1982 (p.532); Campaner, 1984 (p.102, 107, Rem.); Ohtsuka, 1985 (p.305); Mauchline, 1988 (p.706) | | Rem.: | Cf. Metacalanus (part.) & Sarsarietellus (part.) | | | | (12) Scutogerulus Bradford, 1969 | |
| | Ref.: | Bradford, 1969 b (p.495); Razouls, 1982 (p.531); Campaner, 1984 (p.107); Razouls, 1993 (p.306); Ohtsuka & al., 1994 (p.152, 160, Redef., diagnosis emended); Mauchline, 1998 (p.68: F); Bradford-Grieve,1999 b (p.39, Déf.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.64), Ohtsuka & Boxshall, 2004 (p.54: Rem.) | | Rem.: | Type: Scutogerulus pelophilus Bradford, 1969. Total: 2 spp.
Definition after Bradford-Grieve (1999, p.38) female only, male unknown :
As in the family definition with additional characters :
- Body relatively large.
- Cephalosome and pediger 1 separate, pedigers 4 and 5 fused.
- Urosome about 193 the length of prosome.
- Genital segment as long as wide ; gonopore and copulatory pore sharung a common slit-like aperture, gonopore located anteriorly, copulatory pore at innermost corner of slit ; copulatory ductswollen anteriorly ; seminal receptacle relatively small and simple in shape.
- Caudal rami slightly asymmetrical , left ramus longer than the right, longer than wide.
- A1 symmetrical, extending almost at the end of prosome ; posterior margins of proximal segments bear long setules from segment I-XIII.
- A2 Endopd segment 1without an inner seta ; exopod indistinctly 8-segmented.
- Mandibular palp with a rudimentary endopod, 1-segmented with 2 setae ; seta on exopod segment 1 not reduced, segment 5 with a relatively long seta.
- Mx1 inner lobe 1 with 4 spines and a process ; inner lobe 2 with 1 long seta; basipod 2 with 1 short seta ; endopod with 1 seta ; outer lobe 1 with 6 setae.
- Mx2 lobe 1 with 1 relatiçvely well-developed seta and 1 vestigial element ; lobe 2 with 1 seta ; lobe 5 spine with 3 rows of minute spinules ; setae on endopod with a row of triangular spines.
- Mxp with the setal formula on the endopod : 1, 4, 4, 3, 3, 4.
- P1 exopod segment 1 with an outer medial tuft of short setules and a subterminal outer spine.
- Female P5 biramous, with a 1-segmented rudimentary endopod with 1 terminal seta and a 2-segmented exopod with 1 outer spine on segment 1 and 2 terminal setae on segment 2.
Ohtsuka & Boxshall (2004, p.54) point out that the modified 'shield-shaped' setules on Mx2 and Mxp are not found in other calanoids. They are superficially similar to the 'disc' shape on the setae of Euaugaptilus. However, these stuctures differ remarkably in : 1- the 'shield' is crescent-shaped in Scutogerulus, while the 'disc' is semicircular or circular in Euaugaptilus; 2- the 'shields' are arranged in a single row in Scutogerulus, whereas 2 rows of 'discs' arranged either side-by-side or alternately are present in Euaugaptilus; 3- each 'shield' is furnished with two supplementary plates beneath it, which are absent in Euaugaptilus; 4- the 'stalk' connecting the 'schield' to a seta is lamellar in Scutogerulus, but columnar in Euaugaptilus; 5- the 'shield' is positioned obliquely to the seta, while the 'disc' is parallel to it; 6- the outer surface of 'shield' is wrinkled (perhaps an artefact) in Scutogerulus but smooth in Euaugaptilus.
The fine structure of the 'button setae' on the maxillae and maxillipeds of the carnivorous arietelloidean family Augagaptilidae, in particular the genus Euaugaptilus, has been examined by Boxshall (1985) and Matsuura & Nishida (2000). They concluded from a functional morphological point of view that these unique structures may function as shock absorbers during prey capture.
For Ohtsuka & Boxshall the genus Scutogerulus appears to be carnivorous based on gut contents containing copepod fragments. as in other arietellid genera. Shallow-water arietellid genera such as Paramisophria and Metacalanus bear relatively simple spinulose setae on Mx2 and Mxp, while deep-sea forms such as Arietellus and Scutogerulus carry specialized setae on these feeding appendages. This seems to be an adaptative strategy of calanoids in the oligotrophic deep-sea environment. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.225 mm (n = 4; SD = 0.3880. The males are unknown at this date (2014). | | | | | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 23, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|



;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
