|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Temoridae ( Family ) |
|
|
|
Temora ( Genus ) |
|
|
| |
Temora turbinata (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Calanus turbinatus Dana, 1849;
Temora longicornis : T. Scott, 1894 b (p.76, Pl. 9, fig.13);
Temora tenuicauda Brady, 1899 (p.;
Temora africana Brady, 1914 (p.3, figs. F,M) | | | | Ref.: | | | Giesbrecht, 1892 (p.329, 338, 775, figs.F,M); Giesbrecht & Schmeil, 1898 (p.101, Rem. F,M); Thompson & Scott, 1903 (p.234, 249); A. Scott, 1909 (p.118, Rem.); Pesta, 1912 a (p.48, figs.F,M); 1913 (p.32); Sewell, 1914 a (p.227); Früchtl, 1924 b (p.54); Sars, 1925 (p.193); Bigelow, 1926 (p.293, Rem.); Mori, 1929 (p.175, figs.F); Farran, 1929 (p.209, 258); Wilson, 1932 a (p.106, figs.F,M); Sewell, 1932 (p.246); 1933 (p.27); Dakin & Colefax, 1933 (p.207); Farran, 1936 a (p.109); Mori, 1937 (1964) (p.64, figs.F,M); Dakin Colefax, 1940 (p.93, figs.F,M); Sewell, 1947 (p.165); Tanaka, 1960 (p.49); 1963 (p.13, Rem.F,M); Kasturirangan, 1963 (p.40, figs.F,M); Chen & Zhang, 1965 (p.64, figs.F,M); Gonzalez & Bowman, 1965 (p.249, figs.F,M, Rem.); Vervoort, 1965 (p.99, Rem.); Marques, 1966 (p.5, figs.F,M); Saraswathy, 1966 (1967) (p.79); Owre & Foyo, 1967 (p.69, figs.F,M); 1973; Corral Estrada, 1970 (p.170, figs.F,M); Björnberg, 1972 (p.28, figs., Rem.N); Marques, 1973 (p.240); Goswami & Goswami, 1974 (p.109, fig. caryotypes); Fleminger, 1975 (p.397); Bradford, 1977 (p.131, figs.F,M, Rem.); 1979 (p.598, fig.4: biogéo); Li & Fang, 1983 (p.96, figs.juv.); Zheng Zhong & al., 1984 (1989) (p.241, figs.F,M); Koga, 1984 (p.43, figs.F,M, N, juv., Rem.); Kimmerer & al., 1985 (p.426); Kim & al., 1993 (p.270); Chihara & Murano, 1997 (p.916, Pl.180: F,M); Bradford-Grieve & al., 1999 (p.885, 954, figs.F,M); Bradford-Grieve,1999 b (p.158, figs.F,M, Rem., figs.182, 193); Al-Yamani & Prusova, 2003 (p.71, figs.F,M); Conway & al., 2003 (p.139, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.526, figs.F,M, Rem.); Phukham, 2008 (p.103, figs.F,M); Al-Yamani & al., 2011 (p.51, 54, figs.F,M); Lacuna & al., 2013 (p.99, 105, figs.F,M, Rem.); Kos, 2016 (p.50, figs.F, M, Rem.)
| 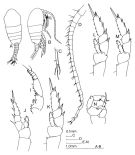 issued from : Bradford-Grieve J.M. in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.157, Fig.110]. Male: A, habitus (dorsal view); B, habitus (dorsolateral view); C, left caudal ramus; D, A1 (right); I, Mxp; J, P1; K, P2; L, P3; M, P4; N, P5. Nota: Similar to T. longicornis but urosome segment 4 longer than the anal segment. - Endopod segments 3 and 4 of Mxp with only 2 inner setae. - Left P5 terminal segment relatively longer, and terminally rather broader.
|
 issued from : Bradford-Grieve J.M. in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p156, Fig.109]. Female: A, habitus (dorsal view); B, lateral view); C, Urosome (dorsal); D, A1; I, Mxp.; J, P1; K, P2; L, P3; M, P4; N, P5. Nota: Similar to T. longicornis but terminal seta 2 of the caudal rami is thicker and asymmetrical. - Endopod segment 3 and 4 of Mxp with only 2 setae. - Swimming legs compact, terminal exopod spine shorter. - Outer spine 2 on exopod segment 3 of P1 with a denticulate margin. - P5 with the inner spine on the terminal segment considerably shorter and thinner than the 2 terminal spines.
|
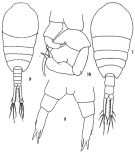 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.20, 7-10]. Female (from E China Sea): 7, habitus (dorsal); 8, P5 (posterior). Male: 9, habitus (dorsal); 10, P5 (anterior).
|
 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.46, fig.8]. Female (from Canarias Is.): 8, P5.
|
 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.47, fig.1]. Male: 1, P5.
|
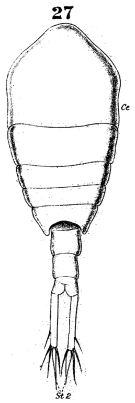 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.38, fig.27]. Female: 27, habitus (dorsal).
