|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Temoridae ( Family ) |
|
|
|
Temora ( Genus ) |
|
|
| |
Temora discaudata Giesbrecht, 1889 (F,M) | |
| | | | | | | Syn.: | Temora stylifera : Mori, 1937 (1964) (p.66, figs. juv.F); ? Dakin & Colefax, 1933 (p.207); 1940 (p.92, figs.F,M); Wilson, 1942 a (p.209); 1950 (p.343, fig.F); Fagetti, 1962 (p.26); ? Chiba, 1953 e (p.722, figs.M); 1953 c (p.695, figs.M); ? Tanaka, 1963 (p.14, Rem.F,M); ? Dias & Araujo, 2006 (p.67, figs.F, M); ? Johan & al., 2012 (2013) (p.1, Table 1);
Temora dubia Brady, 1883; 1914 (p.25 Rem., figs.M).
No Temora discaudatus : Grice & Hart, 1962 (p.287, table 3) | | | | Ref.: | | | Giesbrecht, 1892 (p.328, 338, 775, figs.F,M); Giesbrecht & Schmeil, 1898 (p.101, Rem. F,M); A. Scott, 1909 (p.118, Rem.); Pesta, 1912 a (p.47, figs.F,M); Sewell, 1912 (p.353, 365, figs.F, Rem.); 1914 a (p.227); Früchtl, 1924 b (p.53); Mori, 1929 (p.175, figs.F,M); Sewell, 1932 (p.245); Dakin & Colefax, 1933 (p.207); Farran, 1936 a (p.109); Mori, 1937 (1964) (p.65, figs.F,M); Wilson, 1942 a (p.209, figs.F,M); Pesta, 1943 (p.28, figs.F,M); Sewell, 1947 (p.164); C.B. Wilson, 1950 (p.342, figs.F,M); Chiba & al., 1957 (p.308); 1957 a (p.11); Tanaka, 1960 (p.49); Brodsky, 1962 c (p.126, figs.F,M); Kasturirangan, 1963 (p.40, 41, figs.F,M); Tanaka, 1963 (p.13, Rem.F,M); Chen & Zhang, 1965 (p.65, figs.F,M); Vervoort, 1965 (p.102, Rem.); Gonzalez & Bowman, 1965 (p.249, Rem.); Saraswathy, 1966 (1967) (p.79); Koga, 1968 (p.17, fig.: egg); Itoh, 1970 (tab.1, fig.Md); Kos, 1972 (Vol.1, figs F, M, Descr.); Fleminger & Hulsemann, 1973 (p.344, figs.F,M, chart; Fleminger, 1975 (p.397); Marques, 1976 (p.992, figs.F); Goswami & Goswami, 1978 (p.111, figs.); Greenwood, 1978 (p.2, figs.F, Juv.M, Rem.); Dawson & Knatz, 1980 (p.8, figs.F,M); Marques, 1982 (p.760); van der Spoel & Heyman, 1983 (p.105); Koga, 1984 (p.43, Rem.: p.51); Kimmerer & al., 1985 (p.426); Arcos & Fleminger, 1991 (p.1177, figs.F,M, juv.5, Rem.); Chihara & Murano, 1997 (p.916, Pl.180: F,M); Bradford-Grieve & al., 1999 (p.884, 954, figs.F,M); Bradford-Grieve,1999 b (p.158, figs.F,M, figs.182, 193); Al-Yamani & Prusova, 2003 (p.69, figs.F,M); Conway & al., 2003 (p.138, figs.F,M, Rem.); Phukham, 2008 (p.105, figs.F,M) Blanco-Bercial & al., 2011 (p.103, Table 1, Biol. mol, phylogeny); Al-Yamani & al., 2011 (p.51, 52, figs.F,M); Gusmao & al., 2013 a (p.300, Rem., sex ratio); Kos, 2016 (p.43, figs. F, M, Rem.)
| 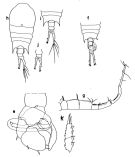 Issued from : Fleminger & Hulsemann in B. Zeitschel, ed., The Biology of the Indian Ocean. 1973. Female: h, habitus (dorsal view); i, Th5 and Urosome dorsal (Mombasa Harbor, Kenya); j, Furca (Manila Bay, Philippines). Male: f, habitus (dorsal); e, P5 (posterior); g, A1 (right); k, exopodite of P2.
