 List of the studied authors
List of the studied authors
Cleve, 1904 a (p.198); Carl, 1907 (p.16); Sewell, 1912 (p.366); Carazzi & Grandori, 1912 (p.12, 38); Chatton, 1920 (p.17); Rose, 1924 d (p.479); Gurney, 1927 (p.151,
Rem.); Wilson, 1942 a (p.209); Massuti Alzamora, 1942 (p.91); Oliveira, 1945 (p.191); Sewell, 1948 (p.391, 406); C.B. Wilson, 1950 (? part., p.343); Krishnaswamy, 1953 (p.125); Deevey, 1960 (p.5, Table II, annual abundance); Cervigon, 1962 (p.181, tables: abundance distribution); Grice & Hart, 1962 (p.287, table 3); V.N. Greze, 1963 a (tabl.2); Shmeleva, 1963 (p.141); Gaudy, 1963 (p.24, Rem.); Björnberg, 1963 (p.46, Rem.); Mazza, 1964 (p.293, weight); De Decker & Mombeck, 1964 (p.14); Roehr & Moore, 1965 (p.565, vertical distribution); Shmeleva, 1965 b (p.1350, lengths-volume-weight relation); Neto & Paiva, 1966 (p.26, Table III, annual cycle); Vives, 1966 (p.81); Pavlova, 1966 (p.43); Mazza, 1966 (p.71); 1967 (p.356, 377, 399, fig.76); Ehrhardt, 1967 (p.738, geographic distribution, Rem.); Séguin, 1968 (p.488); Evans, 1968 (p.15); Delalo, 1968 (p.138); Berdugo & Kimor, 1968 (p.447); Champalbert, 1969 a (p.636); Moraitou-Apostolopoulou, 1969 (p.2, fig.1); El-Maghraby & Dowidar, 1970 (p.81); Dowidar & El-Maghraby, 1970 (p.269); M. Bernard, 1970 (p.75); Park, 1970 (p.476); Shih & al., 1971 (p.49); Salah, 1971 (p.319); Deevey, 1971 (p.225); Gamulin, 1971 (p.381, tab.2); Carli, 1971 (p.372, tab.1); Gaudy, 1971 (p.65, Tabl. 1, fig.1, egg production); 1972 (p.175, 200, figs.13-18, annual cycle); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs. day/night, Table II: %), Table IV; Brodsky, 1972 (p.256); Binet & al., 1972 (p.71); Della Croce & al., 1972 (p.1, Rem.); Riera, 1972 (p.67, morphometry); Razouls S., 1972 (p.95, metabolism); 1972 a (p.145, respiration); 1972 b (p.2, respiration); Champalbert & Gaudy, 1972 (p.159, respiration vs temperature); Valentin, 1972 (p.377: egg diameter = 80 µm); Ibanez & Seguin, 1972 (p.81, annual cycle, multivarite analysis); Nival & al., 1972 (p.63, respiration); Apostolopoulou, 1972 (p.328, 352, fig.4); Boucher & Thiriot, 1972 (p.47, Tableau 4); Roe, 1972 (p.277, tabl.1, tabl.2); C. Razouls & Guinness, 1973 (p.413, size variations & temperature); Champalbert & al., 1973 (p.529, CHN composition); Gaudy, 1973 (p.267, Table I, II, fig.2, 5, respiration); P. Nival & S. Nival, 1973 (p.135, mouth parts, grazing); Desgouille, 1973 (p.1, 131, Rem.: p.138); Guglielmo, 1973 (p.399); C. Razouls, 1973 a (p.361, annual abundance); 1974 (p.51, life history); S. Razouls, 1974 (147, oxygen rate); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Nival & al., 1974 (p.231, respiration & excretion); Gaudy, 1975 (p.109, Table I, respiration); Le Ruyet-Person & al., 1975 (p.283, comparative biology, metabolism activity); S. Razouls, 1975 (p.297, egg production); Vives & al., 1975 (p.43, tab.II, III, IV); Boucher & al., 1975 (p.85, nutrition/enzyme); Fernandez, 1975 (p.1, fig.1, 