|
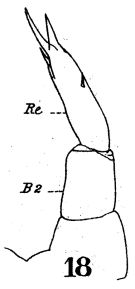 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.17, fig.18]; Female: 18, P5.
|
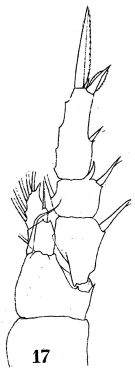 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.17, fig.17]. Male: 17, P1 (anterior view).
|
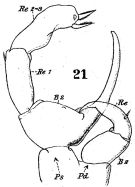 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.17, fig.21]. Male: 21, P5 (anterior view).
|
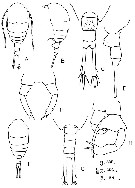 issued from : F.Y. Al-Yamani & I. Prusova in Common Copepods Northwestern Arabian Gulf : Identification Guide. Kuwait Institute for Scientific Research, 2003. [p.70, Fig.25]. Female: A, habitus (dorsal); B, idem (lateral left side); C, urosome (dorsal); D, idem (lateral left side); E, P5. Male: F, habitus (dorsal); G, urosome (dorsal); H, P5.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.32, Figs.3-8]. Female: 3, habitus (dorsal); 6, P5. Nota: The inner marginal spine of the 3rd segment of P5 is shorter than the apical 2 spines. Male: 4, habitus (dorsal); 5, P1; 7, P5; 8, right A1.
|
 issued from : J.M. Bradford in N. Z. Jl Mar. Freshw. Res., 1977, 11 (1). [p.135, Fig.3]. Female (from NE New Zealand): a-b, habitus (dorsal and lateral, respectively); c, genital segment (ventral); d, P5. Nota: The cuticle of body shows the distribution of integumental organs (dots). Rem.: The pattern of integumental organs on females and males found in New Zealand and Hong Kong specimens was very similar, the range of variation being no greater than the variation found among specimens from one area.
|
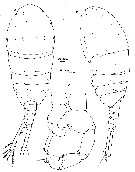 issued from : J.M. Bradford in N. Z. Jl Mar. Freshw. Res., 1977, 11 (1). [p.136, Fig.4]. Male (from NE New Zealand): a-b, habitus (dorsal and lateral, respectively); c, P5 (anterior view). Nota: The cuticle of body shows the distribution of integumental organs (dots).
|
 issued from : J.M. Bradford in N. Z. Jl Mar. Freshw. Res., 1977, 11 (1). [p.134, Fig.2]. Cephalosome of female from coast of Georgia (eastern USA) to show distribution of integumental organs. a: dorsal view; b, lateral view.
|
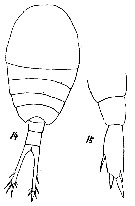 issued from T. Mori in Zool. Mag. Tokyo, 1929, 41 (486-487). [Pl. VI, Figs.14-15]. Female (from Chosen Strait, Korea-Japan): 14, habitus (dorsal); 15, P5.
|
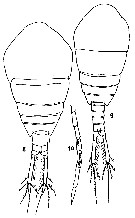 issued from : F. Koga in Bull. Plankton Soc. Japan, 1984, 31 (1). [p.48, Fig.3]. Female (from Fukuoka Bay, Japan): 8, habitus (adult). Male: 9, habitus (adult); 10, distal part of grasping antennule of adult (segments 18-19 and 20-21 fused having a row of small notches).
|
 issued from : F. Koga in Bull. Plankton Soc. Japan, 1984, 31 (1). [p.49, Fig.4]. Female & Male: 6, P1; 12, P2; 17, P3; 21, P4. Female: 27, P5. Male: 24, P5.
|
 issued from : Y. Al-Yamani, V. Skryabin, A. Gubanova, S. khvorov & I. Prusova in Marine Zooplankton Practical Guide for the Northwestern Arabian Gulf, 2, 2011 [p.55, Fig.177]. Female (from Kuwait): a-b, habitus (dorsal and lateral, respectively).
|
 issued from : Y. Al-Yamani, V. Skryabin, A. Gubanova, S. khvorov & I. Prusova in Marine Zooplankton Practical Guide for the Northwestern Arabian Gulf, 2, 2011 [p.55, Fig.178]. Male: a-b, habitus (dorsal and lateral, respectively).