|
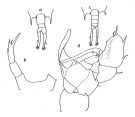 issued from : Bradford-Grieve J.M. in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. Female: A, Urosome (dorsal view); B, P5 Male: C, Urosome (dorsal view); D, P5. Nota female: Similar to T. stylifera but anal segment and caudal rami asymmetrical. Nota male: similar to T. stylifera but segments15 and 16 of geniculate A1 wider, and segments 19-21 more strongly curved. - Terminal spine of exopod segment 3 of left P2 with a row of spines terminally and subterminal outer spine more than 2/3 the length of terminal spine. - P5 with a wider process on left basipod 2 and a much longer terminal hook on the right leg.
|
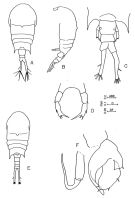 Issued from : Al-Yamani & Prusova in Common Copepods of the Northwestern Arabian Gulf: Identification Guide. Kuwait Institute for Scientific Research. 2003. Female: A, habitus (dorsal view); B, habitus (left lateral view); C, Urosome and P5 (dorsal); D, P5. Male: E, habitus (dorsal); F, P5.
|
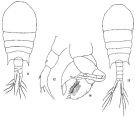 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.20, 11-14]. Female (from E China Sea): 11, habitus (dorsal); 12, left P5 (posterior). Male: 13, habitus (dorsal); 14, P5 (anterior).
|
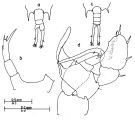 issued from : J.G. Greenwood in Proc. R. Soc. Qd, 1978, 89. [p.3, Fig.1]. Female (from moreton Bay, E Australia): a, posterior prosome and urosome (dorsal); b, right P5. Male: c, posterior prosome and urosome (dorsal); d, P5.
|
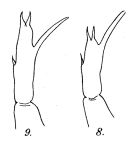 Issued from : R.B.S. Sewell in Rec. Indian Mus., 1912, 7. [Pl.XXII, Figs.8-9]. Female (from Bay of Bengal): 8, P5 (normal); 9, P5 (variety). Nota: In the majority of cases, the inner spine arises close to the end of the terminal segment, but in others it arose from the inner border middle and distal thirds.
|
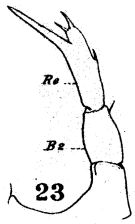 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.17, fig.23]. Female: 23, P5.
|
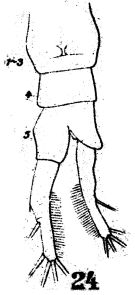 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.38, fig.24]. Female: 24, urosome (ventral).
|
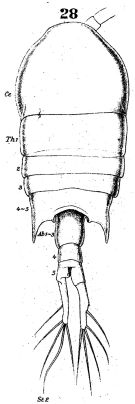 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.38, fig.28]. Female: 28, habitus (dorsal).
|
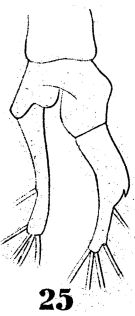 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.38, fig.25]. Female: 25, anal segment and caudal rami (another specimen).
|
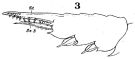 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.17, fig.3]. Male: 3,exopodite 3 of P2 (anterior view).
|
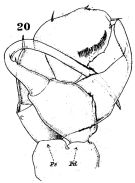 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.17, fig.20]. Male: 20, P5 (anterior view).
|
 issued from : S. Ohtsuka & R. Huys in Hydrobiologia, 2001, 453/454. [p.448, Fig.4, D-E]. Male (from Japan): A, modified elements on geniculate A1 of compound segment XXI-XXIII; E, idem on segment XIX. Scale bars = 0.01 mm.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.32, Figs.9-12]. Female: 10, habitus (dorsal); 12, P5. Nota: The inner marginal spine of the 3rd segment of P5 is much longer than the 2 terminal spines which are about equal length. Male: 9, P5; 11, habitus (dorsal).
|
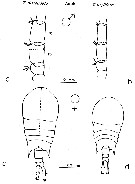 issued from : F. Arcos and A. Fleminger in J. Plankton Res., 1991, 13 (6). p.1183, Fig.5]. Comparison between T. discaudata (from W coast of Mexico) and T. stylifera (from Caribbean Sea off Honduras). a, b, right A1 male (segments 14-16); c, d, habitus (dorsal).
|
 issued from T. Mori in Zool. Mag. Tokyo, 1929, 41 (486-487). [Pl. VI, Figs.8-13]. Female (from Chosen Strait, Korea-Japan): 8, P5; 11, P4; 12, last thoracic segment and urosome. Male: 9, P5 (anterior); 10, last thoracic segment and urosome (dorsal); 13, right A1.
