|
|
 |
|
Calanoida ( Order ) |
|
|
|
Spinocalanoidea ( Superfamily ) |
|
|
| |
| | | |
| Spinocalanidae Vervoort, 1951 ( Spinocalanoidea ) | | Syn.: | no Arctokonstantinidae Markhaseva & Kosobokova, 2001 (p.319) | | Ref.: | Vervoort, 1951 (p.10, 72, Def.); Farran & Vervoort, 1951 g (n°39, p.3); Mazza, 1967 (p.99); Grice, 1971 (p.273, 279, key); Andronov, 1974 a (p.1005); Damkaer, 1975 (p.13, Genera Key); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.137); Fleminger, 1983 (p.605, 607, Genera Key, Rem.); Brodsky & al., 1983 (p.144, 147, 242, Genera Key); Park, 1986 (p.192, 193, 194); Mauchline, 1988 (p.724, 740: cuticular pores); Huys & Boxshall, 1991 (p.467); Razouls, 1991 (p.209, Rem.); Schulz, 1992 (p.176, Genera Key); Razouls, 1993 (p.311); Bradford-Grieve, 1994 (p.94); Schulz, 1996 (p.599, Rem., clé des G.); Bradford-Grieve & al., 1999 (p.878, 903, 904, 913: Genera key); Markhaseva & Kosobokova, 2001 (p.319, 323, Rem.); Ohtsuka & Huys, 2001 (p.461); Boxshall & Halsey, 2004 (p.12, 15; 49; 195: Def.; p.198: Genera Key); Schulz, 2004 (p.211: Rem.); Vives & Shmeleva, 2007 (p.820, part. Genera key); Markhaseva, 2008 (p.15, Rem.); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, molecular biology, phylogeny); Bode & al., 2017 (p.600, Fig.2, 3, 4, 5, Table I, II, III: morphology vs phylogeny); Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships); Hirai & al., 2020 (p.1, Fig.4: metabarcoding, Fig.8: OTUs distribution patterns)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | According to Vervoort (1951, p.72), Farran, in his manuscripts of the 'Zooplankton Sheets', formed a new familly, Spinocalanidae, to comprise such genera as Spinocalanus Giesbrecht, 1888, Monacilla Sars, 1905 and Mimocalanus Farran, 1908. Although Farran gives no diagnosis of this new family it seems clear that he based it on the quite different arrangement of the setae on the swimming legs and the family has been defined on such lines in the revision of the 'Sheets'. 10 G.: Damkaeria, Isaacsicalanus, Kunihulsea, Methanocalanus, Mimocalanus, Monacilla, Mospicalanus, Rhinomaxillaris, Spinocalanus, Teneriforma. When estimating whether a particular characteristic is primitive or advanced, Markhaseva (2008, p.12 and see table) follows the criteria generally accepted among copepodologists : the evolution of copepods is characterized by oligomerization (V.A. Dogel, 1954) and follows, primarily, the pathway of reductions, fusions, and losses (Boxshall, Ferrari & Tiemann, 1984), i.e, the condition of the greatest articulation and setation of appendages is considerd to be the most primitive one.
For Markhaseva (2008, p.12-13 and p.15: table) the genera Arctokonstantinus, Sognocalanus, Foxtonia and Foxtosognus belong to the new family Arctokonstantinidae.
After Markhaseva & Schulz (2008, p.287), concerning the ancestral mode of life of Spinocalanidae, some speculation are possible. Due to a more primitive morphology of pelagic representatives of spinocalanids, it may be assumed that they probably originated in the pelagic environment.
Typically, pelagic spinocalanid genera (Monacilla, Spinocalanus, Mimocalanus, Mospicalanus, Teneriforma) demostrate diversity of some primitive characters not observed in their benthopelagic relatives (Damkaeria, Kunihulsea, Methanocalanus, Isaacsicalanus). The following plesiomorphies are considered typical of pelagic spinocalanids :
1- 11 setae on Md endopod segment 2 in Spinocalanus, vs. 9-10 setae in other spinocalanids.
2- 15 setae on Mx1 praecoxal arthrite in Monacilla, vs. fewer in other spinocalanids.
3- 6 setae on Mx1 coxal endite in Spinocalanus, vs. fewer in other spinocalanids.
4- 6 setae on Mx1 distal basal endite in Teneriforma, vs. fewer in other spinocalanids.
5- 16 setae on endopod and 11 setae on exopod of Mx1 in Monacilla, Spinocalanus, Mimocalanus (but 23 on endopod in M. crassus) ; exopod setation of this type also in Mospicalanus, vs. fewer in other spinocalanids.
6- Mx2 with 9 setae (2+7) on endopod fused to distal basal endite in Monacilla, Spinocalanus, vs. fewer in other spinocalanids.
7- praecoxal endites of Mxp syncoxa with 1, 2, 3 setae in Monacilla, Spinocalanis, vs. fewer in other spinocalanids.
8- coxal endite of Mxp syncoxa with 4 setae in Monacilla, Mimocalanus, vs. fewer in other spinocalanids.
Typically, pelagic Spinocalanidae share some primitive characters with more derived benthopelagic spinocalanid genera :
1- the primitive type of A2 exopod setation 1, 1-1-1, 1, 1, 1, 1, 1 and 3 setae, is shared by pelagic Monacilla, Spinocalanus, Teneriforma and the benthopelagic genus Kunihulsea.
2- the most primitive condition in the armature of the proximal basal endite of Mx1 (4 setae) is found in the pelagic genera Monacilla, Spinocalanus , Mimocalanus, Mospicalanus as well as the benthopelagic Methanocalanus.
3- the most primitive armature of the proximal praecoxal endite of Mx2 (7 setae) is shared by the pelagic Monacilla and the benthopelagic Methanocalanus.
Key to adult females of Spinocalanidae Genera after Schulz (1992, p.176) (Mospicalanus, Rhinomaxillaris, and Methanocalanus not included) :
1 Rostrum absent
.2.
1 - Rostrum present
.. 3.
2 P2 to P4 with spines on posterior surfaces
. Spinocalanus.
2 P2 to P4 lacking spines on posterior surfaces
.. Mimocalanus.
3 Rostrum bifurcate produced into 2 strong spiniform processes
.. Monacilla.
3 Rostrum uniramous, cone-shaped or knob-like
. 4.
4 Mxp basis with 3 medial setae
. 5.
4 » - Mxp basis with 2 medial setae
. 6.
5 A1 reaching to second urosomal segment. P1 exopodal segment 1 with outer spine
. Damkaeria.
