|
|
 |
|
Calanoida ( Order ) |
|
|
|
Calanoidea ( Superfamily ) |
|
|
|
Paracalanidae ( Family ) |
|
|
|
Paracalanus ( Genus ) |
|
|
| |
Paracalanus aculeatus Giesbrecht, 1888 (F,M) | |
| | | | | | | Syn.: | Paracalanus parvus : T. Scott, 1894 b (p.26, figs.F,M);
Acrocalanus pediger (M) Cleve, 1901 (p.35, figs.M);
Paracalanus aculeatus plumulosus Wolfenden, 1905 (1906) (p.999, figs.F);
No Paracalanus clevei Carl, 1907 (p.7, 16); Früchtl, 1923 (p.451); 1924 b (p.35, fig.F: anomaly) | | | | Ref.: | | | Giesbrecht, 1892 (p.164, 170, figs.F); Giesbrecht & Schmeil, 1898 (p.24); A. Scott, 1909 (p.26); Wolfenden, 1911 (p.203); Sewell, 1912 (p.326, 328, 353, 358); Pesta, 1912 a (p.44, figs.F,M); 1913 (p.31); Sewell, 1914 a (p.204, Rem.M); Früchtl, 1924 b (p.20); Sars, 1925 (p.24); Farran, 1929 (p.208, 222); Sewell, 1929 (p.38, 62, figs.F, Rem.: 2 forms); Mori, 1932 a (p.167, figs.M); Rose, 1933 a (p.74, figs.F); Dakin & Colefax, 1933 (p.204); Farran, 1936 a (p.79, fig.F); Mori, 1937 (1964) (p.30, figs.F,M); Dakin & Colefax, 1940 (p.95, figs.F,M); Wilson, 1942 a (p.201, fig.F); Vervoort, 1946 (p.127, Rem.); Sewell, 1947 (p.51); 1951 (p.358, figs.F,M, Rem.: parasites); Farran & Vervoort, 1951 c (n°35, p.3, fig.F); Carvalho, 1952 a (p.142, figs.F); Shen & Bai, 1956 (p.219: Rem.); Tanaka, 1956 c (p.367, Rem.F); Vervoort, 1957 (p.35, Rem.); Tanaka, 1960 (p.22, Rem.); Grice, 1962 (p.183, figs.F, Rem.); Brodsky, 1962 c (p.114, figs.F); Kasturirangan, 1963 (p.24, figs.F,M); Vervoort, 1963 b (p.103, figs.F,M, Rem.); Unterüberbacher, 1964 (p.17); Gonzalez & Bowman, 1965 (p.243, figs.F); Chen & Zhang, 1965 (p.124, 129: Rem.); Owre & Foyo, 1967 (p.39, figs.F,M); Wellershaus, 1969 (p.247); Ramirez, 1971 (p.81, fig.F); Kos, 1972 (Vol I, figs.F,M, Rem.); Marques, 1973 (p.237); 1974 (p.13); 1976 (p.988); Greenwood, 1976 (p.20); Björnberg, 1980 (p.66, figs.F); Björnberg & al., 1981 (p.622, figs.F,M); Marques, 1982 (p.755); Campaner, 1985 (p.10); Nishida, 1989 (p.173, fig. 3, table 2, 3, dorsal hump); He & al., 1992 (p.250); Bradford-Grieve, 1994 (p.67, figs.F,M); Chihara & Murano, 1997 (p.844, Pl.135: F,M); Bradford-Grieve & al., 1999 (p.877, 910, figs.F,M); Paffenhöfer & Loyd, 2000 (p.171, fig.F); Conway & al., 2003 (p.160, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.152, figs.F,M); Mulyadi, 2004 (p.171, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.966, figs.F,M, Rem.); Phukham, 2008 (p.119, figs.F, M); Kesarkar & Anil, 2010 (Rem.: p.403); Soh & al., 2013 (p.63, figs.F) | 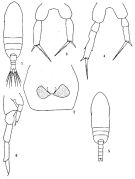 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.9, 1-6]. Female (from E China Sea): 1, habitus (dorsal); 2, genital segment (ventral); 3, P5 (posterior); 4, P5 (posterior, from another specimen, abnormal). Male: 5, habitus (dorsal); 6, P5 (posterior).
