|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Pontellidae ( Family ) |
|
|
|
Labidocera ( Genus ) |
|
|
| |
Labidocera trispinosa Esterly, 1905 (F,M) | |
| | | | | | | Syn.: | Labidocera tenuicauda (M) C.B. Wilson, 1950 (part., p.249, fig.M, no F); Andronov, 2014 (p.32, Fig.M). | | | | Ref.: | | | Esterly, 1905 (p.200, figs.F,M); Fleminger, 1965 (p.128, Rem.); 1975 (p.401 & suiv., figs.F,M); Silas & Pillai, 1973 (1976) (p.815); Dawson & Knatz, 1980 (p.8, figs.F,M); Sazhina, 1985 (p.71, figs.N); Huys & Boxshall, 1991 (p.436) | 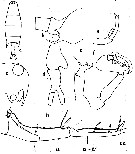 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.20 Fig.481. Female (from San Diego Region): d, urosome (dorsal); g, P5. Male: a, habitus (dorsal); b, lateral portion of last thoracic segment (lateral, right side); c, forehead (lateral); e, left P5; f, right P5; h, grasping A1 (segments 17, 18, 19 to 21, 22).
|
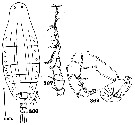 issued from : C.B. Wilson in Bull. U.S. natn Mus., 1950, 100, 14 (4). [Pl.25, Figs.366, 367, 369]. As Labidocera tenuicauda. Male (from 10°07'50''N, 123°57'E): 366, habitus (dorsal); 367, right A1; 369, P5.
|
 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.32, Fig.5, 1]; As Labidocera tenuicauda. After C.B. Wilson, 1950.. 1, distal part of geniculate A1 male.
| | | | | Compl. Ref.: | | | Esterly, 1919 (p.1, 44, light-dark reactions, as trispinosum); Sewell, 1948 (p.393, 411); Sherman, 1963 (p.223: fig.13); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Heinrich, 1964 (p.86: fig.4); Fleminger, 1967 a (tabl.1); Lillelund & Lasker, 1971 (p.655); Landry, 1978 (p.1103, predatory feeding behavior); 1983 (p.614, development times v.s. stages); Brinton & al., 1986 (p.228, fig.11: spatial distribution); Landry & Fagerness, 1988 (p.509, fig.3: clearance rate, Table 3, 5, 6, figs. 4, 7); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Hernandez-Trujillo, 1989 (tab.1); 1989 a (tab.1); 1991 (1993) (tab.I); Huntley & Lopez, 1992 (p.201, Table A1, egg-adult weight, temperature-dependent production); Hernandez-Trujillo, 1994 (tab.1); Landry & al., 1994 (p.55, abundance, grazing); Palomares Garcia & Vera, 1995 (tab.1); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Marcus, 1996 (p.144); Suarez-Morales & Gasca, 1998 a (p.110); Mauchline, 1998 (tab.30, 40, 47, 63, 65); Hernandez-Trujillo, 1999 (p.284, tab.1); Rebstock, 2001 (tab.2); 2002 (p.71, Table 3, 5, 6, Fig.2, climatic variability); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño, Rem.: 67, 70, 71, 73); Hernandez-Trujillo & al., 2010 (p.913, Table 2); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence) | | | | NZ: | 3 | | |
|
Distribution map of Labidocera trispinosa by geographical zones
|
| | | 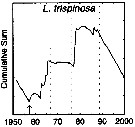 issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 w]. issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 w].
Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California (31°-35°N, 117°-122°W.
Cumulative sums of nonseasonal anomalies from the long-term means of copepod abundance from years 1950 to 2000.
A negative slope indicates a period of below-average anomalies; a positive slope indicates a period of above-average anomalies. Abrupt changes in slope indicate step changes. Step changes are marked with arrow (downward -pointing for decreases).