2, 3, 4, metabolism/lux), Boucher & al., 1976 (p.61, elementary composition); Lakkis & Abboud, 1976 (p.81); Razouls & Razouls, 1976 (p.281, annual variations); Boucher & al., 1976 (p.61, 62); C. Razouls & S. Razouls, 1976 (p.281, caloricity, growth); S. Razouls, 1977 (p.265, caloricity); Weikert, 1977 (p.351, Rem.: p.353, exuviae); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Scaramuzza, 1978 (p.17); Paffenhöfer & Knowles, 1978 a (p.143, feeding); Fernandez, 1978 (p.97, metabolism/food, Rem.: Table 19); S. Razouls & Apostolopoulou, 1978 (p.13, metabolism); Binet, 1979 (p.400); Dessier, 1979 (p.100, 201, 206); Paffenhöfer & Knowles, 1979 (p.35, fecal pellets); Stephen & Iyer, 1979 (p.228, tab.1, 3, 4, figs.3, 4); Vaissière & Séguin, 1980 (p.23, tab.2); Andronov & Maigret, 1980 (p.71, Table 1, 2, 3); Paffenhöfer & Knowles, 1980 (p.355, feeding); Yassen, 1981 (p.125, rearing, mortality rate); Sreekmaran Nair & al., 1981 (p.493, Fig.2); Madhupratap & al., 1981 (p.266, fig.2h: abundance vs. geographic transect); Kovalev & Shmeleva, 1982 (p.84); Vives, 1982 (p.293); Castel & Courties, 1982 (p.417, Table II, spatial distribution); Boucher, 1982 a (p.199, fig.2); Yassen, 1982 (p.125, culture, mortality rate); Riera, 1983 (p.363, biometry); Abou Debs & Nival, 1983 (p.125, egg production vs. diet, temperature); Rivière, 1983 (p.19, enzymes); Greze & al., 1983 (p.17, Rem.: p.24); Turner & Dagg, 1983 (p.16, 22); Moore E.A. & Sander, 1983 (p.113, fig.1: variation % sex ratio); De Decker, 1984 (p.316, 364: chart); Tremblay & Anderson, 1984 (p.7); Turner, 1984 a (p.73, Table 1, fig.4-7: pellet contents); Scotto di Carlo & al., 1984 (1041); Cummings, 1984 (p.163, Table 2); ? Guangshan & Honglin, 1984 (p.118, tab.); Miller & al., 1984 (p.1274, Rem.: p.1278, molting rates); Abou Debs, 1984 (p.213, egg & fecal pellets porduction, respiration rate, C & N budget vs. diet); Gruzov, 1985 (p.633, age dependant feeding); Regner, 1985 (p.11, Rem.: p.34); Brenning, 1985 (p.5, spatial distribution/TS); 1985 a (p.23, Table 2); Jansa, 1985 (p.108, Tabl.I, II, III, IV, V); Petipa & Borichenko, 1985 (tab.2); Gaudy, 1985 (p.279, Tab.3); Garcia-Rodriguez, 1985 a (p.41, 42); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea); Valentin & al., 1986 (p.117, Table V); Riera & Alcaraz, 1986 (p.127, biometry); Ambler, 1986 a (p.957, fig.7, selectivity); Comaschi Scaramuzza, 1987 (tab.1); Boucher & al., 1987 (p.133, spatial distribution/physical structure); Lozano Soldevilla & al., 1988 (p.59); Daan & al., 1988 (p.45, carnivorous behaviour); Ianora & al., 1989 (p.187, reproduction); Kouwenberg & Razouls, 1990 (p.23, climatic change); Suarez & al., 1990 (tab.2); Yoo, 1991 (tab.1); Lindo, 1991 (tab.3); Suarez & Gasca, 1991 (tab.2); Samba Diouf, 1991 (p.104); Carlotti & Nival, 1991 (p.801, variability of development); Suarez, 1992 (App.1); Huntley & Lopez, 1992 (p.201, Table 1, eggs, temperature-dependent production); C. Razouls & S. Razouls, 1992 (p.259, life history); Ayukai & Hattori, 1992 (p.163, Table 5, fecal pellet production rate); Kouwenberg, 1993 (p.215, fig.3, seasonal abundance); 1993 a (p.281, fig.3, 4, sex ratio); Ianora & Poulet, 1993 (p.1615, egg production & viability/food); Seguin & al., 1993 (p.23); Lopes, 1994 (tab.1); Kouwenberg, 1994 (tab.1); Hajderi, 1995 (p.542); Webber & Roff, 1995 (tab.1); Shih & Young, 1995 (p.74); Roff & al., 1995 (p.165, Table 5: naupllii vs. bacteriophagy/picoplankton); Kotani & al., 1996 (tab.2); Webber & al., 1996 (tab.1); Jamet & Ferec-Corbel, 1996 (p.17, tab.1); Paffenhöfer & al., 1996 (p.1699, motion behavior); Ban S, Burns C. & al., 1997 (p.287, fig.1, Table 1, 2, feeding, reproduction); Suarez-Morales & Gasca, 1997 (p.1525); Park & Choi, 1997 (Appendix); Hure Krsinic, 1998 (p.59, 101); Gilabert & Moreno, 1998 (tab.1, 2); Lopes & al., 1998 (p.195); Alvarez-Cadena & al., 1998 (tab.1,3,4); Kouwenberg, 1998 (p.203, seasonal variation, fig.3: global change); Vigoni & al., 1998 (tab.2); Hopcroft & al., 1998 (tab.2); Noda & al., 1998 (p.55, Table 3, occurrence); Hsieh & Chiu, 1998 (tab.2); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p.111); Mauchline, 1998 (tab.8, 16, 21, 25, 26, 27, 30, 38, 45, 47, 48, 58, 61, 62, 63); Lopes & al., 1999 (p.215, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Miralto & al., 1999 (p.173, Rem.: p.175, egg production vs diatoms effect of inhibition); Lapernat, 1999 (p.29, 55); Siokou-Frangou, 1999 (p.476); Lapernat, 2000 (tabl.3, 4); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Suarez-Morales & Gasca, 2000 (1247, tab.1); Seridji & Hafferssas, 2000 (tab.1); Plounevez & Champalbert, 2000 (p.175, Table III, IV, V, abundance vs fish, Rem.: p.185); Pakhomov & al., 2000 (p.1663, Table 2, transect Cape Town-SANAE antarctic base); Lopez-Salgado & al., 2000 (tab.1); d'Elbée, 2001 (tabl. 1); Lapernat & Razouls, 2001 (p.123, tab.1); Pardal & Azeiteiro, 2001 (p.25); El-Serehy & al., 2001 (p.116, Table 1: abundance vs transect in Suez Canal); Calbet & al., 2001 (p.319, Fig.7); Halsband-Lenk & al., 2001 (p.597, seasonal cycle, egg production); Zerouali & Melhaoui, 2002 (p.91, Tableau I); Beaugrand & al., 2002 (p.179, figs.5, 6); Bressan & Moro, 2002 (tab.2); Dunbar & Webber, 2003 (tab.1); Vukanic, 2003 (139, tab.1); Bode & al., 2003 (p.85, Table 1, abundance); Fernandez de Puelles & al., 2003 (p.123, fig.5); Ceballos & Ianora, 2003 (p.189, egg production); Hsiao & al., 2004 (p.326, tab.1); Hsieh & al., 2004 (p.397, tab.1, p.398: Rem., p.399, tab.2); Rezai & al., 2004 (p.486, p.490: tab.2, p.495: tab.8, abundance); Chang & Fang, 2004 (p.456, tab.1); Daly Yahia & al., 2004 (p.366, fig.4, tab.1); Lan & al., 2004 (p.332, tab.1); Di Capua & Mazzocchi, 2004 (p.632); Halsband-Lenk & al., 2004 (p.709, figs.6,7); CPR, 2004 (p.63, fig.189); Vukanic & Vukanic, 2004 (p.9, tab. 3); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Dias & Bonecker, 2005 (p.100 + poster); Di Capua & al., 2005 (p.72 + poster); Lakkis & al., 2005 (p.152); Uriarte & Villate, 2005 (p.863, tab.I); Frangoulis & al., 2005 (p.254, Table I: C/N/P fecal pellet composition); Molinero & al., 2005 (p.640, climate forcing); Lindley & Daykin, 2005 (p.869, occurrence between 1959 to 2000); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: stations group); Isari & al., 2006 (p.241, tab.II); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Dias & Araujo, 2006 (p.67, Rem., chart); Zervoudaki & al., 2006 (p.149, Table I); De Olazabal & al., 2006 (p.966); Koppelmann & Weikert, 2007 (p.266: tab.3); Fernandez de Puelles & al., 2007 (p.338, fig.7); Albaina & Irigoien, 2007 (p.435: Tab.1); Valdés & al., 2007 (p.104: tab.1, figs.7, 8); Khelifi-Touhami & al., 2007 (p.327, Table 1); Busatto, 2007 (p.26, Tab.2); Dur & al., 2007 (p.197, Table IV); Aldamman, 2008 (p.1-154); Kiørboe, 2008 (p.155, Table 2, swimming speed); Cabal & al., 2008 (289, Table 1); Rossi, 2008 (p.90: Tableau XII); Gugliandolo & al., 2008 (p.580, bacteria assiociated); Jerling, 2008 (p.55, Tabl.1); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Goetze & Kiørboe, 2008 (p.185, mating recognition); Fernandes, 2008 (p.465, Tabl.2); Muelbert & al., 2008 (p.1662, Table 3); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Zhang W & al., 2009 (p.261); Brugnano & al., 2010 (p.312, Table 2, 3); Hafferssas & Seridji, 2010 (p.353, Table 2); Eloire & al., 2010 (p.657, Table II, temporal variability); Lidvanov & al., 2010 (p.356, Table 3); Drira & al., 2010 (p.145, Tanl.2); Cornils & al., 2010 (p.2076, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Dias & al., 2010 (p.230, Table 1); Xu & Gao, 2011 (p.514, figs.3, 4, Table 2: optimal salinity); Mazzocchi & Di Capua, 2010 (p.428); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Yen & Lasley, 2010 (p.177, Rem.: p.183, mating): Hafferssas & al., 2010 (p.1281, Table III, abundance vs spatial distribution); Fazeli & al., 2010 (p.153, Table 1); W.-B. Chang & al., 2010 (p.735, Table 2, 4, abundance); Ianora & Miralto, 2010 (p.493, Table I: reproductive responses vs diatom diets); Nowaczyk & al., 2011 (p.2159, Table 2); Shanthi & Ramanibai, 2011 (p.132, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Magris & al., 2011 (p.260, abundance, interannual variability); Costa R.G. da & al., 2011 (p.364, Table 1, seasonal occurrence); Moscatello & al., 2011 (p.80, Table 4); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Barreiro & al., 2011 (p.13, egg production, hatching success, ingestion rate vs food selection); Mazzocchi & al., 2011 (p.1163, Table I, II, fig.6, long-term time-series 1984-2006); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Ianora & al., 2011 (p.265, diet vs egg production, egg viability, fecal pellets production, survival); ); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Delpy & al., 2012 (p.1921, Table 2); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Almeida