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.189, Fig.63]. Female (from W Malay Peninsula): a, habitus (dorsal); b, urosome (dorsal); c, P5. Male: d, habitus (dorsal); e, P5. Body length after the drawings: F = 1.800 mm; M = 1.567 mm.
|
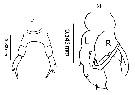 issued from : M.L.D.G. Lacuna, D.C. Sagrado, N.B. España, D.D. Simyunn & R.O. Mejorada in ABAH Bioflux, 2013, 5 (1). [p.104, Fig.6 e; p.105, Fig.7 e]. Female (from Mindanao bays); F: P5. Male: M, P5 (L = left leg; R = right leg).
|
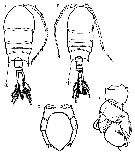 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.918, Pl. 180, fig.281 a-b, e-f]. Female : a, habitus (dorsal); b, P5. Male: e, habitus (dorsal), f, P5. Note characteristics numbered 1 into circles, showing the corner of the last thoracic segment.
|
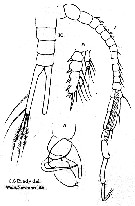 Issued from : G.S. Brady in Ann. Durban Mus., 1914, Vol. III. [Pl. VI, figs. 7-10]. As Temora dubia. Male (from Durban Bay, Natal): 7, Right A1; 8, P3; 9, P5; 10, urosome. Remarks: Brady (1914, p.25) does not admit the synonymy after Giesbrecht (1892).
|
 Issued from : M.S. Kos inZoological Inst. RAS; 2016, 179. [p.44, figs.F, M). Temora discaudata Female and male from 39°34' N, 147°47' E.
|
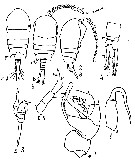 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Brodsky, 1962. Female: 1-2, habitus (doesal and lateral, respectively); 3, corner of the last thoracic segment and abdomen (lateral); 4, abdomen and caudal rami (dorsal); 5, P5. Male: 6, habitus (dorsal); 7, left P5; 8, right P5.
| | | | | Compl. Ref.: | | | Thompson & Scott, 1903 (p.234, 248); Cleve, 1904 a (p.198); Carl, 1907 (p.16); Gurney, 1927 (p.151, Rem.); Sewell, 1948 (p.322, 391); Fox, 1929 (p.843, Rem.: p.851: as T. discordata lapsus calami); Krishnaswamy, 1953 (p.125); Kott, 1957 (p.5); Yamazi, 1958 (p.150, Rem.); Ganapati & Shanthakumari, 1962 (p.8, 16); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); De Decker, 1964 (p.16, 25, 30); De Decker & Mombeck, 1964 (p.14); Anraku & Azeta, 1965 (p.13, Table 2, fish predator); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 9); Fleminger, 1967 a (tabl.1); Grice & Hulsemann, 1967 (p.17); De Decker, 1968 (p.45); Delalo, 1968 (p.138); Brodsky, 1972 (p.256); Heinrich, 1973 (p.95); Björnberg, 1973 (p.337, 389); Patel, 1975 (p.659); Tranter, 1977 (p.598); Carter, 1977 (1978) (p.36); Frontier, 1977 a (p.15); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Stephen & Iyer, 1979 (p.228, tab.1, 3, 4, figs.3, 4); Chen Q-c, 1980 (p.795); Sreekmaran Nair & al., 1981 (p.493, Fig.2); Madhupratap & al., 1981 (p.266, fig.2i: abundance vs. geographic transect); Arashkevich & al., 1982 (p.477, Table 2, diet); Dessier, 1983 (p.89, Tableau 1, 2, Rem., %); Gaudy & Boucker, 1983 (p.37, Table 1, 2, 3, 5, fig.1, 2, Rem.: metabolism); De Decker, 1984 (p.316, 364: chart); Stephen, 1984 (p.161, Distribution vs thermocline & geographic); Guangshan & Honglin, 1984 (p.118, tab.); Binet, 1985 (p.85, tab.3); Sazhina, 1985 a (p.491, tab.1); Petipa & Borichenko, 1985 (tab.2); Sarkar & al., 1986 (p.178); Brinton & al., 1986 (p.228, Table 1, Rem.); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); Renon, 1987 (tab.2); Jimenez-Perez & Lara-Lara, 1988; Wiebe & al., 1988 (tab.7); Dessier, 1988 (tabl.1); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Hirakawa & al., 1990 (tab.3); Othman & al., 1990 (p.561, 563, Table 1); Madhupratap & Haridas, 1990 (p.305, fig.4: vertical distribution night/day, fig.7: cluster); Yoo, 1991 (tab.1); Hattori, 1991 (tab.1, Appendix); Hirakawa, 1991 (p.376: fig.