|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Tortanidae ( Family ) |
|
|
|
Tortanus ( Genus ) |
|
|
|
Eutortanus ( Sub-Genus ) |
|
|
| |
Tortanus (Eutortanus) dextrilobatus Chen & Zhang, 1965 (F,M) | |
| | | | | | | Ref.: | | | Chen & Zhang, 1965 (p.118, 128, figs.F,M); Ohtsuka & al., 1992 (p.112, figs.F,M, Rem.); Ferrari & Ueda, 2005 (p.346, figs.juv,F,M) | 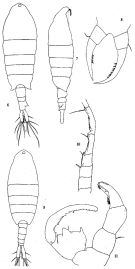 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.53, 6-11]. Female (from Amoy, Formosa Strait): 6, habitus (dorsal); 7, idem (lateral right side); 8, P5 (anterior). Nota: Head and 1st thoracic segment separate, 4th and 5th fused. - Postero-lateral angles of last thoracic segment protuberant and symmetrical. - Urosome 3-segmented. - Anal segment longer than preceding segment, asymmetrical, its upper right side with a triangular process. Furcal rami large, 4 times as long as wide, right furcal seta longer than the left. P5 not very symmetrical (left longer than right), inner margin of both legs with bristles. Male: 9, habitus (dorsal); 10, geniculated portion of right A1; 11, P5 (posterior). Nota: Urosome 5-segmented, anal segment very short. - Furcal rami like those of female. - Right A1 geniculate, 17-segmented, anterior border of 13th-15th armed with denticles. - Right P5 chelate
|
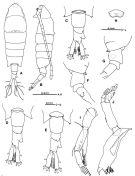 issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.115, Fig.2]. Female (from Somjin River, S Korea): A, habitus (dorsal); B, idem (lateral left side); C-E, 5th pedigerous somite and urosome (dorsal); F, idem (lateral right side); G, idem (lateral left side); H, genital operculum; I, A2; J, Md. Nota: Cephalosome with transverse groove medially and dorsal median protuberance posteriorly. - Head and 1st thoracic segment separate, 4th and 5th fused (suture line slifghtly visible in lateral view). - Eye large, reddish. - A1 17-segmented. - A2 with coxa and basis partly fused, coxa with small medial seta, basis with 2 medial setae of unequal lengths; basis and endopod completely fused; endopod 2-segmented, proximal with 1 seta and row of spinules subterminally, distal with 6 setae terminally and row of spinules subterminally; exopod 3-segmented, proximal unarmed, middle with 3 setae, distal with 2 terminal setae. - Endopod of Md: 2-segmented, proximal segmnt without seta, distal with 5 setae; exopod 5-segmented incompletely fused with 3 rows of minute spinules. Urosome 3-segmented, 2nd urosomal segment with posterior dorsal protuberance on right side, 3rd segment having large irregular dorsolateral process on anterior right side. Anal segment incompletely fused with left caudal ramus and almost completely with right one. - variation: The dorsolateral process on the right side of the 3rd urosomal somite is slightly variable in shape
|
 issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.116, Fig.3]. Female: A, A1; B, Mx1; C, Mx2; D, Mxp; E, P1. Nota: Mx1 with basis, endopod and exopod completely absent; praecoxal arthrite stout, with 12 plumose setae and 1 minute setule (indicated by arrowhead); coxal endite with 3 stout, spinulose setae terminally. Mx2 with proximal and distal endites of praecoxa and coxa having 1, 2, 1 and 3 setae, respectively; basal endite with 1 developed and 2 minute setae; endopod with setal formula 1, 2, 2, 2 (1 seta on 2nd segment rudimentary). Mxp with setal formula on praecoxal and coxal endites: 0, 2, 2, 1; basis unarmed; endopod 1-segmented with 3 inner plumose setae and 1 outer seta.
|
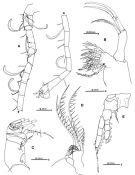
 issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.118, Fig.4]. Female: A, P2 (anterior); B, P3 (anterior); C, P4 (anterior); D, P5 (anterior). Nota: Right and left coxal segments of P5 fused Male: E, habitus (dorsal); F, 5th pedigerous somite and urosome (lateral right side, relatively stout sensilla indicated by arrows); G, idem (dorsal). Nota: 4th and 5th pedigerous somites incompletely fused. Urosome 5-segmented. Genital somite with opening on left lateral side. 1st abdominal somite with 2 relatively stout sensilla ventrolaterally (indicated by arrows in fig.4 F. Anal somite separate from caudal rami. Right caudal ramus slightly longer than left.