5 A1 reaching at least to caudal rami. P1 exopodal segment 1 lacking outer spine
.. Kunihulsea.
6 Total length more than 2.0 mm. Rostral cone short. A1 about as long as prosome. P1 exopodal segment 2 lacking outer spine
.. Isaacsicalanus.
6 Total length more than 2.0 mm. Rostral cone long. A1 reaching third urosomal segment. P1 exopodal segment 2 with outer spine
.. Teneriforma.
Key to genera from Schulz (1996, p.599) [Sognocalanus excluded, belons now to Arctokonstantinidae family] :
1 - Rostrum absent ...... 2.
1' - Rostrum present, bifurcated or simple ....... 4.
2 - P2-P4 exopod and/or endopod with spinules on posterior surface. Male anal somite usually reduced ......Spinocalanus.
2' - P2-P4 exopod, endopod lacking spinules on posterior surface ...... 3.
3 - P3-P4 endopodal segment 3 with 0-1 outer seta. A1 usually narrowing between ancestral segments X-XI and XIII. A2 exopod longer than endopod. Male P5 uniramous ...... Mimocalanus.
3' - P3-P4 endopodal segment 3 with 2 outer setae. A1 of uniform width medially. A2 exopod length subequal to endopod. Male unknown ....... Mospicalanus.
4 - Rostrum with 2 terminal points ..... 5.
4' - Rostrum single, cone-shaped or lobate ...... 6.
5 - Rostrum bifurcate, with 2 strong points in female , reduced to knob in male. P1 exopodal segment 1 with outer distal spine in female, reduced in male. Male anal somite about half length of caudal rami ......... Monacilla.
6 - Rostrum cone-shaped. P1 exopodal segments 2 and 3 each with outer distal spine ...... 7.
6' - Rostrum a simple, lobate process. P1 exopodal segments 1 and 2 without outer distal spine ........ 9.
7 - P1 exopodal segments 1-3 each with long outer distal spine. A2 exopod with 11 setae. Male P5 uniramous, extremely long, present on right side only ....... Damkaeria.
7' - P1 exopodal segment 1 without outer distal spine. A2 exopod with 12 setae ........ 8.
8 - Rostrum a medium-sized cone. A1 extending to caudal rami, with 23 free segments (fusion of ancestral segments XXVI-XXVIII). Mxp basis with 3 medial setae ....... Kunihulsea.
8' - Rostrum a long cone. A1 extending to mid-urosome, with 24 free segments (ancestral segments XXVI and XXVII-XXVIII separated). Mxp basis with 2 medial setae. Male P5 uniramous, present on left side only ........ Teneriforma.
9 - Prosome produced into sharp point posteriorly. Body length > 2 mm. Mxp basis with 2 medial setae. Male unknown ......... Isaacdsicalanus.
9' - Prosome not pointed posteriorly. Body length < 1.5 mm. Mxp basis with 3 unequal medial setae. Male unknown.
Key genera after Boxshall & Halsey (2004, p.198]) (modified after Schulz, 1996), [Methanocalanus not included; Arctokonstantinus, Sognocalanus and Foxtonia excluded] :
1 P2 with 2-segmented endopod ; P3 with 3-segmented endopod ; Rostrum absent
.. 2.
1 - P2 with 2-segmented endopod ; P3 with 3-segmented endopod ; Rostrum present, bifurcate or simple
.. 4.
2 Posterior surfaces of exopods and or endopods of P2 to P4 ornamented with spinules ; male anal somite usually reduced
.. Spinocalanus.
2 Posterior surface s of exopods and or endopods of P2 to P4 lacking spinular ornamentation
.. 3.
3 3rd endopodal segment of P3 and P4 with 1 outer margin seta, or lacking seta on outer margin ; A1 usually narrowing between segments X-XI and XIII ; exopod of A2 longer than endopod ; male P5 uniramous
..Mimocalanus.
3 3rd endopodal segment of P3 and P4 with 2 outer margin setae ; A1 segments X-XI to XIII of uniform width ; rami of A2 about equal in length ; male unknown
Mospicalanus.
4 Rostrum bifurcate, with 2 strong points in female, reduced to knob in male ; 1st exopodal segment of P1 with outer distal spine in female, reduced in male ; male anal somite about half length of caudal rami
..Monacilla.
4 Rostrum simple conical or lobate
5.
5 - Rostrum conical ; 2 nd exopodal segment of P1 with outer distal spine
. 6.
5 Rostrum a simple lobate process ; 2 nd exopodal segment of P1 without outer distal spine
..8.
6 1st exopodal segment of P1 with long outer distal spine ; exopod of A2 with 11 setae ; male P5 uniramous, elongate, present on right side only
Damkaeria.
6 1st exopodal segment of P1 without outer distal spine ; exopod of A2 with 12 setae
. .7.
7 Rostrum a medium-sized cone ; A1 extending to caudal rami with ancestral segment XXVI incorporated into triple apical segment XXVI-XXVIII ; Mxp with 3 setae on basis
.Kunihulsea.7 Rostrum a long cone ; A1 extending to mid-urosome level, with ancestral segment XXVI separate from apical segment XXVII-XXVIII ; Mxp with 2 setae on basis
.Teneriforma.
8 Prosome produced into acute point posteriorly ; basis of Mxp armed with 2 setae medially
. Isaacsicalanus.
8 Prosome not pointed posteriorly ; basis of Mxp armed with 3 setae
.Rhinomaxillaris.
After Bode & al. (2017, p.612), the phylogenetic analysis of both genes yields shows that Mimocalanus and Teneriforma as sister lineages to all pelagic Spinocalanidae. This differs from the phylogenetic hypotheses of the Spinocalanidae based on morphological characters (Fleminger, 1983; Schulz, 1989), which place Monacilla as a sister-group to the genera Spinocalanus, Mimocalanus Teneriforma, Isaacsicalanus, Mospicalanus. The results of the present phylogenetic analyses differ also in the placement of the genus Mospicalanus. While the morphological analysis suggests a close relationship of Mimocalanus and Mospicalanus, the genetic data of the present study rather indicate a close relationship of Mospicalanus and Spinocalanus A (see in Figs. 2 and 3). Furthermore, there is evidence in both gene fragments that the genus Spinocalanus may be polyphyletic, which coincides with the morphological separation of the species groups: Spinocalanus A and Spinocalanus B by Damkaer (1975). It has to be taken into account that only 5 of the 10 known genera of the Spinocalanidae were analysed and , thus, the placement of the genera may change both with the addition of genes with different mutation rates (i.e nuclear 28S rDNA, internal transcribed spacer 1 and 2) and with further genera.