|
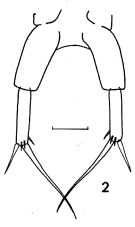 issued from : F.C. Ramirez in Revta Mus. La Plata, Seccion Zool., 1971, XI. [Lam.I, Fig.2]. Female (from off Mar del Plata): 2, P5. Scale bar in mm: 0.02.
|
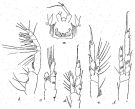 issued from : R.B.S. Sewell in Mem. Indian Mus., 1929, X. [p.63, Fig.20]. As Paracalanus aculeatus forma major. Female (from N Indian Ocean): a, posterior end of thoracic segment and P5 and genital segment (ventral); b, Mxp; c, P1; d, P2; e, P3; f, P4.
|
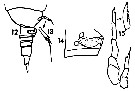 issued from : T.K.S. Björnberg in Bolm Inst. oceanogr., Sao Paulo. [p.66, Figs.12-14]. (from S Brazil): 12, last thoracic segment and urosome (lateral right side); 13, terminal segment of P5; 14, genital segment (lateral). Male: P5.
|
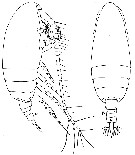 issued from : W. Vervoort in Atlantide Report., 1963, 7. [p.104, Fig.6]. Female (from off Ghana): a-b, habitus (lateral and dorsal, respectively) (x88).
|
 issued from : W. Vervoort in Atlantide Report., 1963, 7. [p.110, Fig.11]. Female: a-d, P1 to P4 (x220); e, genital somite (ventral) (x240); (x220); f, P5 (x375). Male: g, P5 (x375)
|
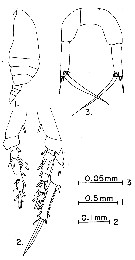 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.184, Pl.4, Figs.1-3]. Female (from equatorial Pacific): 1, habitus (lateral); 2, P4 (one exopod missing); 3, P5. Nota: All specimens belong to P. aculeatus f. major Sewell, 1929.
|
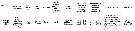 issued from : K.S. Kesarkar & A.C. Anil in J. mar. Biol. Ass. UK, 2010, 90 (2). [p.403, Table 2]. Characteristics of females.
|
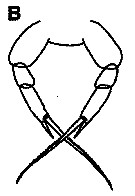 issued from : K.S. Kesarkar & A.C. Anil in J. mar. Biol. Ass. UK, 2010, 90 (2); [p.407, Fig.5, B]. Female: B, P5.
|
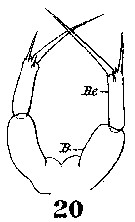 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.9, Fig.20]. Female: 20, P5. B = basipod; Re = exopod.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.26, 30]. Female: 26, P3 (posterior view); 30, P4 (posterior view). B2 = basis; Re = exopod; Ri = endopod; St = terminal spine; Si = inner seta; Se = outer seta.
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. [p.38, Figs.203, 204]. Female: 203, P4; 204, P5.
|
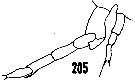 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. [p.38, Fig.205]. Male: 205, P5.
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.202, Fig.76]. Female (from W Malay Peninsula): a-b, habitus (dorsal and lateral, respectively); c, urosome; d, P4; e, P5. Male: f-g, habitus (dorsal and lateral, respectively); h, P5. Body length after drawings: F = 0.838 mm; M = 1,920 mm.
|
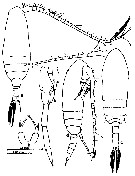 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.172, Fig.95]. Female (from Indonesian Seas): a, habitus (dorsal); b, P5. Male: c, habitus (dorsal); d, same (lateral); e, P5. Nota female: Prosome 3.5 times as long as urosome, without dorsal hump. Cephalon rather vaulted. Rostral filament filiform. A1 longer than the body. Mxp, 3rd lobe of coxa with 3 setae. P2-P4, coxa without surface spinules; endopodite 2 with prominent spinules on posterior surface, posterior surface spines on endopodite 2 more prominent than those of P. parvus. P5, inner terminal spine 2.5-3.0 times as long as outer terminal spine, considerably longer than distal segment. Innermost caudal seta more than twice as long as its own ramus.
