|
|
 |
|
Calanoida ( Order ) |
|
|
|
Bathypontioidea ( Superfamily ) |
|
|
| |
| | | |
| Bathypontiidae Brodsky, 1950 ( Bathypontioidea ) | | Ref.: | Brodsky, 1950 (1967) (p.82, 414); Fosshagen, 1967 (p.313); Bradford, 1969 b (p.500, 503, Table 3); Andronov, 1974 a (p.1005); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.588); Fosshagen, 1983 (Rem.: p.260); Brodsky & al., 1983 (p.143, 146); Mauchline, 1988 (p.722: cuticular pores); Huys & Boxshall, 1991 (p.406); Razouls, 1993 (p.308); Bradford-Grieve & al., 1999 (p.886, 902, 904, 963); Bradford-Grieve, 1999 b (p.17: Def., Rem., fig.189); Ohtsuka & Huys, 2001 (p.445, 461); Boxshall & Halsey, 2004 (p.12, 14, 15; 49; 71: Def.; p.72: Key of Genera); Vives & Shmeleva, 2007 (p.393, Key G.); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, Biol. mol, phylogeny) ; Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm  | | Rem.: | This family has been created by Brodsky (1950) to include the genera Bathypontia (placed in the Parapontellidae by Sars in 1905) and Temorites (placed in the Temoridae by Sars in 1900, followed by Rose in 1933) and adds the genus Zenkevitchiella in 1955.
Hülsemann and Grice (1963) create the genus Foxtonia from an adult female and some male juvenile stages. This form closely matches the family Spinocalanidae, but the structure of the Mx1 and Mx2 divert from it. For these reasons and awaiting the description of an adult male this genus is placed with doubt in this family by the authors.
In 1967 Grice et Hülsemann describe a species under the new genus name Rhinomaxillaris. The female being not intact and the male unknown, a precise diagnosis of this genus was not given. If this species shows certain resemblances with the Clausocalanidae family, various characters surely divert.
Fosshagen (1967) describes the female and the male of a small bathypelagic form with carnivorous tendence. The genus Sognocalanus shows numerous kinships with the other genera already included in this family, but also with the Aetideidae and Monacilla (Spinocalanidae). The author considers this family heterogenous.
Bradford (1969) adds the genus Alloiopodus to this family from a description of a female of which the legs are broken (except P1) and seems to consider the Bathypontiidae as a well distinctive family.
For Fosshagen (1983) the genera divide into 2 groups:
1/: Alloiopodus, Bathypontia, Temorites, Zenkevitchiella.
2/: Damkaeria, Foxtonia, Rhinomaxillaris, Sognocalanus. This second group would belong to the superfamily of the Clausocalanoidea, including presumably Teneriforma (Spinocalanidae). The so constituted family would have kinships with the Spinocalanidae, and Schulz (1996, p.599) transfers Rhinomaxillaris and Sognocalanus to the Spinocalanidae. Foxtonia is included in the family Spinocalanidae by Boxshall et Halsey (2004, p.196).
Bradford-Grieve, 1999 b (p.17) confirms the validity and redefines the family. She considers the genus Bathypontia as a junior synonym of Temorites.
Taxonomic notes after Boxshall & Halsey (2004, p.72): This family is , with the exception of Zenkevitchiella, characterized by the reduction of the maxillulary palo and by the setation pattern of the maxilla. Zenkevitchiella exhibits a much more plesiomorphic condition in the mouthparts and the P5, and was temporarily included in the Bathypontiidae by Brodsky (1955) primarily on the basis of the 23-segmented A1 of the female. This is a weak character upon which to base family placement, especially since the A1 of Temorites are 24-segmented. However, the presence of a modified, curved, outer spine on the basis of P3 is a more significant synapomorphy since it is not shared with related families, such as the Spinocalanidae, but is a characteristic of all bathypontiid genera.
Definition after Bradford-Grieve (1999 b, p.17):
Female:
- head and 1st pediger segment separate, 4th and 5th thoracic segments separate, often produced backward with rounded or acute point.