For Bode & al. (2017, p.612), Spinocalanus genus comprised more species than previously described based on morphological characters [see list in species database. CR]. There is evidence that some of these, such as the S. dispar complex, may be cryptic or pseudocryptic species, which is a common phenomenon in marine copepods (Bucklin & al., 1999; Goetze, 2003; Bucklin & Frost, 2009; Cornils & Held, 2014; Hirai & al., 2015). However, in net samples 2nd exo- and endopodites, as well as A1 are often missing [like other appendages]; this makes molecular species identification methods indispensible for numerous fragile copepods, particularly from deep-sea.
The present study provides first insights into the molecular diversity of the morphologically challenging end ecologically family Spinocalanidae by analysing mitochondrial and nuclear gene fragment , as well as protein mass fingerprints. The different methods (Figs. 2, 3, 4) agree well in the number of lineages found within this family
The study of Bode & al. (2017, p.612) demostrates that many of the spinocalanid species have much wider distributional ranges than previously thought [this is an generality for all the deep-sea, bathypelagic and abyssopelagic, copepods. CR.] and the molecular results provide evidence that family is much more diverse than previously described based on morphological characters alone.. For example S. horridus, S. longispinus and S. stellatus, mainly found in Artic or Antarctic waters, also in the NE and NW Pacific. S. horridus (= longispinus) and S. elongatus were even classified as Arctic endemic (Kosobokova & al., 2011) [if we admitted the synonymy. CR]. Hence, molecular comparisons of populations from tropical and polar regions may be useful into the true diversity and phylogeography of this family. Thus, the true global species richness of this copepod family seems to be underestimated.
After Bode & al. (2018, p.75) the detritivorous species in this family, apparently feed on a large spectrum of particle sizes, while some species have evolved paricularly efficient digestive systems with extremely elongated and looped mid-guts (Kosobokova & al., 2002; Wishner & al., 2008). |  issued from : E.L. Markhaseva in Russian J. Mar. Biol., 2008, 34 (1). [p.15]. Details of armament of mouthparts and P1 in Spinocalanidae. The table presents the most primitive condition of characters that has revealed for the genus. |
 issued from : E.L. Markhaseva & K. Schulz in Zootaxa, 2008, 1866. [p. 279]. Selected character states of Spinocalanoidea females (diagnostic characters of the family Arctokonstantinidae are in bold). Refernces: Hulsemann & Grice, 1963; Fosshagen, 1967, 1983; Park, 1970; Damkaer, 1975; Brodsky & al., 1983; Schulz, 1989, 1996, 2004; Markhaseva & Kosobokova, 2001; Boxshall & Halsey, 2004; Ivanenko & al., 2007; Markhaseva & Schulz, 2008. Nota: According to the authors (p.288), Arctokonstantinidae and Spinocalanidae belong to the Spinocalanoidea, the sister group of the Ryocalanoidea. Diagnostic characters considered to distinguis the superfamily Spinocalanoidea from Ryocalanoidea are not very strong. |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.195]. Armature formula of swimming legs P1 to P4. |
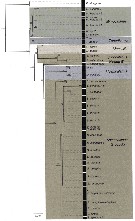 Issued from : M. Bode, S. Laakmann, P. Kaiser, W. Hagen, H. Auel & A. Cornils in J. Plankton Res., 2017, 39 (4). [p.607, Fig.2]. Reconstruction of evolutionary relationships among Spinocalanidae. RAxML Maximum likelihood tree for the 18S rDNA fragments (1674 bp alignment). Numbers at the nodes represent the percentage bootstrap values > 50%. For details on species names see the considered species. Nota: Five genera of Spinocalanidae were identified morphologically: Spinocalanus Giesbrecht, 1888; Mimocalanus Farran, 1908; Monacilla Sars, 1905; Teneriforma Grice & Hülsemann, 1967; Mospicalanus Schulz, 1996. The phylogenetic analysis of the nuclear 18S gene fragment on a subset of 92 specimens yielded a tree with high bootstrap values Except for the genus Spinocalanus, all the other four genera were monophyletic [bootstrap support (BS): 87-100%]. The genus Spinocalanus, however, was polyphyletic. One highly supported clade consisted of S. magnus Wolfenden, 1904 and S. antarcticus Wolfenden, 1906 (BS: 100%), forming a well-supported clade (BS: 90%) with the genus Monacilla. A second clade was formed by a singletion sequence of S. angusticeps Sars, 1920, and a third large clade comprised all other identified Spinocalanus species (BS: 99%), forming a moderately supported (BS: 78%) clade with the genus Mospicalanus. From hereon we will refer to the three species S. magnus, S. antarcticus, S. angusticeps as Spinocalanus B and to all other Spinocalanus species as Spinocalanus A, following the morphological classification of Damkaer (1975). In the phylogenetic tree of 18S, the genera Miimocalanus and Teneriforma formed a highly supported (BS: 100%) sister-group to all other pelagic spinocalanid genera (figs.2 and 3). This pattern is also visible in the maximum likelihood tree of the mitochondrial COI, however, the nodes are not well supportede. The genera Monacilla and part of Spinocalanus B ( S. magnus, S. antarcticus) were sisters to S. angusticeps, Mospicalanus and Spinocalanus A (BS: 91%). This pattern differed slightly in the phylogenetic tree of COI, placing Spinocalanus B as sister to all remaining taxa, and Monacilla again as sister to S. angusticeps, Mospicalanus and Spinocalanus A. In the phylogenetic analysis of COI, Mospicalanus and Spinocalanus A could not be separated. Genetic divergence between the 7 clades (measured as uncorrected p distance) ranged between 0.3 and 2.9%. The divergence within clades ranged between 0 and 0.5%, except for Spinocalanus A, where p distance varied between 0.0 and 1.4% due to high divergences of Spinocalanus brevicaudatus Brodsky, 1950 and S. elongatus 2 [see fig. 3; and in species database. CR]. |
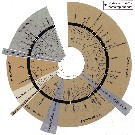 Issued from : M. Bode, S. Laakmann, P. Kaiser, W. Hagen, H. Auel & A. Cornils in J. Plankton Res., 2017, 39 (4). [p.608, Fig.3]. Condensed Maximum likelihood tree for the mtCOI gene fragments (667 bp alignment). Nodes with bootstrap support <50% were collapsed, nodes with bootstrap support >90% are marked with black stars (for all bootstraps see Supplentary Fig.2. Currently acknowledged genera and species illustrated. For details species names see Table I [[indicated in the species database. CR.]] |
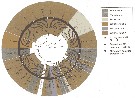 Issued from : M. Bode, S. Laakmann, P. Kaiser, W. Hagen, H. Auel & A. Cornils in J. Plankton Res., 2017, 39 (4). [p.609, Fig.4]. Cluster analysis of the MALDI-TOF mass spectra based on Euclidean distance. Black stars at the nodes represent the percentage bootstrap values >90% (fotr all bootstraps >50%, see Supplementary Fig.3). Currently acknowledged genera and species are illustrated as in Fig.2 and 3. Black dots indicate additional taxa in comparison with COI data. Black square indicates incongruence to COI data. For details species names see Table I [[indicated in the species database. CR.]]. Nota: the last metasomal segment/urosomes of 193 spinocalanoid adult females were applying the MALDI-TOF MS technology according to available protocols (Riccardi & al., 2012; Laakmann & al., 2013). Additionally, seven whole animals were analysed to test whether they clustered correctly with the urosome-only samples. | | | | | Arctokonstantinus Markhaseva & Kosobokova, 2001 | | Ref.: | Markhaseva & Kosobokova, 2001 (p.320); Boxshall & Halsey, 2004 (p.198); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | Cf.