|
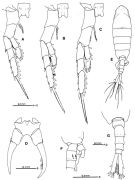
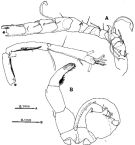 issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.119, Fig.5 A-B]. Male: A, right A1; B, P5 (anterior). Right A1 geniculate, indistinctly 16-segmented but consisting of 14 fully separated segments (geniculation between 14th and 15th). Right and left coxal segments of P5 fused; basis and 1st exopod segment of right leg fused; basis of left leg with outer seta subterminally, proximal exopod segment with short, thick spine subterminally, distal exopod segment with tuft of hairs and small seta on proximal inner margin, tuft of spinules along distal inner margin and 1 medial inner and 2 terminal short spines, terminal outer portion with 13 transverse and 3 longitudinal ridges. Variation: The number of the ridges along the inner margin of right P5 is 24 or 25 (number of individuals examined: 3).
|
 issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.117]. Female: Seta (Arabic numerals) and spine (Roman numerals) of swimming legs P1 to P4.
|
 Issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [p.130 ]. Comparison of distinguisehed features between T. dextribobatus, T. derjugini, denticulatus.
| | | | | Compl. Ref.: | | | Chen Q., 1983 (p.133); Dai & al., 1991 (tab.1); Myung & al., 1994 (tab.1); Shih & Young, 1995 (p.74); Ohtsuka & al., 1995 (p.159); Cordell & al., 1997 (p.2, 7, invasive species, % change 1980-1997); Orsi & Ohtsuka, 1999 (p.128); Bollens & al., 2002 (p.87); Hooff & Bollens, 2004 (p.167, predatory impact); Ohtsuka & al., 2008 (p.115, Table 6); Cordell & al., 2008 (p.753); Bollens & al., 2011 (p.1358, Table III, fig.8); Sakaguchi & al., 2011 (p.18, Table 2, occurrences); Winder & al., 12011 (p.749, figs.2, 6, invasion chronology); Moon & al., 2012 (p.1, Table 1); Soh & al., 2013 (p.723: Rem.); Zhang H. & al., 2013 (p.57, Rem. p.58); Seo & al., 2013 (p.448, Table 1, as dextrirobatus, occurrence); Park E.-O. & al., 2013 (p.165, Fig.5, distribution vs stations); | | | | NZ: | 3 | | |
|
Distribution map of Tortanus (Eutortanus) dextrilobatus by geographical zones
|
| | | | | | 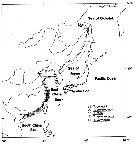 issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.120, Fig.6]. issued from : S. Ohtsuka, Y.H. Yoon & Y. Endo in The J. Oceanol. Soc. Korea, 1992, 27 (2). [p.120, Fig.6].
Schematic illustration of distribution patterns of five sepecies of the subgenus Eutortanus. |
 Issued from : M. Winder, A.D. Jassby & R.M. Nally in Ecology Letters, 2011, 14. [p.753, Fig.3, f]. Issued from : M. Winder, A.D. Jassby & R.M. Nally in Ecology Letters, 2011, 14. [p.753, Fig.3, f].
Historical abundances of introduced species in the upper San Francisco Estuary between 1972 and 2009.
Adult stages are shown because immature stages were not continuously identified. |
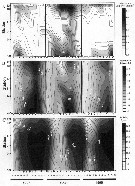 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.171, Fig.2]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.171, Fig.2].
A: Abundance of Tortanus dextrilobatus (Log [individuals + 1] m3); all copepodid stages combined.
B: Water column average values temperature, and C: salinity obtained simultaneously with zooplankton collections in the San Francisco Estuary, 1997 to 1999.
Open circles on x-axis represent sampling dates; dark circles on y-axis are sampling locations (plotted as distance) as represented in the map (fig.1) including South Bay, Central Bay and San Pablo Bay (north). |
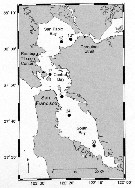 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.168, Fig.1]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.168, Fig.1].