- Rostrum usually without filaments, in the form of a blunt or short process.
- Urosome 4-segmented.
- Genital segment with slightly protruding ventral process.
- Caudal rami short with 4-5 setae.
- A1 23-segmented, segments 8-9 and 24-25 fused,; 24-segmented (Temotites discoveryae) with numerous aesthetascs proximally, amy be asymmetrically developed (Alloiopodus pinguis).
- A2 (based on Bathypontia) rami almost equal in length; basipods 1 and 2 with 1 and 2 setae, respectvely; endopod segment 1 with 1-2 setae just distal to midlength (1 seta small), segment 2 with 7+9 setae; exopod 7-segmented with setal formula 0, 1, 1, 1, 1, 1, 1+3; other genera have warious reductions in segment numbers and setation §exopod 6-segmented in Alloiopodus, Temorites discoveryae, Zenke vitchiella).
- Md (based on Bathypontia gnathobase with large tooth often separated by some distance from numerous small teeth; basipod 2 large with 3-4 setae; endopod segments 1 and 2 with 2-3 and 8-11 setae, respectively; exopod 5-segmented with setal formula 1, 1, 1, 1, 1-2, respectively; other genera with reduced setation on endopod (Alloiopodus endopod with 0, 3 setae; Temorites discoveryae 0, 1 setae)
- Mx1 (based on Bathypontia) inner lobe 1 (arthrite) with 9-10 long straight spines; inner lobes 2 and 3 and basipod reduced in size and setation with 1 seta each; endopod absent; exopod well-developed with 10 setae; outer lobe 2 absent; outer lobe 1 with 3-4 setae; other genera and species with variously reduced setation (Temorites discoveryae has the greatest reduction inner lobe 1 with 4 spines and setae; inner lobes 2-3, basipod, endopod, and outer lobes 1 and 2 devoid of setae, exopod with 6 setae.
- Mx2 (based on Bathypontia) large, lobes 1-5 with 0-1, 1, 2, 3, 3-4 setae, respectively (one of them enlarged); endopod with 6 enlarged, sometimes flattened, distally serrate seta; other genera with more or less modified setae and in some genera (e.g. Alloiopodus) basipod 2 elongated.
- Mxp (based on Bthypontia) with relatively short setae; basipod 2 with 3 setae at about midlength; endopod 6-segmented, segment 1 almost completely fused to basipod 2 with 1-2 setae, segments 2-5 with 4, 3-4, 2, 2, 2-4 setae, respectively.
Swimming leg1 basipod 1 (coxa) with 1 inner distal seta.
- P1-P4 with 3-segmented exopod; endopod of P1 1-2 segmented, P2-P4 3-segmented.
- OP1 exopod segments 1 and 2 with or without outer edge spines; exopod segment 3 of P2-P4 with 3 outer edge spines
- Basipod 2 (basis) of P3 with long thin gently curving outer spine extending to distal border of exopodal segment 1.
- Seta and spine formula of Bathypontia see table.
- P5 absent or uniramous; 3-segmented, with 2 terminal spines, no other armature present (bathypontia, Temorites); or biramous with 2-3-segmented exopod and 1-segmented endopod (Zenkevitchiella, Alloiopodus).
Male:
- A1 19-20-segmented, prehensile on right; terminal part composed of 4 narrow elongate segments, proximal segment of which is characteristically curved.
- Mouthparts similar to those of the female.
- P2 may be symmetrical or asymmetrically developed on each side with exopod segment 2 outer edge spine greatly elongate on one side.
- P5 3-5-segmented; endopod may or may not be present.
Key of genera after Boxshall & Halsey (2004, p.72) :
1 - Outer margin of endopod of P1 linear ........ 2.
1' - Outer margin of endopod of P1 with rounded lobe ........Zenkevitchiella.
2 - 1st and 2nd exopodal segments of P1 with outer spines ............ Alloiopodus.