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Kim & al., 1993 (p.270); Tiwari & Nair, 1993 (p.67); Verheye & al., 1994 (p.155); Palomares Garcia & Vera, 1995 (tab.1); Shih & Young, 1995 (p.74); Kotani & al., 1996 (tab.2); Padmavati & Goswami, 1996 a (p.85, fig.3, Table 4, vertical distribution); Sharaf & Al-Ghais, 1997 (tab.1); Park & Choi, 1997 (Appendix); Padmavati & al., 1998 (p.349); Noda & al., 1998 (p.55, Table 3, occurrence); Achuthankutty & al., 1998 (p.1, Table 2, fig.6, seasonal abundance vs monsoon); Hwang & al., 1998 (tab.II); Hsieh & Chiu, 1998 (tab.2); Wong & al, 1998 (tab.2); Noda & al., 1998 (p.55, Table 3, occurrence); Mauchline, 1998 (tab.8); Smith S. & al., 1998 (p.2369, Table 6, moonsoon effects); Suarez-Morales & Gasca, 1998 a (p.111); Hernandez-Trujillo, 1999 (p.284, tab.1); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); El-Serehy, 1999 (p.172, Table 1 as Temara, occurrence); Dolganova & al., 1999 (p.13, tab.1); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Escribano & Hidalgo, 2000 (p.283, tab.2); Suarez-Morales & al., 2000 (p.751, tab.1); Hidalgo & Escribano, 2001 (p.159, tab.2); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters); Lo & al., 2001 (1139, tab.I); Ali-Khan & Ali-Khan, 2002 (p.311); Hernandez-Trujillo & Suarez-Morales, 2002 (p.748, tab.1); Rebstock, 2002 (p.71, Table 3, Fig.5, climatic variability); Osore & al., 2003 (p.69); Hwang & al., 2003 (p.193, tab.2); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.326, tab.1); Hsieh & al., 2004 (p.397, tab.1); Rezai & al., 2004 (p.490, tab.2, p.495, tab.8); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Lo & al.*, 2004 (p.218, tab.1, fig.6); Lo & al., 2004 (p.468, tab.2); Chang & Fang, 2004 (p.456, tab.1); Lan & al., 2004 (p.332, tab.1, tab.2); Gallienne & al., 2004 (p.5, tab.3); Satapoomin & al., 2004 (p.109, tab.6); Lo & al., 2004 (p.89, tab.1); Ali-Khan & Ali-khan, 2004 (p.31, Rem.); Kazmi, 2004 (p.229); Shimode & al., 2005 (p.113 + poster); Alvarez-Silva & al., 2005 (p.39); Smith & Madhupratap, 2005 (p.214, tab.5); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño); Zuo & al., 2006 (p.159, tab.1, 3, abundance, fig.8: stations group); Lavaniegos & Jiménez-Pérez, 2006 (p.157, tab.2, 3, Rem.); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Hwang & al., 2006 (p.943, tabl. I); Hwang & al., 2007 (p.25); Dur & al., 2007 (p.197, Table IV); Jitlang & al., 2008 (p.65, Table 1); Lan Y.-C. & al., 2008 (p.61, Table 1, % vs stations); Rakhesh & al., 2008 (p.154, abundance vs stations); McKinnon & al., 2008 (p.844: Tab.1); Humphrey, 2008 (p.84: Appendix A); Morales-Ramirez & Suarez-Morales, 2008 (p.522); Selifonova & al., 2008 (p.305, Tabl. 2); Fernandes, 2008 (p.465, Tabl.2); Lopez Ibarra, 2008 (p.1, Table 1, figs.11, 16: abundance, Table 3: N, C isotopes, Table 4: index trophic); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Ohtsuka & al., 2008 (p.115, Table 5); Tseng & al., 2008 (p.402, Table 2); Tseng L.