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.190, Fig.64]. Female (from W Malay Peninsula): a-b, habitus (dorsal and lateral, respectively); c, urosome (dorsal); d, P5. Male: e-f, habitus (dorsal and lateral, respectively); g, P5. Body length after the drawings: F = 1.172 mm; M = 0.944 mm.
|
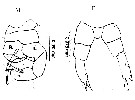 issued from : M.L.D.G. Lacuna, D.C. Sagrado, N.B. España, D.D. Simyunn & R.O. Mejorada in ABAH Bioflux, 2013, 5 (1). [p.112, Figs.17, e; 18, e]. Female (from Mindanao bays); F: P5. Male: M, P5 (L = left leg; R = right leg). Nota: In P5 male is devoid of setules on the terminal setae, however these were seen on the same organism from the Indian and Florida waters.
|
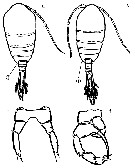 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.919, Pl. 181, fig.283 a-c]. Female : a, habitus (dorsal); b, P5. Male: e, habitus (dorsal); f, P5. Note characteristics numbered 1 into circles, showing the corner of the last thoracic segment.
|
 Issued from : G.S. Brady in Ann. Durban Mus., 1914, Vol. I. [Pl. III, figs. 1-8]. As Temora africana. Female & Male (from Durban Bay, Natal): 1, habitus female (dorsal); 2, Right A1 male; 3, abdomen male; 4, abdomen female; 5, median apical seta of caudal ramus; 6, P4; 7, P5 female; 8, P5 male. Nota female: - Cephalothorax very broad, tapering gradually backwards, rounded off posteriorly. - Abdomen 3-segmented, much narrower than the cephaloyhorax. - Caudal rami very long and slender, equal in length to the entire abdomen, bearing 4 short apical setae, the principal one lancet-shaped; both caudal rami and setae non plumose. - A1 reaching nearly to the extremity of the cephalothorax, 24-segmented and very sparingly setiferous. - Inner branches (= endopod) of all legs bi-articulate. - P5 unbranched, simple, 3-segmented and bearing 2 short apical spines. Nota male: -Right A1 scarcely at all swollen geniculated between the 18th and 19th segments, very slightly setiferous; - P5 strongly prehensile, similar to those of T. longicornis, but stoiter and slightly different in spinous armature
|
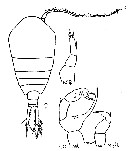 Issued from : M.S. Kos in Zoological Institut RAS, St. Petersburg, 2016, 179, [p.51, Fig.23]. Temora turbinata female and male.
| | | | | Compl. Ref.: | | | Carl, 1907 (p.16); Sewell, 1912 (p.353, 366); Wilson, 1932 (p.33); Oliveira, 1945 (p.191); Sewell, 1948 (p.323, 514); C.B. Wilson, 1950 (p.343); Krishnaswamy, 1953 (p.124); Kott, 1957 (p.5, 8); Yamazi, 1958 (p.150, Rem.); Deevey, 1960 (p.5, Table II, annual abundance); Zoppi, 1961 (p.219); Ganapati & Shanthakumari, 1962 (p.8, 16); Cervigon, 1962 (p.181, tables: abundance distribution); De Decker, 1964 (p.16, 25, 30); De Decker & Mombeck, 1964 (p.14); Neto & Paiva, 1966 (p.26, Table III, annual cycle); Reeve, 1970 (p.894, Rem.: p.906); Park, 1970 (p.476); Deevey, 1971 (p.225); Binet & al., 1972 (p.71); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %), Table IV: seasonal abundance; Brodsky, 1972 (p.256); Binet, 1973 (p.77); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Patel, 1975 (p.659); Grahame, 1976 (p.219, Table X, III); Frontier, 1977 a (p.15); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Weikert, 1977 (p.351, Rem.: p.353, exuviae); Carter, 1977 (1978) (p.36); Ikeda, 1977 d (p.263, feeding); Paffenhöfer & Knowles, 1978 a (p.143, feeding); Stephen & Iyer, 1979 (p.228, tab.1, 3, 4, figs.3, 4); Binet, 1979 (p.400); Dessier, 1979 (p.100, 201, 206); Chen Q-c, 1980 (p.795); Andronov & Maigret, 1980 (p.71, Table 2, 3); Hirota 1981 (p.19, Table 1, length-weight-CHN); Sreekumaran Nair & al., 1981 (p.493, fig.2), Vives, 1982 (p.293); Turner & Dagg, 1983 (p.16, 22); Dessier, 1983 (p.89, Tableau 1, Rem., %); Turner, 1984 a (p.73, Table 1, fig.4, 7: pellet contents); De Decker, 1984 (p.316, 365: chart); Binet, 1984 (tab.3); Stephen, 1984 (p.161, Distribution vs thermocline & geographic); Paffenhöfer & al., 1984 (p.995, figs.12-15, patches); Kimmerer & McKinnon, 1985 (p.149); Brenning, 1985 (p.5, spatial distribution/TS); 1985 a (p.28, Table 2); Binet, 1985 (p.85, tab.3); Paffenhöfer, 1985 (p.104, distribution); Sarkar & al., 1986 (p.178); Madhupratap & Haridas, 1986 (p.105, tab.1); Ambler, 1986 a (p.957, fig.7, selectivity ); Dagg & Walser, 1986 (p.1066, Chl. a concentration vs. fecal pellets); Diouf & Diallo, 1987 (p.260); Paffenhöfer & al., 1987 (p.403, Rem.: patch); Jacoby & Greenwood, 1988 (tab.1); Lozano Soldevilla & al., 1988 (p.59); Dessier, 1988 (tabl.1); Mitra & al., 1990 (fig.3); Suarez & al., 1990 (tab.2); Othman & al., 1990 (p.561, 563, Table 1); Yoo, 1991 (tab.1); Dai & al., 1991 (tab.1); Samba Diouf, 1991 (p.104); Lindo, 1991 (tab.3); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Verheye & al., 1994 (p.155); Hwang & Turner, 1995 (p.207, Table 1, 2, figs. 1, 2, 3: swimming behaviour); Roff & al., 1995 (p.165, Table 1, 3, 5, nauplii development times, nauplii copepodites growth rates, naupllii vs. bacteriophagy/picoplankton, naupliar production); Webber & Roff, 1995 (tab.1); Shih & Young, 1995 (p.74); Webber & al., 1996 (tab.1); Ramaiah & al., 1996 (p.3); Padmavati & Goswami, 1996 a (p.85, fig.3, Table 4, vertical distribution); Madhupratap & al., 1996 (p.866); Kotani & al., 1996 (tab.2); Suarez-Morales & Gasca, 1997 (p.1525); Sharaf & Al-Ghais, 1997 (tab.1); Park & Choi, 1997 (Appendix); Ramaiah & Nair, 1997 (tab.1); Padmavati & al., 1998 (p.349); Achuthankutty & al., 1998 (p.1, Table 2, fig.6: salinity range, seasonal abundance vs monsoon); Hwang & al., 1998 (tab.II); Lopes & al., 1998 (p.195); Alvarez-Cadena & al., 1998 (tab.1,2,3,4); Hopcroft & al., 1998 (tab.2); Hsieh & Chiu, 1998 (tab.2); Wong & al, 1998 (tab.2); Suarez-Morales & Gasca, 1998 a (p.111); Mauchline, 1998 (tab.8, 24, 25, 47, 51); El-Serehy, 1999 (p.172, Table 1, occurrence); Lopes & al., 1999 (p.215, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Razouls & al., 2000 (p.343, Appendix); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Ueda & al., 2000 (tab.1); Suarez-Morales & Gasca, 2000 (1247, tab.1); Lopez-Salgado & al., 2000 (tab.1); Alvarez-Silva & Gomez-Aguirre, 2000 (p.163: tab.2); Dalal & Goswami, 2001 (p.22, fig.2); Lo & al., 2001 (1139, tab.I); Ara, 2001 b (p.121); 2002 (p.399, temporal variability, production); Greenwood & al., 2002 (p.17, Table 2); Beaugrand & al., 2002 (p.179, figs.5, 6); Dunbar & Webber, 2003 (tab.1); Osore & al., 2003 (p.69); Hwang & al., 2003 (p.193, tab.2); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.326, tab.1); Hsieh & al., 2004 (p.397, tab.1); Rezai al., 2004 (p.490, tab.2, p.495, tab.8); Lo & al.*, 2004 (p.218, tab.1, fig.6); Lo & al., 2004 (p.468, tab.2); Krumme & Liang, 2004 (p.407, tab.1); Lan & al., 2004 (p.332, tab.1, tab.2); Ara, 2004 (p.179, figs. 3,4,5); Yin & al., 2004 (p.3); Lo & al., 2004 (p.89, tab.1); Kazmi, 2004 (p.229); Shimode & al., 2005 (p.113 + poster); Dias & Bonecker, 2005 (p.100 + poster); Waggett, 2005 (p.17, Table 3.3, behaviour); Rezai & al., 2005 (p.157, Table 5: spatial & temporal variations); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: stations group); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Hwang & al., 2006 (p.943, tabl. I) ; Fang & al., 2006 (p.224, metals concentration); Sterza & Fernades, 2006 (p.95, Table 1, occurrence); Dias & Araujo, 2006 (p.68, Rem., chart); Dur & al., 2007 (p.197, Table IV); Rakhesh & al., 2008 (p.154, abundance vs stations); McKinnon & al., 2008 (p.844: Tab.I, p.848: Tab. IV, fig.7); Jerling, 2008 (p.55, Tabl.1); Morales-Ramirez & Suarez-Morales, 2008 (p.514); Selifonova & al., 2008 (p.305, Tabl. 2); Fernandes, 2008 (p.465, Tabl.2); Ohtsuka & al., 2008 (p.115, Table 4, 5, in Japan Harbors); Waggett & Buskey, 2008 (p.111, fig.3, Table 1); Lan Y.-C. & al., 2008 (p.61, Table 1, % vs stations, Table 2: indicator species, Rem.: p.72); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); Tseng L.-C. & al., 2008 (p.402, Table 2); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons, fig.8, table 3: indicator species); C.-Y. Lee & al., 2009 (p.151, Tab.2); Miyashita & al., 2009 (p.815, Tabl.II); Zhang W & al., 2009 (p.261); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Hwang & al., 2009 (p.49, fig.4, 5); Zhang G.-T. & al., 2010 (p.492, fig.1, Table1, 2, Rem.); Cornils & al., 2010 (p.2076, Table 3, Fig.5); Fazeli & al., 2010 (p.153, Table 1); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); W.-B. Chang & al., 2010 (p.735, Table 2, 4, fig.5, abundance); Hsiao & al., 2010 (p.179, Table I, II, III, trace metal concentration); Shanthi & Ramanibai, 2011 (p.132, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I: as Tem. turbinate, lapsus calami); Hsiao & al., 2011 (p.232, Table 1: abundance, %, Table 2: trace metal concentration, vs transect); Zhang G.