LR. & al., 2012 (p.13, Table 1, abundance); Salah S. & al., 2012 (p.155, Tableau 1); Zizah & al., 2012 (p.79, Tableau I, Rem.: p.89); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Vidjak & al., 2012 (p.243, Rem.: p.254); Aubry & al., 2012 (p.125, table 1, 3, fig. 6, 8 a, b, interannual variation); Johan & al., 2012 (2013) (p.1, Table 1); Turner & al., 2012 (p.63, toxic dinoflagellate effects); Drillet & al., 2012 (p.155, Table 1, culture); Bode & al., 2012 (p.108, spatial distribution vs time-series, % biomass); Mazzocchi & al., 2012 (p.135, annual abundance 1984-2006); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Neffati & al., 2012 (p.80, egg production & nauplii survival vs environmental factors); Buttino & al., 2012 (p.247, culture); Gusmao & al., 2013 (p.279, Table 4, sex ratio); Nikolioudakis & al., 2012 (p.173, Table 2: Ivelv's selectivity index); Gubanova & al., 2013 (in press, p.4, Table 2); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Teuber & al., 2013 (p.1, Table 1, 2, abundance vs oxygen minimum zone, respiration rates); Jagadeesan & al., 2013 (p.27, Table 3, fig.11, seasonal abundance); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Sobrinho-Gonçalves & al., 2013 (p.713, Table 2, fig.8, seasonal abundance vs environmental conditions); Terbiyik Kurt & Polat, 2013 (p.1163, Table 2, fig.9, seasonal distribution); Lidvanov & al., 2013 (p.290, Table 2, % composition); El-Serehy & al., 2013 (p.2099, Rem.: p.2101, as
Tempora stylifera); Varadharajan & Soundarapandian, 2013 (p.2: occurrence vs stations); Fernandez de Puelles & al., 2014 (p.82, Table 3, seasonal abundance); Bonecker & al., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Pansera & al., 2014 (p.221, Table 2, abundance); Zakaria, 2014 (p.3, Table 1, abundance vs. 1960-2000); Zaafa & al., 2014 (p.67, Table I, occurrence); Mazzocchi & al., 2014 (p.64, Table 3, 4, 5, spatial & seasonal composition %); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season, fig.8); Araujo & al., 2016 (p.1, Table 3, abundance, %) ; Zakaria & al., 2016 (p.1, Table 1, 3); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Atique & al., 2017 (p.1, Table 1); El Arraj & al., 2017 (p.272, table 1, 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Chaouadi & Hafferssas, 2018 (p.913, Table III, fig.4, 5: occurrence, abundance, seasonal variation); Belmonte, 2018 (p.273, Table I: Italian zones); Dias & al., 2018 (p.1, Tables 2, 4, 5: vertical distribution, abundance vs. season); Dias & al., 2018 a (p.189, Rem.: p.196); Hure M. & al., 2018 (p.1, Rem: p.11); Yebra & al., 2019 (p.1, Table 4: length, width, juv., F, M); Richirt & al., 2019 (p.3, Table 1, figs. 2, 3, 4, 5, abundance changes vs. years 1998-2014, table 2: diversity index); Acha & al., 2020 (p.1, Table 3: occurrence % vs. ecoregions, Table 5: indicator ecoregions).


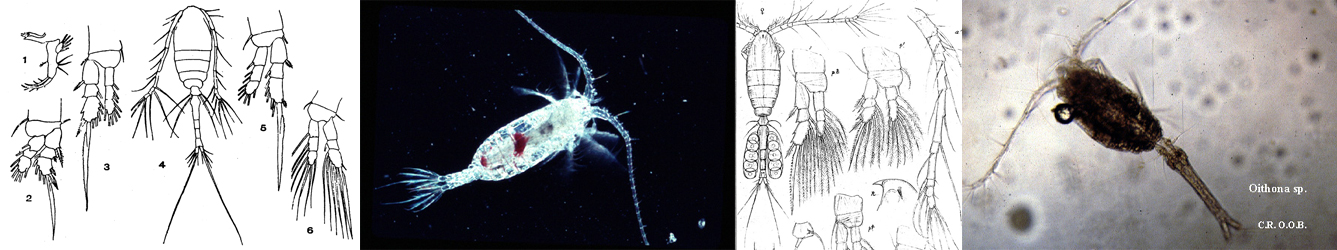

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}