Arctokonstantinidae | | | | (1) Damkaeria Fosshagen, 1983 | |
| | Ref.: | Fosshagen, 1983 (p.260, 261); Schulz, 1989 (p.195, Rem.); Bradford-Grieve, 1994 (p.95, Def.); Mauchline, 1998 (p.87, 89: F); Boxshall & Halsey, 2004 (p.198) ; Schulz, 2004 (p.198, Def.); Markhaseva, 2008 (p.15, Rem.); Renz & Markhaseva, 2015 (p.96, Table 4, fig.3, biogeography) | | Rem.: | Type: Damkaeria falcifera Fosshagen,1983.
This genus, included previously among the Bathypontiidae, was transferred in Spinocalanidae family by Bradford-Grieve (1994, p.95). Total: 2 spp.
Diagnosis from Bradford-Grieve (1994, p.95) :
- As for the family definition.
- All pedigers separate.
- pediger 5 rounded posteriorly.
- Rostrum pointed, single and directed backwards.
- Caudal rami slightly longer than the anal segment.
- A1 female extends a little beyond genital segment, 23-segmented, segments 8-9 and 24-25 fused.
- A1 male 22-segmented on the right, 23-segmented on the left ; segments 20-21 fused on the right.
- A2 exopod longer and stronger than the endopod.
- Md with 3 setae on the inner margin of basis ; blade with sharp teeth, the most ventral of which is set apart from the others.
- Mx1 female with some reduction in inner lobe 3, basis and endopod where the size and number of setae reduced ; inner lobes 2 and 3 with 3 and 4 setae respectively.
- Mx1 male shows further reduction, particularly in inner lobe 2 which is smaller and has 2 small setae ; inner lobe 1, exopod an douter lobe 1 are similar to that of the female.
- Mx2 bears a strong modified claw-like spine on lobe 4 ; distally this spine is strongly armed.
- Mxp female with coxo (basipod 1) and basis (basipod 2) about the same length ; there is a longitudinal row of spinules on basis ; endopod segments 1 and 2 of about equal length.
- Mxp male reduced in size and has a reduced number of setae compared with the female.
- Basis of P1 without minute outer distral seta ; exopodal segments 1 and 2 with an outer-edge spine ; endopod with 2 terminal and 3 inner edge setae ; male exopod outer edge spines shorter and weaker than in the female.
- There are weak surface spinules on the basipods of P2-P4 which are particularly dense on the posterior surface of basis of P4 ; the surfaces of the exopods and endopods of P2-P4 without ornamentation.
- Endopodal segment 2 of P2 with 1 outer-edge seta,
- Endopodal segment 3 of P3 and P4 with 2 outer-edge setae.
- Endopodal segment 1 of P3 and P4 apparently with outer distal corner rounded.
- Female P5 absent.
- Male P5 present on the right side only as a uniramous, 4-segmented, long appendage which is twice the length of the urosome when extended ; first segment broad with a proximal thickening, other segments slender, and the distal segment has a longitudinal row of stiff spinules which increase in length distally. | | Remarks on dimensions and sex ratio: | | The mean female size is 0.870 mm (n = 4; SD : 0.0589) and the mean male size 0.718 mm (n = 4; SD = 0.0350). The size ratio (male : female) is 0.825.
The sex ratio is 1. | | | | Foxtonia Hulsemann & Grice, 1963 | | Ref.: | Hulsemann & Grice, 1963 (p.733); Razouls, 1982 (p.593); Fosshagen, 1983 (p.260); Razouls, 1993 (p.308); Mauchline, 1998 (p.66); Boxshall & Halsey, 2004 (p.198) | | Rem.: | Cf.
Arctokonstantinidae | | | | (2) Isaacsicalanus Fleminger, 1983 | |
| | Ref.: | Fleminger, 1983 (p.606, 607, 615); Schulz, 1989 (p.195, Rem.); Razouls, 1993 (p.311); Mauchline, 1998 (p.87: F); Boxshall & Halsey, 2004 (p.198); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | Type: Isaacsicalanus paucisetus Fleminger,1983. Total: 1 sp.
Diagnosis from Fleminger (1983, p.606) :
- Female with elongate urosome about half length of prosome.
- Short A1 about as long as prosome.
- Rostrum short, lobate without any semblance of bifurcation.
- Mx1 with basis fused to proximal part of endopodite and bearing 4 inner setae; inne lobe 2 with 1 seta, inner lobe 3 lacking inner seta.
- Distal inner setae of Mx2 and Mxp unarmed except for very fine setules on proximal side.
- Mxp coxa with 2 inner setae.
- Exopodal segments 1 and 2 of P1 lacking outer setae.. Endopod with 5 inner setae and prominent shoulder (lateral swelling produced anterodistad, knob-like) at segment's midlength and bearing spinules concentrated at apex.
- P2-P4 with enxopodal segment 3 outer seta 3 about twice as long as outer seta 1 and seta 2. Posterior surfaces of P2-P4 without spines.
- Exopodal segment 1 of A2 without inner seta, exopodal segments 2 and 3 fused, exopodal segment 2 with 1 inner seta.
Diagnosis from Bradford-Grieve (1994, p.95) :
- As for the family definition.
- Pediger segments 4 and 5 partly fused.
- Caudal rami asymmetrical because of the differing position of the inner-edge setae ; the left inner seta located on the ventral surface and projects ventrally, the right inner seta located on the dorsal surface, inserted more distally than the left seta, and directed dorsally.
- Rostrum short, lobate, without bifurcation.