-T. & Wong, 2011 (p.277, fig.6, 7, 8, abundance, indicator); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Hsiao & al., 2011 (p.317, Table 2, 3, fig.6, indicator of seasonal change); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Tseng & al., 2011 (p.301, adaptability vs T. discaudata); Beltrao & al., 2011 (p.47, Table 1, density vs time); Guo & al., 2011 (p.567, fig.5, table 2, abundance vs spatial distribution, indicator); Magris & al., 2011 (p.260, abundance, interannual variability); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Chen M. & al., 2011 (p.1075, Fig.8, annual abundance variation); Glushko & Lidvanov, 2012 (p.138, Tableau 1, 3); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Jose & al., 2012 (p.20, fig.3 a,b,c: % vs monsoon); Johan & al., 2012 (2013) (p.1, Table 1); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Tseng & al., 2012 (p.621, Table 3: abundance, indicator species); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Almeida LR. & al., 2012 (p.13, Table 1, abundance); Naz & al., 2012 (p.61, Table 4, relative abundance); Gubanova & al., 2013 (in press, p.4, Table 2); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Hsiao & Fang, 2013 (p.175, Hg bioaccumulation); Tseng & al., 2013 (p.507, seasonal abundance); Tseng & al., 2013 a (p.1, Table 3, 5, abundance); Jagadeesan & al., 2013 (p.27, Table 3, 4, 5, 6, fig.11, seasonal abundance); Rakhesh & al., 2013 (p.7, Table 1, 4, abundance vs stations); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Hirai & al., 2013 (p.1, Table I, molecular marker); Suzuki, K.W. & al., 2013 (p.15, Table 2, 3, 4, estuaries, annual occurrence); Lidvanov & al., 2013 (p.290, Table 2, % composition); Varadharajan & Soundarapandian, 2013 (p.2: occurrence vs stations); Chen M. & al., 2013 (p.47, clearance & ingestion rate); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Ortega & al., 2014 (p.495, Table I, %); Fuentes-Reinés & Suarez-Morales, 2015 (p.369, Table 1, Rem. p.374); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season, fig.8); Nakajima & al., 2015 (p.19, Table 3: abundance); Araujo & al., 2016 (p.1, Table 3, 5, abundance, %); Zakaria & al., 2016 (p.1, Table 1, Rem.); Ortega & al., 2017 (p.123, fig. 1, 6, table 2, abundance, %) ; Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Ohtsuka & Nishida, 2017 (p.565, 518, Table 22.1); Dias & al., 2018 (p.1, Table 2: vertical distribution, abundance vs. season); 2018 a (p.189, fig.11: seasonal variation, Rem.: p.196). | | | | NZ: | 15 | | |
|
Distribution map of Temora turbinata by geographical zones
|
| | | | | | | | | | | | | | |  issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.68]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.68].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S).
Nota: sampling 237 specimens. |
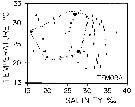 issued from : M.R. Reeve in Bull. mar. Sc., 1970, 20 (4). [p.902, Fig.4]. issued from : M.R. Reeve in Bull. mar. Sc., 1970, 20 (4). [p.902, Fig.4].
Temperature-salinity diagram for Temora turbinata in Biscayne Bay (Florida, Miami). |
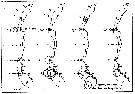 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 34. Jahrgang 1985. Mat.-nat. wiss. Reihe, 6. [p.12, Fig.9]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 34. Jahrgang 1985. Mat.-nat. wiss. Reihe, 6. [p.12, Fig.9].
Spatial distribution for Temora turbinata and Temora longicornis from 8° S - 26° N; 16°- 20° W, for different expeditions (V1: Dec. 1972-Jan. 1973; V2: Feb./mar. 1973; VI: May 1974; IV: Jun./Jul. 1972). |
 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 34. Jahrgang 1985. Mat.-nat. wiss. Reihe, 6. [p.12, Fig.9]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 34. Jahrgang 1985. Mat.-nat. wiss. Reihe, 6. [p.12, Fig.9].
T-S Diagram for Temora turbinata and Temora longicornis from 8° S - 26° N; 16°- 20° W.
SO: Southern Surface Water (S °/oo: 34,50; T°C: 29,0); ND: Northern Water of the Surface Layer (S °/oo: 37,5; T°C: 21,0); SD: Southern Deep Water of the surface layer (S °/oo: 35,33; T°C: 13,4). (see commantary in Temora stylifera and Brenning, 1985 a, p.6) |
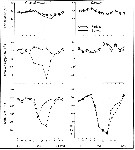
 issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.8, Fig.6]. issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.8, Fig.6].
Salinity ranges for T. turbinata in coastal and estuarine waters of Goa (India).