|
 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.65, Fig.37]. Female (from Korean waters): A-B, habitus (dorsal and lateral, respectively); C, forehead (ventral view); D, head (lateral); E, corner of last thoracic segment and genital segment (dorsal); F, P5. Scale bars: A, B = 200 µm; C, E = 50 µm; D, 100 µm.
|
 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.66, Fig.38]. Female: A, P1; B, P2; C, P3; D, P4. Scale bar: 100 µm.
|
 Issued from : M.S. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Brodsky, 1962. Female: 1-2, habitusn (dorsal and lateral, respectively); 3, P3; 4, outer margin of exopodal segment 3 of P3; 5, P4; 6, P5; 7, seminal vesicle. Male: 8, habitus (lateral); 9, P5. For Brodsky, the total length of P. aculeatus minor is 0.7-0.8 mm and for P. aculeatus major: F = 0.9-1,4 mm; male = 0,9-1,4 mm.
| | | | | Compl. Ref.: | | | Cleve, 1904 a (p.194); (Carl, 1907 (p.16); Rose, 1934 (p.145); C.B. Wilson, 1950 (p.275); Krishnaswamy, 1953 (p.114); Chiba & al., 1957 (p.306); 1957 a (p.11); Kott, 1957 (p.6, 13); Yamazi, 1958 (p.148, Rem.); Wickstead, 1959 (p.69, Rem.: p.71); Fagetti, 1962 (p.11); Cervigon, 1962 (p.181, tables: abundance distribution); Fish, 1962 (p.10); Grice & Hart, 1962 (p.287, 293: Rem.); Gaudy, 1962 (p.93, 99, Rem.: p.103); Ganapati & Shanthakumari, 1962 (p.7, 15); Shmeleva, 1963 (p.141); Gaudy, 1963 (p.21, Rem.); Björnberg, 1963 (p.25, Rem.); Unterüberbacher, 1964 (p.17); De Decker, 1964 (p.16, 23, 29); De Decker & Mombeck, 1964 (p.13); Grice & Hulsemann, 1965 (p.223); Anraku & Azeta, 1965 (p.13, Table 2, fish predator); Chen & Zhang, 1965 (p.41, figs.F,M); Shmeleva, 1965 b (p.1350, lengths-volume-weight relation); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 10); Neto & Paiva, 1966 (p.20, Table III, annual cycle); Mazza, 1966 (p.69); Pavlova, 1966 (p.43); Grice & Hulsemann, 1967 (p.14); Fleminger, 1967 a (tabl.1); De Decker, 1968 (p.45); Vinogradov, 1968 (1970) (p.268); Delalo, 1968 (p.137); Dowidar & El-Maghraby, 1970 (p.269); Park, 1970 (p.475); Deevey, 1971 (p.224); Timonin, 1971 (p.281, trophic group); Apostolopoulou, 1972 (p.327, 337); Binet & al., 1972 (p.71); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %, Table IV); Patel, 1975 (p.659); Timonin, 1976 (p.79, vertical distribution); Carter, 1977 (1978) (p.35); Timonin & Voronina, 1977 (p.286, fig.5); Ikeda, 1977 d (p.263, feeding); Frontier, 1977 a (p.14); Binet, 1979 (p.400); Dessier, 1979 (p.132, 201, 204); Chen Q-c, 1980 (p.795); Porumb, 1980 (p.168); Sreekumaran Nair & al., 1981 (p.493, fig.2cont.), Vives, 1982 (p.290); Brenning, 1982 (p.1, spatial distribution, T-S diagram, Rem.); Kovalev & Shmeleva, 1982 (p.82); Smith S.L., 1982 (p.1331, abundance, monsoon effect); Guangshan & Honglin, 1984 (p.118, tab.); Stephen, 1984 (p.161, Distribution vs. thermocline & geographic); De Decker, 1984 (p.315); Binet, 1984 (tab.3); 1985 (p.85, tab.3); Brenning, 1985 a (p.23, Table 2); Kinzer & Schulz, 1985 (tab.4); Madhupratap & Haridas, 1986 (p.105, tab.1); Renon, 1987 (tab.2); Mitra & al., 1990 (fig.3); Othman & al., 1990 (p.561, 563, Table 1); Hirakawa & al., 1990 (tab.3); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Hirakawa, 1991 (p.376: fig.2); McKinnon, 1991 (p.471); Dai & al., 1991 (tab.1); Yoo, 1991 (tab.1); Suarez, 1992 (App.1); Gajbhiye & Abidi, 1993 (p.137); Shih & Young, 1995 (p.71); Roff & al., 1995 (p.165, Table 3, 5; nauplii, copepodites growth rates, bacteriovory); Webber & al., 1996 (tab.1); Kotani & al., 1996 (tab.2); Bundy & Paffenhöfer, 1996 (p.113, flow fields); Go & al., 