- Female A1 short, 23-segmented with segments 8-9, 24-25 fused.
- A2 exopod 1 without setae, exopodal segments 2 and 3 not fused, exopodal segment 2 with 1 seta.
- Mx1 with basis fused to the endopod and bearing 8 setae ; inner lobe 2 with 1 seta, inner lobe 3 without setae.
- Mx2 and Mxp with distal setae unarmed except for very fine setules on the proximal side ;
- Mxp basipod 1 with 2 setae, basipod 2 with a short row of stiff hairs proximal to the medial setae.
- P1 exopodal segments 1 and 2 without an outer-edge spine ; endopod with 5 setae and a prominent lateral swelling at the segments midlength which bears spinules concentrated at its apex.
- Outer distal spine on exopodal segment 3 of P2-P4 about twice as long as the two proximal spines.
- Posterior surfaces of P2-P4 without spines.
- Female P5 absent.
For Fleminger (1983) this genus occupies a highly derived position within the Spinocalanidae. Schulz (1989) points out the similarities between Isaacsicalanus and Teneriforma. | | | | (3) Kunihulsea Schulz, 1992 | |
| | Ref.: | Schulz, 1992 (p.176); Mauchline, 1998 (p.87: F); Boxshall & Halsey, 2004 (p.198) ; Schulz, 2004 (p.206, Rem. : emend.); Markhaseva, 2008 (p.15, Rem.); Renz & Markhaseva, 2015 (p.96, Table 4, Fig.3, biogeography) | | Rem.: | Type: Kunihulsea arabica Schulz,1992. Total: 2 spp.
Diagnosis from Schulz (1992, p.176) :
Very small body length .
- Female with short urosome about 1/3 length of prosome.
- Rostrum a single short cone.
- A1 reaching to caudal rami; segments 24 and 25 fused.
- Exopod of A2 about as long as endopod.
- Lobe 3 of Mx1 smaller than lobe 2.
- Mxp with 3 medial setae on basis; endopodal segments 1 to 4 armed with 2 setae each.
- P1 exopodal segment 1 without outer spine; endopod particularly long, with small outer lobe.
- P2 to P4 with numbers of segments and setae typical of the family; exo- and endopods without posterior spines.
- Male (juvenil CV) with left-handed P5.
For Schulz (1992, p.177) this genus shows affinities to spinocalanids: 1- 4 outer setae on the 3rd exopodal segment of P1; 2- 5 setae on the 3rd exopodal segment of P2; 3- separation of pedigers 1 and 2 as well as pedigers 4 and 5; 4- the shape of the lateral posterior corner of pediger 5; 5- the conical undivided rostrum, are characters reminiscent of most or at least some spinocalanid genera, respectively. Among Spinocalanoidae, the genus might occupy a phylogenetic position between the three basic genera, Monaciulla, Spinocalanus, Mimocalanus, and the two most derived taxa Teneriforma, Isaacsicalanus. The conical rostrum as a synapomorphy seems to link Kunihulsea with Damkaeria and the two latter genera, provided the origin of this structure is monophyletic within the Spinocalanoidea.
Other plesiomorphic characters found in both Kunihulsea and Damkaeria may prove a grouping prior to the splitting off from the stem leading to Teneriforma and Isaacsicalanus: (a) 14 (Damkaeria) and 13 setae (Kunihulsea), but 11 (Teneriforma), respectively, on the 1st inner lobe (arthrite) of Mx1, and (b) 8 and 7 (5) setae on the syncoxa of Mxp, and 3 medial setae (2 in Teneriforma) on the basis of Mxp.
However, any definite placement of Kunihulsea and Damkaeria must await future assessement when exact setal armament of the mouth appendages has been verified. | | Remarks on dimensions and sex ratio: | | The mean female size is 0.597 mm. (n = 4, SD = 0.229). Adult males unknown. | | | | (4) Methanocalanus Ivanenko, Defaye & Cuoc, 2007 | |
| | Ref.: | Ivanenko & al., 2007 (p.38); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | 1 sp. | | | | (5) Mimocalanus Farran, 1908 | |
| | Ref.: | Farran, 1908 b (p.22); Sars, 1925 (p.34); Rose, 1933 a (p.86); Vervoort, 1946 (p.156); Davis, 1949 (p.23); Farran & Vervoort, 1951 h (n°40, p.3); Damkaer, 1975 (p.67, spp. Key); Brodsky, 1950 (1967) (p.137); Tanaka, 1956 c (p.387); Fleminger, 1983 (p.607, 615); Brodsky & al., 1983 (p.303, spp. Key) ; Razouls, 1982 (p.147); Schulz, 1989 (p.195, Rem.); Razouls, 1993 (p.311); Bradford-Grieve, 1994 (p.98, Def.); Mauchline, 1998 (p.87: F; p.90: M); Boxshall & Halsey, 2004 (p.198); Vives & Shmeleva, 2007 (p.821, spp. Key); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | type: Mimocalanus. cultrifer Farran,1908. Total: 10 spp.
Diagnosis from Bradford-Grieve (1994, p.95) :
- As for the family definition.
- Male anal segment not reduced.
- Caudal rami symmetrical, caudal setae usually incomplete on specimens examined : no trace of outer (6th) seta, outer and inner terminal setae usually completely detached, middle 2 setae usually broken at bases, inner seta directed ventrally, small or reduced.
- Rostrum ansent.
- Female A1 with segments 8-9 partly or completely fused, limb narrowing between segments 9 and 11.
- Male A1 with segment 9 partly fused with segment 10, right segment 20 merging with segment 21, as in other left-handed species, limb narrowing between segments 8-10, segment 25 not reduced.
- A2 exopod exceeding the length on the endopod by 20%, exopodal segment 1 without setae, exopodal segments 2 and 3 separate, exopodal segment 2 without setae.
- Md basis with 2 or 3 inner setae, surface nude, endopodal segment 1 surface nude.
- Mx1 inner lobe 1 (praecoxa arthrite) proximal surface without spinules, distal anterior seta present ; inner lobe 2 (coxal endite) with 4 setae, inner lobes 2 and 3 (basal endite) relatively large ; proximal part of the endopod with 3 or 7 proximal and 3 or 7 distal setae ; small nude outer lobe 2 (basal exite) sometimes present.
- Mx2 length twice its wide, without a proximal hump ; lobe 5 with 3 strong and 1 reduced setae, posterior surface of its base without spines ; lobe 6 reduced, apparently with only 1 seta ; endopod segments reduced ; outer seta apparently absent.