Shaded area indicates the range of higher abundance. |
 issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.6, Fig.5]. issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.6, Fig.5].
Monthly occurrence of Temora turbinata in coastal (black circle) in front of Goa and and the Mandovi estuary (clear circle). |
 issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.3, Fig.2]. issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.3, Fig.2].
Hydrography of the coastal station in front of Goa and Mandovi estuarine station (W India).
Nota: The seasons are arbitrarily classified into the southwest monsoon (June-September), postmonsoon (October-January) and premonsoon (February-May). |
 Issued from : S.-H. Hsiao, J.-S. Hwang & T.-H. Fang in Crustaceana, 2010, 83 (2). [p.184, Table I]. Issued from : S.-H. Hsiao, J.-S. Hwang & T.-H. Fang in Crustaceana, 2010, 83 (2). [p.184, Table I].
Copepods collected from the sea around northern Taiwan and from southern East China Sea extending to the Okinawa Trough, to study possible spatial heterogeneity. The content of the same metal shows considerable variation both intra- and inter-specifically. The metal concentrations in males are higher than in females. Copepod metal quota display spatial variation: coastal water > southern East China Sea > Kuroshio water, suggesting that the metal contents of copepods are influenced by the water quality of their marine environment. |
 Issued from : D.-h. Guo, J.-q. Huang & S.-j. Li in Cont. Shelf Res., 2011, 31. [p.572, Fig.5]. Issued from : D.-h. Guo, J.-q. Huang & S.-j. Li in Cont. Shelf Res., 2011, 31. [p.572, Fig.5].
Distribution of Temora turbinata abundance (ind/m3) during August 31 to September 3, 1988 (a); August 29 to September 8, 1994 (b); July 4 to 15, 2005 (c); and June 20 to July 2, 2006 (d).
Nota: The same sampling procedure was used for all cruises. Zooplankton samples were collected by vertical tows from 3 m of the bottom to the surface (plankton net : 505 µm mesh size).
The aim of the research is the relationship between copepods and associated water masses in the southern Taiwan Strait. |
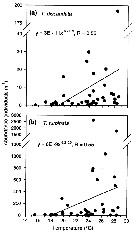 Issued from : L.-C Tseng, R. Kumar, Q.-C. Chen & J.-S. Hwang in Hydrobiologia, 2011, 666. [p.313, Fig.9]. Issued from : L.-C Tseng, R. Kumar, Q.-C. Chen & J.-S. Hwang in Hydrobiologia, 2011, 666. [p.313, Fig.9].
Densities (ind./ m3) of T. discaudata (a) and T. turbinata (b) as a function of seawater temperature (°C) in the southern margin of the East China Sea in the vicinity of Nuclear Power Plant-I and II of northern Taiwan. |
 Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6]. Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6].
Variations in the most abundant copepod species (mean ± SE) along the transect in the boundary waters between the northern part Taiwan Strait and the East China Sea i March (black bar) and October (grey bar) 2005 (Mann-Whitney U test, sig. **P <0.01.
See drawings of Hydrological conditions and superficial marine currents in Calanus sinicus. |
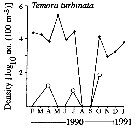
 Issued from : C. de O. Dias, Carvalho P.F. de, A.C.T Bonecker & S.L.C. Bonecker in Ocean Coastal Management, 2018, 161. [p.198, Fig.11]. Issued from : C. de O. Dias, Carvalho P.F. de, A.C.T Bonecker & S.L.C. Bonecker in Ocean Coastal Management, 2018, 161. [p.198, Fig.11].
Variation of seasonal mean (± SD) density (Log ind.m3. of Temora turbinatain Guanabara bay (22°50'S, 43°10'W).
Zooplankton collected from January 2012 to December 2013 in front of Icaral beach during the flood time, by horizontal subsurface hauls (1 m depth), net of 200 µm mesh aperture.Issued from : C. de O. Dias, Carvalho P.F. de, A.C.T Bonecker & S.L.C. Bonecker in Ocean Coastal Management, 2018, 161. [p.198, Fig.11].
Variation of seasonal mean (± SD) density (Log ind.m3. of Temora turbinatain Guanabara bay (22°50'S, 43°10'W).
Zooplankton collected from January 2012 to December 2013 in front of Icaral beach during the flood time, by horizontal subsurface hauls (1 m depth), net of 200 µm mesh aperture.
For the authors, the study shows that the poor conditions in the Guanabara Bay are aggravated during periods of rainfall, and that the increase of micriobiological indictors affectes the biodiversity of organisms in the mesoplanktonic community.
The copepods Temora turbinata and Acartia tonsa presented the highest densities throughout the study perod. The dominance in the Bay has changed considerably in recent years; untill two decades ago Temora turbinata and Acartia tonsa were not even registered in the Bay, probably explained by introduction through ship ballast water at ports on the Brazilian coast. These two species have been reported as dominant species which shows their great capacity for adaptation and reproduction in a short time period.
Acartia tonsa has been reported on Guanabara Bay since 1990s, while T. turbinata in 2000 (Guenther & al., 2012; Bonecker & al., 2012).