1997 (tab.1); Paffenhöfer & al., 1996 (p.1699, motion behavior); Suarez-Morales & Gasca, 1997 (p.1525); Ramaiah & Nair, 1997 (tab.1); Park & Choi, 1997 (Appendix); Hure & Krsinic, 1998 (p.26, 100); Nair & Ramaiah, 1998 (p.272, fig.4); Noda & al., 1998 (p.55, Table 3, occurrence); Lopes & al., 1998 (p.195); Alvarez-Cadena & al., 1998 (tab.1,2,3,4); Hopcroft & al., 1998 (tab.2); Hsieh & Chiu, 1998 (tab.2); Mauchline, 1998 (tab.30, 47, 51, 62, 63); Wong & al, 1998 (tab.2); Smith S. & al., 1998 (p.2369, Table 6, moonsoon effects); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p.110); El-Serehy, 1999 (p.172, Table 1, occurrence); Dolganova & al., 1999 (p.13, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Lopes & al., 1999 (p.215, tab.1); Razouls & al., 2000 (p.343, Appendix); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Alvarez-Silva & Gomez-Aguirre, 2000 (p.163: tab.2); Ueda & al., 2000 (tab.1); Dalal & Goswami, 2001 (p.22, fig.2); McKinnon & Duggan, 2001 (p.121); Lo & al., 2001 (1139, tab.I); Sameoto al., 2002 (p.13); Dunbar & Webber, 2003 (tab.1); McKinnon & al., 2003 (p.101, tab.2, fig.11); Hsiao & al., 2004 (p.326, tab.1); Hsieh & al., 2004 (p.397, tab.1, p.399, tab.2); Rezai & al., 2004 (p.486, tab.2, 3, abundance, Rem.); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Krumme & Liang, 2004 (p.407, tab.1); Daly Yahia & al., 2004 (p.366, fig.4); Lan & al., 2004 (p.332, tab.1, tab.2); Kang & al., 2004 (p.1524, fig.7); Shimode & Shirayama, 2004 (p.607, tab. 2); Yin & al., 2004 (p.3); Lo & al., 2004 (p.89, tab.1); Kazmi, 2004 (p.230); Dias & Bonecker, 2005 (p.100 + poster); Obuid Allah & al., 2005 (p.123, occurrence % vs metal contamination); Wiggert al., 2005 (p.1013: feeding behavior); Smith & Madhupratap, 2005 (p.214, tab.4,6); Rezai & al., 2005 (p.157, Table 2, 5: spatial & temporal variations); Prusova & Smith, 2005 (p.75, 78); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño, Rem.: p.69, 72, 73, 74); Zuo & al., 2006 (p.163: tab.1, 3, figG;8: stations group); Hwang & al., 2006 (p.943, tabl. I); Araujo, 2006 (p.173, Tab.3): Dias & Araujo, 2006 (p.61, Rem., chart); Dur & al., 2007 (p.197, Table IV); Khelifi-Touhami & al., 2007 (p.327, Table 1); Madhu & al., 2007 (p.54, Table 4, abundance vs monsoon); Lan Y.-C. & al., 2008 (p.61, Table 1, % vs stations, Table 2: indicator species, Rem.: p.71); Tseng L.-C. & al., 2008 (p.153, Table 2, fig.5, occurrence vs geographic distribution, indicator species); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons, table 3: indicator species); Jitlang & al., 2008 (p.65, Table 1); McKinnon & al., 2008 (p.844: Tab.1, p.846: Tab.II, fig.7, f. major and f. minor); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.514, 521); Fernandes, 2008 (p.465, Tabl.2); Wishner & al., 2008 (p.163, Table 2, fig.8, oxycline); Shmeleva & al., 2008 (p.31, Table 1); Ohtsuka & al., 2008 (p.115, Table 5); Rossi, 2008 (p.90: Tableau XII); Pagano, 2009 (p.116); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Skovgaard & Salomonsen, 2009 (p.425, Table 2); Tseng & al., 2009 (p.327, fig.5, feeding); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Hwang & al., 2009 (p.49, fig.4, 5); Chiba & al., 2009 (p.1846, Table 1, occurrence vs temperature change); Gusmao & McKinnon, 2009 (p.1101, sex ratios); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Cornils & al., 2010 (p.2076, Table 3, Fig.5); Sun & al., 2010 (p.1006, Table 2); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Mazzocchi & Di Capua, 2010 (p.426); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Fazeli & al., 2010 (p.153, Table 1, as aculatus); W.