2' - 1st and 2nd exopodal segments of P1 without outer spines ........Temorites. | 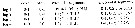 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.71]. Armature of swimming legs P1 to P4. Setation sometimes modified or reduced. Outer basal element of P3 enlarged and forming curved spine. Exopodal spines sometimes enlarged on P2 on one side only in males. Nota: Female P5 biramous in Zenkevitchiella and Alloiopodus; comprising 2-segmented protopod, 2 or possibly 3-segmented exopod and 1-segmented endopod bearing 3 to 5 distal and inner elements. Proximal exopodal segment double in Zenkevitchiella, due to fusion of 1st and 2nd exopodal segments, armed with 2 outer elements and 1 inner element; distal segment with 3 elements.Female P5 typically uniramous, comprising coxa, unarmed basis and 1-segmented exopod armed with 2 apical elements. - Male P5 asymmetrical, biramous or, typically uniramous; right leg larger than left; right leg 5-segmented in some Temorites species, including 3-segmented exopod. Endopod usually absent or represented by small knob. Exopod typically ending in spine which is frequently incorporated into distal segment. Exopod sometimes 2-segmented. Basis and proximal exopodal segment often ornamented with long spinules in Temorites species. Left leg with 2-segmented endopod in some Zenkevitchiella; usually uniramous, 4 or 5-segmented, including 3 exopodal segments; distal exopodal segment typically spiniform. - Eggs release into water |
 Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, Part I, No 166. [p.73, Fig. 8]. Bathypontiidae. A, Temorites elongata (as Bathypontia elongata) habitus female (dorsal); B, female Mx1; C, female Mx2; D, P5; E, Temorites brevis habitus male (lateral view); F, male P5; G, female P3; H, Zenkevitchiella abyssalis female P5. [Sars, 1924: A-D; Sars, 1900: E-G; Brodsky, 1955: H]. |
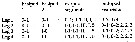 Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.17]. Armature of the swimming legs P1 to P4 after the genus bathypontia. | | | | | (1) Alloiopodus Bradford, 1969 | |
| | Ref.: | Bradford, 1969 b (p.499, Table 3, Rem.: p.503); Razouls, 1982 (p.594); Fosshagen, 1983 (p.260); Razouls, 1993 (p.308); Bradford-Grieve, 1999 b (p.20: Déf., Rem.); Bradford-Grieve, 2004 (p.287); Boxshall & Halsey, 2004 (p.72) | | Rem.: | Type: Alloiopodus pinguis Bradford, 1969. Total: 1 sp.
This genus, established from a female of which the legs were damaged, is placed by Bradford (p.500) among the Bathypontiidae family, defined by Brodsky, 1950 (1967, p. 414).
After Boxshall & Halsey (2004, p.72) the family is typically oceanic, bathypelagic forms and occur widely in tropical to temperate waters. The genus Alloiopodus is probably hyperbenthic.
Diagnosis from Bradford-Grieve (1999 b, p.20) :
- Head and pediger somite 1 separate, pediger somites 4 and 5 separate.
- Pediger somite 5 with asymmetrical posterior flaps.
- A1 very short, 23-segmented, unequal.
- Rostrum bifurcate.
- A2 exopod and endopod equal in length, endopod twice the width of exopod.
- Md gnathobase with 1 tooth large and set apart from others.
- Mx1 reduced, without an endopod or setae on outer lobe 1.
- Mx2 well developed with lobe 4 bearing a strong seta.
- Mxp with a gripping region between coxa and basis.
- P1 endopod 1-segmented.
- P2 endopod 3-segmented; (endopods of P2 and P4 unknown).
- P1 exopod 3-segmented (exopods of other pairs unknown).
- Basis of P3-P5 bearing a stout external spine.
- P5 endopod 1-segmented, exopod not known.