-C. & al., 2008 (p.153, fig.5, Table 2, occurrence vs geographic distribution, indicator species); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons); C.-Y. Lee & al., 2009 (p.151, Tab.2); Tseng & al., 2009 (p.327, fig.5, feeding); Zhang W & al., 2009 (p.261); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Zhang G.-T. & al., 2010 (p.492, Table 2); Cornils & al., 2010 (p.2076, Table 3); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Hidalgo & al., 2010 (p.2089, Table 2); Fazeli & al., 2010 (p.153, Table 1); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Xu & Gao, 2011 (p.514, figs.3, 4, Table 2: optimal salinity); Shanthi & Ramanibai, 2011 (p.132, Table 1); Tutasi & al., 2011 (p.791, Table 1, 3, fig.9, abundance distribution vs La Niña event, Rem. p.798, fig.11); Hsiao S.H. & al., 2011 (p.475, Appendix I); Hsiao & al., 2011 (p.232, Table 1: abundance, %, Table 2: trace metal concentration, vs transect); Hsiao & al., 2011 (p.317, Table 2, fig.6, indicator of seasonal change); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Tseng & al., 2011 (p.301, adaptability vs T. turbinata); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Johan & al., 2012 (2013) (p.1, Table 1); Jose & al., 2012 (p.20, fig.3 a,b,c: % vs monsoon); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Tseng & al., 2012 (p.621, Table 3: abundance, indicator species); Naz & al., 2012 (p.61, Table 4, relative abundance); Lavaniegos & al., 2012 (p. 11, Table 1, seasonal abundance, Appendix); Gubanova & al., 2013 (in press, p.4, Table 2); Palomares-Garcia & al., 2013 (p.1009, Table I, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Tseng & al., 2013 (p.507, seasonal abundance); Tseng & al., 2013 a (p.1, Table 3, abundance); Jagadeesan & al., 2013 (p.27, Table 3, 4, 6, fig.11, seasonal abundance); Rakhesh & al., 2013 (p.7, Table 1, abundance vs stations); Varadharajan & Soundarapandian, 2013 (p.2: occurrence vs stations); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Hirai & al., 2013 (p.1, Table I, molecular marker); Mendoza Portillo, 2013 (p.37: Fig.7, seasonal dominance); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Lopez-Ibarra & al., 2014 (p.453, fig.6, biogeographical affinity); Chiba S. & al., 2015 (p.968, Table 1: length vs climate); Nakajima & al., 2015 (p.19, Table 3: abundance); Zakaria & al., 2016 (p.1, Table 1, Rem.); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); Palomares-Garcia & al., 2018 (p.178, fig.3: relative frequency, Table 1) | | | | NZ: | 12 + 1 doubtful | | |
|
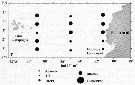
Distribution map of Temora discaudata by geographical zones
|
| | | | | | | | | | | |  Chart of 1996 Chart of 1996 | |
 Distribution of Temora discaudata (hatching) and T. stylifera (cross-hatching); modified from Fleminger and Hulsemann; issued from : F. Arcos and A. Fleminger in J. Plankton Res., 1991, 13 (6). [Fig.7, p.1184] Distribution of Temora discaudata (hatching) and T. stylifera (cross-hatching); modified from Fleminger and Hulsemann; issued from : F. Arcos and A. Fleminger in J. Plankton Res., 1991, 13 (6). [Fig.7, p.1184] |
 issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.8, Fig.6]. issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.8, Fig.6].
Salinity ranges for T. discaudata in coastal and estuarine waters of Goa (India).