-B. Chang & al., 2010 (p.735, Table 2, 3, 4, fig.5, abundance); Hsiao S.H. & al., 2011 (p.475, Appendix I); Hsiao & al., 2011 (p.317, Table 2, 3, fig.6, indicator of seasonal change); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Tseng L.-C. & al., 2011 (p.239, Table 3, abundance %); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Zhang G.-T. & Wong, 2011 (p.277, fig.6, 7, 8, abundance, indicator); Zhaoli & al., 2011 (p.84, abundance %); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Chen H; & al., 2011 (p.84, spatial & temporal variations); Chew & Chong, 2011 (p.127, Table 3, abundance vs location); Beltrao & al., 2011 (p.47, Table 1, density vs time); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Huo & al., 2012 (p.1, Table 1: dominance); Moon & al., 2012 (p.1, Table 1); Gubanova & al., 2013 (in press, p.4, Table 2); Cornils & Blanco-Bercial, 2013 (p.861, Table 1, molecular analysis, figs.3, 4, 5); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Tseng & al., 2013 (p.507, seasonal abundance); Jagadeesan & al., 2013 (p.27, Table 3, seasonal variation); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Pansera & al., 2014 (p.221, Table 2, abundance); Mazzocchi & al., 2014 (p.64, Table 4, 5, abundance); Zaafa & al., 2014 (p.67, Table I, occurrence); Nakajima & al., 2015 (p.19, Table 3: abundance); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); Ohtsuka & Nishida, 2017 (p.565, Table 22.1); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Belmonte, 2018 (p.273, Table I: Italian zones); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Abo-Taleb & Gharib, 2018 (p.139, Table 5, occurrence %); Dias & al., 2018 a (p.189, Rem.: p.196); Palomares-Garcia & al., 2018 (p.178, fig.3: relative frequency, Table 1); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution). | | | | NZ: | 20 | | |
|
Distribution map of Paracalanus aculeatus by geographical zones
|
| | | | | | | | | | | | | | |  issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 9 ]. Paracalanus aculeatus (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 9 ]. Paracalanus aculeatus (from South Adriatic).
Dimensions, volume and Weight wet. Means for ca. 100 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
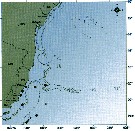 issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.61]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.61].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S).
Nota: sampling 13 specimens. |
 issued from : M.H. Bundy & C._A. Paffenhöfer in Mar. Ecol. Progr. Ser., 1996, 133. [p.103, Fig.2]. issued from : M.H. Bundy & C._A. Paffenhöfer in Mar. Ecol. Progr. Ser., 1996, 133. [p.103, Fig.2].
Feeding current of a tethered female Paracalanus aculeatus (cephalothorax length = 1.0 mm), collected off the coast of Georgia (USA), feed with Gymnodinium nelsoni and Rhodomonas sp. After acclimatation, the copepods were placed in complete darkness prior to videotaping in a 6 l cubic vessel, in the dark.
a: Flow field velocity. Contours represent fluid speeds and are plotted at 1 mm/sec intervals.
b: Flow field geometry. Vectors indicate direction and magnitude of flow.
Soingle large arrow represents copepod swimming trajectory. Isolines are plotted at 1 mm/sec intervals. |
 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 31. Jahrgang 1982. Mat.-nat. wiss. Reihe, 6. [p.1, Figs.1, 2]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 31. Jahrgang 1982. Mat.-nat. wiss. Reihe, 6. [p.1, Figs.1, 2].