Bradford (1969 b, p.500) places this genus in the Bathypontiidae because of the degree of fit to Brodsky's (1950) [1967] definition of the family and the general similarity of its mouthparts, especially the reduction of setation on Mx1, to the other genera in the family.. As pointed out by other authors (Fosshagen, 1967; Grice & Hulsemann, 1967) there are many combinations of characteristics of rostrum, P5, reduction of Mx1 parts and segmentation of the limbs.
Bradford (1969 b, p.503) underline that this genus is the first benthic bathypontiid to be described as such, but the stoutness of the external spines on the swimming leg exopods of any other genera as Sognocalanus, Zenkevitchiella show that these could be benthic genera too. Some other genera, e.g. Aetideopsis spp., also look as if they live associated with the bottom in some way. Matthews (1964) , used plankton samplers to collect copepods from on or near the bottom, found that calanoud copepods are modified for a hyperbenyjonic life: fewer eggs than pelagic species; gradation of characters which depend on how closely the species is associated with the bottom, for example, in truly benthic species there are fewest naupliar stages, attachment of eggs to the sea bed becomes less fortuitous 'some are self-adhesive, others are stuck down by the female) and there are the largest numbers of setae,often annulate, on antennulae. The characters most consistently present are compact body shape, robust long extarnal spines on swimming leg exopods and smaller numbers of setae on the mouthparts. | | Remarks on dimensions and sex ratio: | | The body length in 1 female is 3.050 mm. No male founded. | | | | | | Syn.: | Isocalanus Wolfenden,1906 (p.35); 1911 (p.347) | | Rem.: | Bradford-Grieve, 1999 b (p.21) considers this genus as a junior synonym of Temorites. | | | | Damkaeria Fosshagen, 1983 | | Ref.: | Fosshagen, 1983 (p.260, 261) | | Rem.: | This genus, included in the Bathypontiidae, could, together with Teneriforma (Spinocalanidae) and other genera, constitute a new family. Cf. Spinocalanidae. | | | | Foxtonia Hulsemann & Grice, 1963 (? Bathypontiidae ) | | Ref.: | Hulsemann & Grice, 1963 (p.733); Razouls, 1982 (p.593); Fosshagen, 1983 (p.260); Razouls, 1993 (p.308) | | Rem.: | Provisionally included in the family Bathypontiidae. For Schulz (1996, p.599) this genus could be included in the Spinocalanidae. Position followed by Boxshall & Halsey, 2004 (p.196, Rem.). Cf. Spinocalanidae. | | | | | Syn.: | Isocalanus Wolfenden, 1906 (p.35); 1911 (p.347);
Temoropsis Wolfenden, 1911 (p.355);
Bathypontia Sars, 1905 c (p.23); A. Scott, 1909 (p.182); Sars, 1925 (p.355); Sewell, 1932 (p.391); Wilson, 1932 a (p.549); Rose, 1933 a (p.266); Brodsky, 1950 (1967) (p.415); Tanaka, 1965 (p.379); Grice & Hulsemann, 1965 (p.249); Bradford, 1972 (p.30); Deevey, 1973 a (p.357, 359, Rev., spp. Key); 1979 (p.724, spp.F Key); Razouls, 1982 (p.588); Fosshagen, 1983 (p.260); Mauchline, 1988 (p.722, 740); Razouls, 1993 (p.308); Mauchline, 1998 (p.95); Bradford-Grieve & al., 1999 (p.964, spp. Key); Bradford-Grieve, 2004 (p.287, Rem.) | | Ref.: | Sars, 1900 (p.100); Van Breemen, 1908 a (p.97); Rose, 1933 a (p.171); Brodsky, 1950 (1967) (p.416); Tanaka, 1965 (p.16); Razouls, 1982 (p.404, 592); Fosshagen, 1983 (p.260); Razouls, 1993 (p.309); Mauchline, 1998 (p.76); Bradford-Grieve, 1999 b (p.21: Def., Rem.); Boxshall & Halsey, 2004 (p.72); Vives & Shmeleva, 2007 (p.394, spp. Key) | | Rem.: | Type: Temorites brevis Sars, 1900. For Bradford-Grieve (1999 b, p.21) the type species is Bathypontia elongata Sars, 1905. Total: 15 spp. + 1 unidentified.