For the authors, the restructuring of the local community may be related to the constant degradation of the environment, particularly in the inner areas of the bay, reflecting in the entire ecosystem (Gomes, 2007). |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcrift in Aquat. microb Ecol., 1995, 9. [p.171, Figs. 1, 2]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcrift in Aquat. microb Ecol., 1995, 9. [p.171, Figs. 1, 2].
Examples of results of feeding experiments. (photomicrographs of tropical marine crustacean from Jamaica) fed fluorescently labelled bacteria (FLB) at concentrations of 1.5 to 2.5 x 106. Fig.1: Temora turbinata stage 4 nauplius showing concentrations of FLB in foregut (live, partly squashed preparation).
Fig.2: same species, nauplius stage 5 showing concentrations of FLB in hindgut (live preparation).
FLB: Escherichia coli (± 0.7 µm3 cell volume); experimental concentrations used 1.5 to 2.5 x 106cells ml-1.
Note little yellow spot in the gut.
The authors show of the 11 species examined (Centropages velificatus, Acartia lilljeborgi, Paracalanus aculeatus, Temora stylifera, Undinula vulgarius, Euchaeta marina, Clausocalanus spp., Onaea sppp., Corycaeus spp.), 7 showed clear evidence of feeding on FLB (see table 5), with 92% of their nauplii ingesting FLB, clearly visible in the guts of many nauplii, and usually comprisised a discrete feeding (foregut) or 'fecal' (hindgut) bolus of cells. After only 30 min, fecal pellets were being produced already filled with FLB. Presence of FLB in guts cannot be interpreted as incidental feeding or 'drinking', because 4 species failed to ingest them (Centropages velificatus, Euchaeta marina, Clausocalanus sp., Corycaeus sp.). The very earliest stages may not ingest bacteria or other particles because they may have yolk reserves (for example Temora turbinata N1 nauplii did not ingest FLB, but later stage nauplii did). |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.170, Table 5].
Summary observations on naupliar bacterovory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Kingston Harbour (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB. |
 Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.172, Table 6]. Issued from : J.C. Roff, J.T. Turner, M.K. Webber & R.R. Hopcroft in Aquat. microb. Ecol., 1995, 9. [p.172, Table 6].
Summary observations on adult female Temora turbinata bacterivory on FLB (fluorescently labelled bacteria) a partir Escherichia coli (±0.7 µm3 cell volume) in Kingston Harbour (Jamaica).
Fluorescence levels: 0 = no materal for observation; - = no fluorescence observed; + = low fluorescence due to few FLB observe; * = highly fluorescence due to many FLB.
Among species examined, adult females of this species and others as Paracalanus aculeatus) and early copepodites of Oithona spp. (p.171, Fig.4) showed clear evidence of bacteriovory. |
 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2]. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.209: Table 1, p.210: Table 2].
Table 1: Experimental protocol.
Table 2:Swimming versus non-swimming behaviour, and behavioural transitions per minute. Means (bold type), ranges, and standard deviations (SD). Number of repeated observations = 500.
Copepods collected in surface net from waters near Pitou (NE coast Taiwan: 25°N, 122°E) in daylight in July 1993.
Compare with Oithona nana, O. similis, Oncaea venusta, Macrosetella gracilis and Undinella vulgaris var. taiwanicus by the same authors.
Nota: This species swam nearly continuously for extended periods, with almost no periods of rest, and almost no transitions from swimming to non-swimming.. Perioods of non-swimming and behavioural transitions/minute were so few that it was impraticable to calculate standard deviations. |
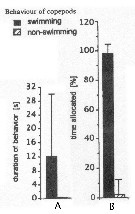 Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2]. Issued from : J.-S. Hwang & J.T. Turner in P.S.Z.N. 1: Mar. Ecol., 1995, 16 (3). [p.211: Figs.1, 2].
Fig.1: A, Mean durations periods (seconds) of swimming versus non swimming behaviour by Temora turbinata from north-east coast of Taiwan (near Pitou) in daylight in July 1993.