Shaded area indicates the range of higher abundance. |
 Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.799, Fig.9 c] Issued from : P. Tutasi, S. Palma & M. Caceres in Scienc. Mar., 2011, 75 (4). [p.799, Fig.9 c]
Geographic distribution of Temora discaudata in September and October 2001, associated with the weak La Niña event of 2001. |
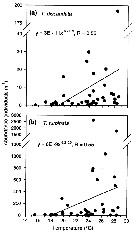 Issued from : L.-C Tseng, R. Kumar, Q.-C. Chen & J.-S. Hwang in Hydrobiologia, 2011, 666. [p.313, Fig.9]. Issued from : L.-C Tseng, R. Kumar, Q.-C. Chen & J.-S. Hwang in Hydrobiologia, 2011, 666. [p.313, Fig.9].
Densities (ind./ m3) of T. discaudata (a) and T. turbinata (b) as a function of seawater temperature (°C) in the southern margin of the East China Sea in the vicinity of Nuclear Power Plant-I and II of northern Taiwan. |
 Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6]. Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6].
Variations in the most abundant copepod species (mean ± SE) along the transect in the boundary waters between the northern part Taiwan Strait and the East China Sea i March (black bar) and October (grey bar) 2005 (Mann-Whitney U test, sig. *P <0.05.
See drawings of Hydrological conditions and superficial marine currents in Calanus sinicus. |
 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 34- Temora discaudata.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : J.R. Palomares-Garcia, S. Hernandez-Trujillo, G.M. Esqueda-Escarcega & A. pérez-Morales, 2018 (p.L178, Fug.3]. Relationships frequency vs abundance of pelagic copepods in the La Paz Bay and the 22 main species (particularly the 3 dominant) over 96 species in the zone (about 21% of the total from the tropical Pacific species.); 59 species (13%) in the La Paz Bay show an annual cycle. Issued from : J.R. Palomares-Garcia, S. Hernandez-Trujillo, G.M. Esqueda-Escarcega & A. pérez-Morales, 2018 (p.L178, Fug.3]. Relationships frequency vs abundance of pelagic copepods in the La Paz Bay and the 22 main species (particularly the 3 dominant) over 96 species in the zone (about 21% of the total from the tropical Pacific species.); 59 species (13%) in the La Paz Bay show an annual cycle. |
 Issued from : Ya-Qu. Chen inCalCOFI Rep., 1986, Vol. XXVII. [p.220]. Issued from : Ya-Qu. Chen inCalCOFI Rep., 1986, Vol. XXVII. [p.220].
For the author, during the Krill Expedition from 23°N to 3°S in the Pacific East tropical (May-June 1974), this species shows a mean percentage of the total copepods: 0.9 % (day: 0.7, night: 1.0), but with a bongo net (0.333 µm and 220 µm). This species was found commonly in coastal waters of tropical to warm-temperate areas.
It occurred mainly above 400 m over the entire area of investigation, except for the oxygen-deficient region. It main concentrations were found between 50 to 100 m.
It maximum concentration (11,000 individuals/m 3) was in a day sample from 0 to 50 m at station 10, although numbers were usually higher in night samples than in day samples. (see in Subeucalanus subtenuis: Chart Fig.1, hydographic conditions Figs.12, 13, after the same author). |
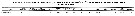 Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.37, Table 3]. Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.37, Table 3].
Summary of abundance and steady isotopics 15N and 13C in Subeucalanus subcrassus.
O = omnivore; T = tropical biogeographical affinity; zones (a-f) = Cf. fig.1 in the same author.