Diagnosis after Bradford-Grieve (1999 b, 21): As in the family with the following additional characters :
- Head separate or partly fused with 1st pedigerous , 4th and 5th segment usually separate.
- Posterior border of last thoracic segment asymmetrically extended in some species.
- Rostrum large, formed of a stumpy or rounded plate which is sometimes bifid at its tip, without filaments.
- Female urosome 4-segmented, 5-segmented in male, caudal rami usually short.
- A1 22-23-segmented ; male right A1 modified, 19-20-segmented.
- A2 and Md palp with equal rami.
- Mx1 with a well-developed exopod, endopod absent.
- Mx2 strong, with 6 or more long curved spines, often serrulate distally.
- Mxp weak.
P1 endopod 1-2-segmented, exopod segments 1 and 2 without outer edge spines.
- P2 with 2-3-segmented endopods.P1-P4 with 3-segmented exopods..
- P2 with outer edge spine of right exopodal segment 2 much larger than other spines in some species, especially in males. Marked asymmetry of P2, due to enlargement of this spine, occurs most notably in male B. spinifera, and to a lesser extent in male B. elongata, B . elegans, B. longicornis, B. similis ; P2 symmetrical in male B. minor
- P3 and P4 basis with an outer spine which is long on P3.
- Female P5 3-segmented, usually symmetrical, with an apical spine and usually 1 small external spine.
- Male P5 of 4-5 segments, in some species almost symmetrical, in others notably asymmetrical, the left leg sometimes with an inner plumose seta on basis. | | Remarks on dimensions and sex ratio: | | The mean female size is 3.198 mm (n = 22; SD = 1.5272), and in male 3.162 mm (n = 13; SD = 1.3392). The size ratio (male: Female) is 0.989. The sex ratio (Female: Male) is 1.75. | | | | | (3) Zenkevitchiella Brodsky, 1955 | |
| | Ref.: | Brodsky, 1955 a (p.205); Razouls, 1982 (p.593); Fosshagen, 1983 (p.260); Brodsky & al., 1983 (p.143, 146); Razouls, 1993 (p.309); Mauchline, 1998 (p.77); Bradford-Grieve, 1999 b (p.22, Def.); Boxshall & Halsey, 2004 (p.72) | | Rem.: | Type: Zenkevitchiella abyssalis Brodsky, 1955. Total: 4 spp.
Diagnosis after Bradford-Grieve (1999 b, p.22): As in the family definition with the following additional characters :
- Head and 1st pedigerous segment fused or separate, 4th and 5th thoracic segments separate.
- Rostrum with 2 filaments.
- A1 23-segmented in female ; male right A1 geniculate, 18- or 19-segmented.
- A2 rami of equal length.
- Mx1 may have all lobes well developed or have inner lobes 2, 3 and endopod reduced.
- Mx2 with strong bent setae, not reduced on proximal lobes.
- Exopod of P1-P4 3-segmented.
- Endopod of P1 1- or 2-segmented.
- Endopod of P2-P4 3-segmented.
- Exopodal segments 1-3 of P1 with 1, 1, 2 outer edge spines resoectively.
- Basis of P3 and P4 with 1 outer edge spine.
- Female P5 biramous, endopods 1-segmented, exopod 2- or 3-segmented, with 1 inner distal spine on penultimate segment.
- Male P5 biramous, endopods 1- or 2-segmented or rudimentary, left exopod 1-segmented, right exopod 2- or 3-segmented. | | Remarks on dimensions and sex ratio: | | The mean female size is 1.200 mm (n = 4; SD = 0.7836), and in male 0.780 mm (n : 2; SD = 0.0778). Only one species shows measurements male in the two sexes, the size ratio (male: female) is 0.890. The sex ratio (female: male) is 1.5. | | | | | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed December 03, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|



;)
{kind=link}
{kind=link}
{kind=link}