Spatial distribution for Paracalanus aculeatus, and Paracalanus scotti (= Parvocalanus scotti) from 8° S - 26° N; 16°- 20° W, for diferent expeditions (V1: Dec. 1972- Jan. 1973; VI: May 74). |
 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 31. Jahrgang 1982. Mat.-nat. wiss. Reihe, 6. [p.1, Figs.1, 2]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 31. Jahrgang 1982. Mat.-nat. wiss. Reihe, 6. [p.1, Figs.1, 2].
T-S diagram for Paracalanus aculeatus, and Paracalanus scotti (= Parvocalanus scotti) from 8° S - 26° N; 16°- 20° W and Namibia for diferent expeditions (V1: Dec. 1972- Jan. 1973; VI: May 74 and VIII: 21/9-17/12 1976).
SO: Southern Surface Water (S °/oo: 34,50; T°C: 29,0); ND: Northern Water of the Surface Layer (S °/oo: 37,5; T°C: 21,0); SD: Southern Deep Water of the surface layer (S °/oo: 35,33; T°C: 13,4). See commentary in Temora stylifera and Brenning (1985 a, p.6). |
 Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6]. Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6].
Variations in the most abundant copepod species (mean ± SE) along the transect in the boundary waters between the northern part Taiwan Strait and the East China Sea i March (black bar) and October (grey bar) 2005 (Mann-Whitney U test, sig. *P <0.51.
See drawings of Hydrological conditions and superficial marine currents in Calanus sinicus. |
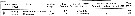 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5].
Summary observations on naupliar bacterovory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Kingston Harbour (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB. |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.172, Table 6]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.172, Table 6].
Summary observations on adult female and copepodites bacterivory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Kingston Harbour (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB. |
| | | | Loc: | | | sub-Antarct. (Indian), South Africa (E & W, off Cape of Good Hope), Namibia, Angola (Baia Farta), Congo, off Ascension Is., off E St. Paul Is., Atlant. (central equatorial), G. of Guinea, off Lagos, Ivorian shelf, Dakar, Cape Verde Is., off Morocco-Mauritania, off W Canary Is., Patagonia, Brazil (Paranagua, Rio de Janeiro, Guanabara Bay, Campos Basin, Vitoria-Cabo de Sao Tomé, Camamu, off Natal, Tocantins' mouth, Bragança: Caeté Bay), off Amazon, Barbados Is., Caribbean Colombia, Bahia de Mochima (Venezuela), Cariaco Basin, Caribbean Sea, Yucatan, Jamaica (Kingston Harbour), Yucatan (Ascension Bay), Cuba, E Costa Rica, G. of Mexico, Florida, off Bermuda, New York, off Nova Scotia E, off W Tangier, Medit. (Alboran Sea, Sidi Fredj coast, Gulf of Annaba, El Kala shelf, Marseille, Toulon Harbour, Ligurian Sea, Tyrrhenian Sea, Lake Faro (Sicily), Strait of Messina, Adriatic Sea, coast from Conero to Istria, Aegean Sea, Lebanon Basin, Alexandria, Black Sea, Black River estuary), Taba, Hurghada, Safâga, Red Sea, Gulf of Oman, G. of Aden, Arabian Sea, Arabian Gulf, UAE coast, Somalia, Maldive Is., Sri Lanka, Natal, Madagascar (Nosy Bé), India (Saurashtra coast, Bombay, Kerala, Lawson's Bay, Mandarmani), Indian, Madras, Gulf of Mannar, Palk Bay), Bay of Bengal, Andaman & Nicobar Is., Batten Island, S Burma, W Malay Peninsula Andaman Sea), Straits of Malacca, Sangga estuary (mainly nearshore and offshore), off Singapore, G. of Thailand, Indonesia-Malaysia, off