Compare with Subeucalanus subcrassus and other species (Tabla 3). |
| | | | Loc: | | | South Africa (E & W), Agulhas Current, ? Brazil ( Vitoria-Cabo de Sao Tomé), no off Bermuda (in Grice & Hart, 1962, p.294, 295), Chesapeake Bay (in Wilson, 1932, p.32), E Mediterranean Sea (in Thompson & Scott, 1903), W Egyptian coast, W Black Sea, Canal of Suez, Red Sea, Gulf of Oman, G. of Aden, Arabian coast, Karachi coast, Arabian Sea, UAE coast, Kuwait, Laccadive and Maldive archipelagos,Agatii Is. (lagoon), Sri Lanka Natal, Madagascar (Nosy Bé), Rodrigues Is. - Seychelles, Mascarene Basin, India (Saurashtra coast, Mangalore coast, Goa - Gujarat, off Cochin, Coom & Adyar, G. of Mannar, Manamelkudi, Palk Bay, Madras, Lawson's Bay, Kakinada Bay, Hooghly estuary, Godavari region), E India, Bay of Bengal, Nicobar Is., Batten Island, W Malay Peninsula, off Phuket, G. of Thailand, Australia (Shark Bay, North West Cape), Straits of Malacca, Thailand, Indonesia-Malaysia, Sarawak: Bintulu coast, Ambon Bay, Tioman Is., SW Celebes, SE Philippines (Mindanao Bays) Viet-Nam ( Cauda Bay), Gulf of Tonkin, Hong Kong, China Seas (Yellow Sea, East China Sea, South China Sea, Changjiang River estuary, Xiamen Harbour), Taiwan Strait, Taiwan (S, E, SW, W, Tapong Bay, Kaohsiung Harbor, NW, N, NE, Mienhua Canyon), S Korea, Japan Sea, Nagazaki, Japan, Kuchinoerabu Is., Sagami Bay, Tanabe Bay, Tokyo Bay, off Sanriku, E Tori Is., W Pacif. (equatorial), Australia (G. of Carpentaria, SE, Great Barrier, Moreton Bay), New Caledonia, New Zealand (Cook Strait), E Pacif. (equatorial), Guaymas Basin, California, W BaJa California, Bahia Magdalena (abundant), Gulf of California (.., Bahia de los Angeles, La Paz), Zihuatanejo Bay, W Mexico, G. of Tehuantepec, Clipperton Is., W Costa Rica, Bahia Cupica (Colombia), Galapagos-Ecuador, off Peruvian coast, off N Chile, Chile (N-S, Mejillones Peninsula)
Locality in Kos (2016, p.42): 39°34' N, 147°47' E. | | | | N: | 220 ? | | | | Lg.: | | | (26) F: 2,05-1,62; M: 1,88-1,68; (47) F: 2-1,7; M: 1,9-1,7; (66) F: 1,8-1,73; (91) F: 2-1,7; M: 1,9-1,7; (150) F: 1,7; M: 1,7; (290) F: 1,8-1,95; M: 1,7-1,8; (332) F: 1,85-1,66; M: 1,84-1,67; (333) F: 1,88-1,69; M: 1,97-1,73; (334) F: 1,9; M: 1,8; (530) F: 2; M: 1,8; (578) M: 1,07-1,03; (786) F: 2,05-1,73; M: 1,95-1,78; (795) F: 1,7; M: 1,5; (864) F: 1,11-1,63; (866) F: 1,58-1,95; M: 1,68-1,93; (937) F: 1,79-1,85; M: 1,79-1,83; (991) F: 1,69-2; M: 1,7-1,97; (1047) F: 1,6; (1085) F: 1,6-2,0; M: 1,7-1,85; (1125) F: 2,0; (1140) F: 1,8; M: 1,77; (1230) F: 1,7-2,0; M: 1,7-1,9; {F: 1,11-2,05; M: 1,50-1,97}
Chiba S. & al., 2015 (p.971, Table 1: Total length female (June-July) = 1.8 mm [optimal SST (°C) = 15.2].
| | | | Rem.: | nearshore-off shore. Shelf.
See Remarks from Gurney (1927, p.151) concerning the two very closely allied species T. discaudata and T. stylifera in the expedition's collections in the Suez Canal, from Port Said to Suez Bay); for the author T. discaudata is a purely Indo-Pacific species and T. stylifera north Atlantic-Mediterranean species.
For Gonzales & Bowman (1965) Wilson's (1942, 1950) records of T. stylifera from the Pacific were misidentifications of T. discaudata, as were those of Mori (1937) and Chiba (1953 a, 1953 b). They suggest the two species are allopatric, T. discaudata being Indo-Pacific and T. stylifera being Atlantic. According to Greenwood (1978, p.4) however, whilst it may be true that many of the remaining records of T. stylifera in faunistic lists from the north-west Pacific could be misidentified, this cannot be verified for they are not illustratd. T. stylifera is well recorded from throughout the Indian Ocean, though less abundantly than from the Atlantic and into the Indo-Malaysian region. It would not be surprising if it appeared in the western Pacific, together with the resident T. discaudata.
For Itoh (1970 a, fig.2, from co-ordonates) the Itoh's index value of the mandibular gnathobase = 480, and for Gaudy & Boucher (1983): 500.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a coarse-filter feeder herbivorous.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 09/12/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed April 16, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