Fig.2: B, mean percentages of time allocated to swimming versus non-swimming behaviour by Temora turbinata. |
| | | | Loc: | | | sub-Antarct. (SW Pacif.), Agulhas Bank, South Africa (E & W), Namibia, Angola, Baia Farta, Congo, G. of Guinea, off Lagos, Ivorian shelf, Casamance, Dakar, Morocco-Mauritania, Banc d'Arguin, Canary Is., Brazil (S, Paranagua Bay, off Rio de Janeiro, Campos Basin, Vitoria Bay, Vitoria-Cabo de Sao Tomé, Camamu, Guarairas Lagoon, Mucuri estuary, Cananeia Lagoon, off Natal), Barbados Is., Venezuela, Cariaco Basin, Colombia (Laguna Navio Quebrado), Caribbean Sea, Caribbean Colombia, Bahia de Mochima (Venezuela), Yucatan, Porto Rico, Jamaica (Kingston Harbour), E Costa Rica, G. of Mexico, Aransas Ship Channel, off W Mississipi River mouth, lagunas de Veracruz, Cuba, Florida, Sargasso Sea, off Bermuda: Station S (32°10N, 64°30W), Delaware Bay (outside), Chesapeake Bay, Long Island, Woods Hole, G. of Maine, E Medit. (W Egyptian coast), W Black Sea, Sharm El-Sheikh, Red Sea, Gulf of Oman, Arabian Gulf, UAE coast, Kuwait, Arabian Sea, Karachi coast, Sri-Lanka, Natal, Madagascar (Nosy Bé), India (Mangalore coast, Saurashtra coast, W, Goa - Gujarat, off Cochin, Bombay, Madras, Burhabalanga estuary, Godavari region, Kakinada Bay, Lawson's Bay, G. of Mannar, Manamelkudi, Palk Bay, Mandarmani, Hooghly estuary), E India, Bay of Bengal, Burma (coast), Kurau River, W Malay Peninsula (Andaman Sea), Straits of Malacca, G. of Thailand, Indonesia-Malaysia, Sarawak: Bintulu coast, Ambon Bay, Tioman Is., SW Celebes, SE Philippines (Mindanao bays), Viet-Nam (Cauda Bay), G. of Tonkin, Hainan (Sanya Bay), Hong Kong, China Seas (Yellow Sea, East China Sea, South China Sea, Changjiang River estuary, Xiamen Harbour), Taiwan Strait, Taiwan (S, E, SW, W, Tapong Bay, Bali, NW, off Danshuei River, N, NE, Pitou, Mienhua Canyon, Kueishan Is.), S Korea, Japan Sea, Japan (Kyushu: Ariake Bay, Sagami Bay, Suruga Bay, Tokyo Bay, Setonaikai, Tanabe Bay, south estuaries: Chikugo, Midori & Kuma Rivers), off SE Japan, Australia (Townsville: indhore waters, Great Barrier, Brisbane River estuary, Moreton Bay, Melbourne, Shark Bay, North West Cape, G. of Carpentaria), Tasman Sea, New Caledonia, New Zealand (NW North Island), E Pacif., G. of California (in Suarez-Morales & Gasca (1998 a) | | | | N: | 304 | | | | Lg.: | | | (16) F: 1,25-1,1; M: 1,2; (26) F: 1,57-1,41; M: 1,43-1,32; (45) F: 1,6-1,4; M: 1,5-1,3; (47) F: 1,6-1,46; M: 1,5-1,4; (66) F: 1,22; 1,1; M: 1,05; (91) F: 1,6-1,1; M: 1,5-1,1; (104) F: 1,5; M: 1,4; (180) F: 1,34-1,1; M: 1,13-1,08; (290) F: 1,3-1,5; M: 1,25-1,35; (334) F: 1,5; M: 1,4; (336) F: 1,2-1,1; M: 1,2-1,1; (354) F: 1,7-1,31; M: 1,3-1,18; (530) F: 1,4; M: 1,3; (581) [N.Z] F: 1,61-1,2; M: 1,56-1,01; [Hong Kong] F: 1,15-1,05; M: 1,09-0,93; (786) F: 1,16; 1,45-1,34; M: 1,28-1,18; (795) F: 1,4; M: 1,3; (870) F: 0,95-1,45; M: 0,89-1,37; (937) F: 1,33-1,46; M: 1,28-1,31; (991) F: 1,05-1,61; M: 0,93-1,56; (1085) F: 0,9-1,61; M: 0,9-1,4; (1140) F: 1,64; M: 1,71; (1220) F: 1,1-1,6; M: 1,1-1,5; (1308) F: 1,01-1,53; M: 1,08-1,44; {F: 0,90-1,70; M: 0,89-1,71} | | | | Rem.: | Epipelagic. Coastal, estuaries.
Sampling depth (sub-Antarct.) : 0-50 m. Sargasso Sea: 0-500 m (Deevey & Brooks, 1977, Station "S"); After Brady (1914, p.3) this species is almost a fac-simile of the northern species T. longicornis.
For Vervoort (1965, p.99) this species is widely distributed in the tropical, subtropical and temperate parts of the Indian Ocean and the Indo-Westpacific area. In the Indian and Pacific Oceans it goes about as far south as 35°S; in the Pacific Ocean it goes as far as north as 40°N. In the Atlantic the distribution is more or less erratic, it occurs chiefly in the tropical and sub-tropical eastern Atlantic; in the West Atlantic it reaches the Gulf of Maine.
The coastal and neritic geographical distribution of this estuary species seems to be correlated to the major subtropical surface currents during the oligocene (cf. Bradford, 1979, figs.4 et 7).
According to Owre & Foyo (1967, p.69) distributional records indicate that it is more abundant close to land masses in the Florida region.
Some confusions have been possible between this species and T. longicornis (cf. Corral Estrada, 1970, p.171).
Ohtsuka & Nishida (2017, p.578) point to the Japanese coasts adjacent to the Kuroshio Current are distinguished by warm-water species T. turbinata. | | | Last update : 19/06/2023 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 17, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