Labuan (W Java), off Lonbok Sea, Tioman Is., Ambon Bay (S Ceram Is.), SW Celebes, Philippines, Viet-Nam ( Cauda Bay), Hainan (Sanya Bay), Hong Kong, China Seas (Yellow Sea, East China Sea, South China Sea, Xiamen Harbour, E Kuroshio Current, Hainan Is.), Taiwan Strait, Taiwan (S, E, W, SW, NW, off Danshuei River, N, Mienhua Canyon, NE), Okinawa, S & E Korea, Muan Bay, Japan Sea, Nagazaki, Japan (Ariake Bay, Kuchinoerabu Is., Tokyo Bay, Honshu: Suruga Bay), Bikini Is., Aleutian Is., Bering Sea, Pacif. (W equatorial), S Tasmania, Australia (SE, Townsville: inshore waters, Great Barrier Reef, Moreton Bay, Shark Bay, North West Cape, G. of Carpentaria, Darwin Harbour), New Caledonia, off New Zealand, Pacif. (equatorial), off California, Baja California (Bahia Magdalena), G. of California, La Paz, Acapulco Bay (rare), off E Hawaii, W Costa Rica, W Panama, Clipperton Is., off Ecuador, off NW Marquesas Is., off NE Tuamotu Is., Bahia Cupica (Colombia), Galapagos-Ecuador, off Peru, Chile
Data from Cornils & Blanco-Bercial (2013): 05°11'S; 143°16'W. | | | | N: | 317 ? | | | | Lg.: | | | (14) F: 1,15-0,99; M: 1,27-1,1; (28) M: 1,05-0,92; (34) F: 1,23-0,78; (35) F: 1,14-1,08; (47) F: 1,2-0,85; (55) F: 1,36-1,18; M: 1,36-1,27; (66) F: 1,21-0,78; (82) M: 1,2; (104) F: 1,25; M: 1,2; (150) F: 1,3-1,2; (151) M: 0,71; (172) F: 0,87; (179) F: 1,08-1,02; (237) F: 1,2; M: 1,2-1,0; (332) F: 1,36-0,9; M: 1,31; (333) F: 0,86; 0,82; M: 0,84; (334) F: 1,25; M: 1,2; (335) F: 1,14-0,9; (336) F: 1,18-0,89; M: 1,15-0,9; (432) F: 1,15-0,85; (785) F: 1,12-1,01; (795) F: 1,2; M: 1; (1112) F: 1,06-1,39; M: 1,01-1,25; (1122) F: 0,75; M: 1,05; (1174) F: 1,45; {F: 0,75-1,45; M: 0,71-1,36}
The mean female size is 1.094 mm (n = 42; SD : 0.1929), and the mean male size is 1.119 mm (n = 22; SD = 0.1757. The size ratio (male : female) is approximatively 1.023. | | | | Rem.: | epipelagic.
Paracalanus aculeatus major Sewell,1929 (F)
Ref.: Sewell, 1929 (p.63, figs.F); Farran, 1936 a (p.80, fig.F); Vervoort, 1946 (p.128); Tanaka, 1956 c (p.367); Silas, 1972 (p.645); Grice, 1972 (p.183, figs.F); Wellershaus, 1969 (p.247, Rem.); McKinnon & a., 2008 (p.844 : Tab.1)
Loc.: Bay of Bengal, Nicobar Is. (Nankauri Harbour), Australia (North West Cape), Indo-Malaysia, Pacif. (equatorial)
Lg.: (28) F: 1,23-0,9; (29) F: 0,99-0,89; (55) F: 1,36-1,18; M: 1,36-1,27; (101) F: 1,17-1
For Björnberg (1963, p.26) in Brazilian waters this species is the most frequent, though not numerous copepod in samples of water of high salinity (above 36.00 p.1000) and high temperature (above 20°C); in samples of high salinity and low temperatures (less than 20°C) it was not very frequent; it was present in samples from surface layers of low salinity (under 35.00 p.1000) and temperatures ranging from 18 to 28°C; it was dominant or subdominant among copepods in coastal and shelf waters of low salinity.
According to Mulyadi (2004, p.171) this species is rare in inshore Indnesian waters, but fairly common in offshore, which only very occasionally penetrates into deeper waters.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a fine-filter feeder.
Cf Calanus pacificus (figs 3,4 and Table 1, from Lopez-Ibarra & Palomares-Garcia).
See in DVP Conway & al., 2003 (version 1)
R. Stephen, 2007 : Data sheets of NIO, Kochi, India (on line). | | | Last update : 28/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 14, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