- Mxp coxa and basis without a transverse spine-comb, coxa with a proximal group of 2 setae, midlength group of 3 setae, and distal group of 3 or 4 setae, basis with longitudinal row of reduced spinules or denticles, 3 setaze about midlength, and 2 distal setae, endopodal segments 4 and 5 with outer setae usually moderately long, with very long plumes on each side, endopodal segment 5 inner edge without spinules.
- P1 exopodal segment 1 without an outer distal spine ; exopodal segment 1 and 2 together longer than exopodal segment 3 ; the endopod with 1-2 inner-edge setae.
- P2-P4 exopod terminal seta with finely serrate outer edge (48-90 teeth), outer flange wide.
- P1 endopod with 1-2 setae ; P2 endopodal segment 2 with 0-1 outer setae.
- P3-P4 endopodal segment 3 with 0-1 outer-edge setae.
- Surfaces of the swimming-leg segments without spines, except coxa of female P4 which usually has a transverse row of thin setules on the posterior surface proximal to the inner seta.
- Males of some species have an inner seta on coxa of P4.
- Male P5 uniramous, left handed, variably asymmetrical ; exopod essentially 3-segmented, exopodal segments 1 and 2 may be partly fused. | | Remarks on dimensions and sex ratio: | | The mean female size is 2,082 mm (n= 10; S= 0,910; Cv= 0,437) and the mean male size is 1,770 mm (n= 6; S= 0,643; Cv= 0,369). The size ratio (M/F) is 0,902 ou 90,2 % (n= 6; S= 0,091; Cv= 0,101) | | | | | Syn.: | Oxycalanus Farran,1908 b (p.25); A. Scott, 1909 (p.33); Wolfenden, 1911 (p.220); Hypsicalanus Wolfenden, 1911 (p.219) | | Ref.: | Sars, 1905 b (p.8); A. Scott, 1909 (p.34); Sars, 1925 (p.37); Sewell, 1929 (p.95); Rose, 1933 a (p.87); Vervoort, 1946 (p.157); Farran & Vervoort, 1951 h (n°40, p.3); Tanaka, 1956 c (p.395); Grice,1971 (p.274); Damkaer, 1975 (Déf., p.14, 62, clé spp.); Razouls, 1982 (p.149) Brodsky & al., 1983 (p.324, clé spp.); Fleminger, 1983 (p.607, 610, 612, 615); Mauchline,1988 (p.724); Schulz, 1989 (p.195, Rem.); Ferrari, 1992 (p.392, tab.3); Razouls, 1993 (p.311); Bradford-Grieve, 1994 (p.99, Déf.); Mauchline, 1998 (p.78: F; p.80: M); Bradford-Grieve & al., 1999 (p.914: clé spp.); Boxshall & Halsey, 2004 (p.198) ; Vives & Shmeleva, 2007 (p.828, spp. Key); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | type: Monacilla typica Sars,1905. Total: 3 spp. + 1 iuncertained.
Diagnosis from Bradford-Grieve (1994, p.99) :
- As for the family definition.
- Head rounded or with a prominent crest.
- Rostrum 2-pointed, sometimes asymmetrical.
- Pedigerous segment 5 with lateral corners prolonged,
- Pedigerous segments 1-4 without lateral spinules.
- Prosome length 3.-3.5 times the length of the urosome.
- Genital segment sometimes asymmetrical, protruding ventrally 0.5 times the depth of the rest of the urosome.
- Anal segment reduced in the male.
- Caudal rami and setae symmetrical.
- Female A1 with segments 8-9 fused.
- Male A1 with segments 9-10 fused, right segment 20 merging with segment 21, segment 25 reduced.
- A2 exopod length exceeding the endopod by 20%, exopodal segment 2 probably with 2 setae anda t least partly fused with segment 3.
- Md basis with 4 setae.
- Mx1 inner lobe 1 as in Spinocalanus as the distal anterior set ais apparently absent ; inner lobe 2 with 5 setae, outer lobe 2 small with 1 short seta.
- Mx2 length 1.5 times its width with proximal outer hump nude and the outer seta apparently absent, lobe 5 with 4 strong setae and the postrior surface of its base without spines, lobe 6 small with 2 setae, endopodal segments not reduced.
- Mxp coxa with or, apparently without a transverse spine-comb at midlength on the outer anterior and posterior edge, basis without a spine-comb ; coxa with the proximal and midlength group of 3 setae each, distal group apparently with 4-6 setae ; basis with a longitudinal row of stiff hairs, 2 setae at about midlength, 1 seta more distal, and 2 distal setae ; endopodal segments 4 and 5 outer setae relatively short and thin, probably lightly plumose ; endopodal segment 5 inner edge without spinules.
- Male Mx1 and Mxp setae reduced in size.
- P1 exopodal segment 1 with outer distal spine, reduced in the male, and male with the distal outer spine on exopodal segment 2 forming a sharply curved hook ; exopodal segments 1 and 2 together longer than exopodal segment 3 which is apparently without posterior surface spines ; endopod with 3 inner setae.
- P2 endopodal segment 2 with 1 outer seta.
- P3 and P4 endopodal segment 3 with 2 outer-edge setae.
- Posterior surface of certain segments of the swimming legs with rows of spines.
- Male P5 biramous, asymmetrical, subequal, endopods 1-segmented, sometimes filiform ; exopods well built and 2- or 3-segmented. | | Remarks on dimensions and sex ratio: | | The mean female size is 2.188 mm (n = 28; SD = 0.1925), and the mean male size is 1.732 mm (n = 12; SD = 0.4078). The size ratio (male : female) is about 0.79. | | | | (7) Mospicalanus Schulz, 1996 | |
| | Ref.: | Schulz, 1996 (p.595); Boxshall & Halsey, 2004 (p.198); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | Type: Mospicalanus schielae Schulz,1996. Total: 1 sp.
Diagnosis adult female from Schulz (2996, p.595) :
- Posterior corner of prosome only slightly produced posteriorly to form rounded lobe;
- Urosome short, less than1/3 length of prosome.
- Rosytum absent.
- Exopod of A2 with 10 setae, with 2nd segment bearing 2 setae.
- Md gnathobase armed with closely spaced teeth; basis carrying 4 inner setae.
- Coxal endite of Mx1 with 5 and distal basal endite with 4 setae.
- Syncoxa of Mxp armed with 2, 3, and 3 setae; endopod with 2, 3, 3, 2, 3, and 4 setae; outer setae of both distal segments extremely long.
- P1 endopod 1-segmented, P2 2-segmented, P3 and P4 3-segmented.
- Exopods of P1 to P4 3-segmented.