The October 1966 cruise (prior to the increase in sampling depth), March 1976 cruise (prior to the 1976-77 climatic regime shift), and October 1988 cruise (prior to the hypothesized 1989 climatic regime shift) are marked with vertical lines. |
 Issued from : M.R. Landry in Limnol. Oceanogr., 1978, 23 (6). [p.1106, Fig.1].Feeding of Labidocera female on an Artemia nauplius (N) showing typical orientation of the prey az it is detected (A) and in position to be captured (B). Issued from : M.R. Landry in Limnol. Oceanogr., 1978, 23 (6). [p.1106, Fig.1].Feeding of Labidocera female on an Artemia nauplius (N) showing typical orientation of the prey az it is detected (A) and in position to be captured (B).
Nauplius is entrained in copepod's swimming and is accelerating toward predator. Copepod is also accelerating toward prey.
Photographs were recreated from a feeding sequence filmed at 50 frames per second using hokographic techniques (see Stewart & al., 1973). Time interval is 0.10 s.
Nota: Animals captured by subsurface net tows (about 5 m) from the coastal waters off La Jolla (California), 1 or 2 days before being used in feeding experiments.from nearshore southern California) |
 Issued from : E. Brinton, A. Fleminger & D. Siegel-Causey in CalCOFI Rep., 1986, 27. [p.244, Fig.11]. Issued from : E. Brinton, A. Fleminger & D. Siegel-Causey in CalCOFI Rep., 1986, 27. [p.244, Fig.11].
labidocera copepods of the Gulf of California and Bja California. Overlapping ranges of three sepcies of the ''Labidocera trispinosa group'', an inshore ''green-water'' group whose members do not usually co-occur in samples with species of the ''L. jollae group'' |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.513, Fig.3]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.513, Fig.3].
Clearance rates of Labidocera tripsinosa feeding on naupliar (closed circles) and copepodid stages (open circles).
Data from Landry (1978). |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.514, Table 3]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.514, Table 3].
Comparison of size of body and appendages for seven species of marine copepods. Relationships among predation rates, selection patterns, swimming speed and orientation, and morphology of prey sensory and capture appendages. |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.522-523, Table 5]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.522-523, Table 5].
Comparison of the predatory characteristics of seven planktonic marine copepods.
F max is the mean, maximum clearance rate measured for the predator feeding on its preferred prey. Prey are developmental stages of common species of planktonic copepods.
F = female, M = male, C5 = stage 5. |
| | | | Loc: | | | California, San Diego (La Jolla), Santa Monica Basin, La Jolla, W Baja California (Bahia Magdalena (very abundant, La Paz), G. of California, Bahia Cupica (Colombia), [Indian: Durban (in Brady, 1915)] | | | | N: | 25 | | | | Lg.: | | | (137) M: 1,95; (142) F: 1,6; M: 2,2-1,7; (1259) F: 2,6; {F: 1,60-2,6; M: 1,70-2,20}
(1259) F;
(1259) F: 2,6 (total length), 1,9 (prosome length); A1 = 9,0 mm. (total length). ( in Landry & Fagerness, 1988, Table 3). | | | | Rem.: | The presence of this species in SE of South Africa (Durban) is very doubtful.
Diagnosis for L. trispinosa in the 'darwinii' species group after Prusova & Al-Yamani (2014, p.1165) :
Females:
- 1: Urosome of 3 somites.
- 2: Genital somite asymmetrical, caudal rami of approximately equal length.
- 3: Caudal rami symmetrical, or roughly symmetrical, separate from anal somite.
- 4: Exopod of nearly symmetrical P5 with 2 or 3 terminal points, genital pore ventro-lateral.
- 5: P5 exopod with 2 terminal points, inner one larger.
- 6: 2nd urosomal somite in dorsal view with small lateral swelling on left proximally.
Males:
- 1: Urosome of 5 somites.
- 2: Prosome posterior corners asymmetrical, right corner longer or modified.
- 3: Prosome posterior corners strongly asymmetrical, with 3 prominent spiniform prongs on right, lateral-most the largest.
- 4: Lateral-most prosomal prong crescent-like in lateral view. | | | Last update : 05/11/2021 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 27, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