- Swimming legs lacking spinulation on posterior faces, except basis of P4.
- P1 endopod with 2 inner setae; outer distal spine of 1st exopodal segment absent.
- Distal endopodal segments of P2-P4 armed with 0, 2, and 2 outer setae respectively.
- Female P5 absent.
For Schulz (1996, p.600) this genus exhibits a close relationship with Mimocalanus because of a number of further synapomorphies: 1- absence of the outer seta of the caudal rami; 2- the reduced setal numbers of the A2 exopod; (10 in Mospicalanus, 8 in Mimocalanus vs plesiomorphic 12 setae); 3- only 10 (vs 11) setae on the 2nd endopod segment of Md; 4- reduced number of armature elements (11 vs 15) on the praecoxal arthrite of Mx1, and only 5 setae (vs 6) on the coxal endite; 5- absence of an outer coxal seta of Mx2, and reduced setal numvers on basis and 1st endopod segment (3 and 1 vs 4 and 2, respectively); 6- lack of a praecoxal seta on Mxp; 7- absence of the outer distal spine on the 1st exopod segment of P1. | | | | (8) Rhinomaxillaris Grice & Hulsemann, 1967 | |
| | Ref.: | Grice & Hulsemann, 1967 (p.39, 40, Rem.); Razouls, 1982 (p.593); Fosshagen, 1983 (p.260); Razouls, 1993 (p.309); Schulz, 1996 (p.599, Rem.); Markhaseva & Kosobokova, 2001 (p.319, 323, Rem.); Boxshall & Halsey, 2004 (p.198); Andronov, 2014 (p.92, 155) | | Rem.: | Type: Rhinomaxillaris bathybia Grice & Hulsemann,1967. Total: 1 sp.
Formerly included in the Bathypontiidae family.
Diagnosis based on adult female from Grice & Hulsemann (1967, p.39) :
- Head and 1st thoracic segment incompletely separate, 4th and 5th thoracic segments inccompletely separate.
- Abdomen 4-segmented.
- Anal segment longer than preceding segment and caudal rami.
- Rostrum large and obtuse;
- A1 reaching to 5th thoracic segment; segments 8 and 9, 24 and 25 fused.
- Exopod of A2 7-segmented, about 2 times the length of endopod.
- Mx1 with 2 setae on 2nd inner lobe; 5 setae on endopod and 2 setae on exopod.
- Mx2 with 5 lobes bearing 1, 1, 3, 3, and 2 setae, respectively.
- Mxp large, with long and coarse setae distally.
- P1 exopod 3-segmented and endopod 1-segmented; 1st and 2nd exopodal segments without outer spines.
- Exopods of P2-P4 broken off.
- Endopods of P2 2- or 3-segmented, of P3 and P4 3-segmented.
- 3rd endopodal segments of P3 and P4 with 6 setae.
For Grice & Hulsemann (1967, p.39) it was difficult to assign the female species Rhinomaxillaris bathybia to an existing family. The new genus is distinguished from Bathypontia (= Temorites) by the long exopod in the A2, the shape and arrangement of the teeth on the Md blade, and the absence of P5.
Rhinomaxillaris differs from Zenkevitchiella and Foxtonia in the long exopod of A2, the Md blade and the peculiar rostrum.
From Zenkevitchiella, Rhinomaxillaris is further distinguished by the absence of external spines on the 1st and 2nd exopodal segments of P1, and the absence of P5.. It differs from Foxtonia in the increased number of segments in the endopods of P2 and P3 and in the absence of spinelike setae between the bases of A1.
Rhinomaxillaris also resembles certain genera in the family Pseudocalanidae. The large and blunt rostrum, the dfused 24th and 25th segments in A1, and the absence of external spines on the 1st and 2nd exopodal segments of P1 prevent Rhinomaxillaris from being placed in this family. | | | | Sognocalanus Fosshagen, 1967 | | Ref.: | Fosshagen, 1967 (p.308, 313); Razouls, 1982 (p.594); Fosshagen, 1983 (p.260); Razouls, 1993 (p.309); Schulz, 1996 (p.599, Rem.); Mauchline, 1998 (p.87: F; p.90: M); Markhaseva & Kosobokova, 2001 (p.319, 323, Rem.); Boxshall & Halsey, 2004 (p.198) | | Rem.: | Cf.
Arctokonstantinidae | | | | (9) Spinocalanus Giesbrecht, 1888 | |
| | Ref.: | Giesbrecht, 1892 (p.52, 209); Giesbrecht & Schmeil, 1898 (p.31); Sars, 1900 (p.75); 1901 a (1903) (p.22); Wolfenden, 1906 (p.43); Esterly, 1906 a (p.55); van Breemen, 1908 a (p.28); Wolfenden, 1911 (p.215); Rose, 1933 a (p.84); Vervoort, 1946 (p.146); Davis,1949 (p.21); Brodsky, 1950 (1967) (p.122, spp. key); Farran & Vervoort, 1951 g (n°39, p.3); Tanaka, 1956 c (p.389); Park, 1970 (p.482, spp. Key F); Grice, 1971 (p.275, 279, spp. Key); Damkaer, 1975 (p.15, spp. Key, Rev.); Gardner & Szabo, 1982 (p.191); Razouls, 1982 (p.137); Brodsky & al., 1983 ( p.247, spp. Key); Fleminger, 1983 (p.607, 615); Mauchline, 1988 (p.724, pores cuticulaires); Schulz, 1989 (p.195, Rem.); Razouls, 1993 (p.311); Bradford-Grieve, 1994 (p.100, Def.); Mauchline, 1998 (p.78: F, p.80: M, figs.167, 169); Bradford-Grieve & al., 1999 (p.914: clé spp.); Boxshall & Halsey, 2004 (p.198); Vives & Shmeleva, 2007 (p.831, spp. Key); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | type: S. abyssalis Giesbrecht,1888. Total: 29 spp. + 4 unidentified.
Diagnosis from Bradford-Grieve (1994, p.100) :
- As for the family definition.
- Rostrum absent.
- Female prosome in dorsal view slender to robust ovoid, lateral corners of pedigerous segment 5 sometimes prolonged, pedigerous segments 1-4 sometimes with lateral spinules, sometimes left and right sides with variable numbers.
- Male prosome in dorsal view usually not ovoid, anterior part irregular or rounded, with a central knob-like protrusion, posterior part usually somewhat rectangular, head usually expanded dorsally to nearly cover pedigerous segment 1, head and pedigerous segment 1 separate or partly fused.
- Male anal segment usually reduced.
- Female caudal rami and caudal setae sometimes asymmetrical.
- Female A1 with segments 8-9 fused.
- Male A1 with segment 9 partly fused with segment 10, segment 25 usually reduced.
- A2 exopod length exceeds the endopod by 20%, exopodal segment 1 with 0-2 setae, exopodal segments 2 and 3 fused, exopod segment 2 with 2 setae
- Md basis with 4 inner setae, and usually with a distal row of long setules on the anterior surface, endopodal segment 1 also with a row of long setules.
- Mx1 inner lobe 1 posterior surface with proximal clusters of spinules and denticles, proximal posterior surface with 3 or 4 setae, distal anterior seta present or absent, inner lobe 2 with 6 setae, proximal part of the endopod with 4 proximal and 5 distal setae, outer lobe 2 absent.
- Mx2 length 1.5-1.75 times its width with a small to strong proximal outer hump usually armed with hairs, setules, and/or spinules, lobe 5 with 4 strong setae and its posterior surface may have spines; lobe 6 small with 2 setae (1 seta small); endopod segments not reduced; outer seta present or, apparently, absent.
- Mxp coxa sometimes with a transverse spine-comb at midlength on the outer and posterior edge, basis sometimes also with a transverse spine-comb at about 0.33 of its length on the anterior and posterior surfaces; coxa with 3 groups of 3 setae each; basis witha longitudinal row of stiff hairs or stiff setules, 2 setae at about midlength, 1 seta more distal, and 2 distal setae; the form of the outer setae on endopodal segments 4 and 5 varying with species.
- P1 exopodal segment 1 with an outer distal spine, exopodal segments 1 and 2 together longer than exopodal segment 3 which sometimes has posterior surface spines ; endopod with 1-3 inner setae.
- P2 endopodal segment 2 with 0-1 outer seta.
- P3 and P4 endopodal segment 3 usually with 2 outer setae.
- Posterior surface of certain segments of the swimming legs with rows of spines.
- P2-P4 exopodal segment 2 distal anterior surface with 1 row of spinules.
- P2 exopodal segment 2 posterior surface usually with 1 row of spines.
- P3 and P4 exopodal segment 2 posterior surface with 1-2 rows of spines.
- P2 and P3 exopodal segment 3 posterior surface usually with 1-3 rows of spines.
- P4 exopodal segment 3 with or without posterior surface spines or spinules.
- Terminal exopod seta of P2-P4 with a moderately coarse serrate outer edge (20-50 primary teeth), the outer flange narrow.
- Endopodal segment 2 of P2-P4 with posterior surface with 2 rows of spines, 1 row sometimes reduced to small spinules on a few species.
- Endopodal segment 3 of P3 with the posterior surface usually with 2 rows of spines, but at least 1 row of spines or denticles.
- Endopodal segment 3 of P4 posterior surface with 2 rows of spines..
- Coxa of P4 usually with a transverse row of setules on the posterior surface near the inner seta.
- Male swimming legs generally similar to those of the female with variable armature, rows of spines on the posterior surface of some segments in the female are sometimes in circular clusters in the male ; coxa of P4 without inner transverse row of setules or spinules on the posterior surface, with or without a seta.
- Male P5 biramous with styliform endopods ; or very asymmetrical uniramous, usually left-handed ; exopods usually 3-segmented. | | Remarks on dimensions and sex ratio: | | The mean female size is 1,954 mm (n= 25; S= 0,696; Cv= 0,356) and the mean male size is 2,032 mm (n= 19; S= 0,789; Cv= 0,388). The size ratio (M/F) is 0,919 or 91,9 % (n= 27; S= 0,097; Cv= 0,106). 0,953 or 95,3 % (n= 16; S= 0,15; Cv= 0,157). The sex-ratio (F/M) is 1,315. It is exceptional that the male size is superior to that of the females; the obtained mean size is from the sex ratio and is not confirmed in the ratio of the sizes (M/F) obtained from the paired female and male data. | | | | (10) Teneriforma Grice & Hulsemann, 1967 | |
| | Syn.: | Tanyrhinus Farran, 1936 a (p.85) | | Ref.: | Grice & Hulsemann, 1967 (p.22); Grice, 1971 (p.274); Damkaer, 1975 (p.15, 82, Rev.); Fosshagen, 1983 (p.260, 261, Rem.); Brodsky & al., 1983 (p.331); Fleminger,1983 (p.607, 615); Razouls,1982 (p.152); Schulz, 1989 (p.195, Rem.); Razouls, 1993 (p.311); Bradford-Grieve, 1994 (p.105, Def.); Mauchline, 1998 (p.87: F; p.90: M); Boxshall & Halsey, 2004 (p.198); Vives & Shmeleva, 2007 (p.846); Markhaseva, 2008 (p.15, Rem.) | | Rem.: | type: Tanyrhinus naso Farran,1936. Total: 4 spp.
Diagnosis from Bradford-Grieve (1994, p.105) :
- As for the family definition.
- All pedigerous segments separate.
- Pedigerous segment 5 prolonged, extending 0/33 times the length of the genital segment.
- Rostrum single, long, blunt cone directed backwards.
- Caudal rami symmetrical, length 2.0-2.5 times their width, longer than or equal to the anal segment.
- Male anal segment not reduced.
- A1 female extends to the anal segment, 24-segmented, segments 8-9 fused.
- Male A1 reaching the end of the caudal rami, 24-segmented, segments 8-8 fused, on the left segments 20 and 21 distinct, segment 25 not reduced.
- A2 exopod equals the endopod.; exopodal segment 1 with 1 seta, exopodal segment 2 and 3 fused, exopod segment 2 with 2 setae.
- Md with 3 setae on the inner margin of basis.
- Mx1 female with some reduction in inner lobe 3, basipod, and the endopod, where the size and number of setae are reduced; inner lobes 2 and 3 with 4 and 2 setae respectively.
- P1 basis without minute outer distal seta, with inner distal seta; exopodal segment 1 without an outer distal spine, exopodal segment 2 with an outer-edge spine; endopod with 2 terminal and 2-3 inner-edge setae.
- Surface of the swimming legs without spinules.
- Endopodal segment 2 of P2 with 1 outer-edge seta.
- endopodal segment 3 of P3 and P4 with 2 outer-edge setae.
- Female P5 absent.
- Male P5 very asymmetrical present on the left side as a uniramous, 5-segmented appendage, right basis reduced to a small knob. | | Remarks on dimensions and sex ratio: | | The mean female size is 0.965 mm (n = 8; SD = 0.1912), and the mean male is 0.830 mm. The size ratio (male : female) is about 0.87. | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 22, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|



;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