Map of San Francisco Estuary, including South Bay, Central Bay, and San Pablo Bay.
Numbers refer to the US Geological Survey historical stations sampled for zooplankton abundance and composition. |
 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.172, Fig.3]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.172, Fig.3].
GFunctional response of adult females feeding on Oithona davisae (CV-CVI) at (A, C) 14°C (solid line) and at (B, D) 19°C (dashed line).
Values expressed as (A,B) mean number individuals consumed per predator and per day (black point) and (C,D) % body carbon consumed per predator and per day (black triangle). Error bars represent ±1 SD. |
 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.172, Fig.4]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.172, Fig.4].
Functional response of adult females feeding on Acartia (Acartiura) sp. (CV-CVI) at (A, C) 14°C (solid line) and at (B, D) % body carbon consumed per predator and per day (black point) and body carbon consumed per predator and per day. Error bars represent ±1 SD. |
 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.173, Table 1]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.173, Table 1].
Functional response parameters (based on Ivlev's equation) of adult females feeding on adult female copepodids of two prey items at two temperatures (Temp °C ).
I = Imax (1 - e 'exponent' -ac), where ingestion maximum (Imax) and the proportionality constant (a) are estimated parameters fitting the curvilinear regression ofv ingestion (I) as a function of initial prey density (c).
marin & al (1986) discuss statistical complications that may arise from presenting ingestion as a function of mean prey density (vs initial prey density); a dependent variable choice which may more accurately reflect a decreased prey density during incubation time, but which also tends to result in comparably higher ingestion rate estimates. In order to provide more conservative estimates of predatory impact, the authors have opted to present our ingestion rates as a function of initial prey density. |
 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.173, Table 2]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.173, Table 2].
Length and chemical composition values for the predator adult female Tortanus dextrilobatus and two prey Oithona davisae and Acartia sp. in the Bay of San Francisco for two experimental temperature. |
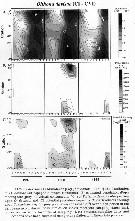 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.174, Fig.5]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.174, Fig.5].
Oithona davisae.
A: Abundance and distribution in the Bay of San Francisco.
B: estimated population feeding rate of Tortanus dextrilobatus preying upon O. davisae
C: potential predatory impact of T. dextrilobatus feeding upon O. davisae prey.
Over 3 yr period.
Stations: see map in fig.1 Bay of San Francisco. |
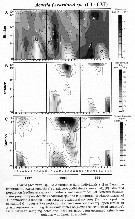 Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.175, Fig.6]. Issued from : R.C. Hooff & S.M. Bollens in Mar. Ecol. Prog. Ser., 2004, 277. [p.175, Fig.6].
Acartia (Acartiura) sp..
A: Abundance and distribution in the San Francisco Bay.
B: estimarted population feeding rate of T. dextrilobatus preying upon Acartia sp prey.
Over a 3 yr period.
Stations: see map in fig.1: Bay of San Francisco. |
| | | | Loc: | | | China Seas (East China Sea, South China Sea, Yellow Sea, Xiamen Harbour, Jiaozhou Bay), Amoy, S & W Korea (Seomjin River estuary, Muan Bay, Asan Bay), Ariake Sea, NE Pacif. (San Francisco Estuary) | | | | N: | 17 | | | | Lg.: | | | (290) F: 2,25-2; M: 2,05-2; (641) F: 1,79-1,74; M: 1,76-1,53; {F: 1,74-2,25; M: 1,53-2,05} | | | | Rem.: | After Ohtsuka & al. (1992, p.119), Chen & Zhang (1965) reported that the specimen from Amoy (type locality) show the 5th pedigerous somite of the female is fused with the 4th and symetrically produced. However, the present specimens of the female have incompletely fused 4th and 5th pedigerous somites and slightly asymmetrical distal corners of the 5th pedigerous somite. In males from the Somjin River, the numbers of ridges on the outer margin of the distal exopodal segment of left P5 and on the inner margin of the exopodal segment of right P5 are 13 and 24-25, respectively, whereas those from Amoy are 9-10 and 19-21, respectively. These may suggest morphological and ecological differences between populations in Amoy and in South Korea.
This species is most closely related to T. derjugini in having the following synapomorphic characters in the sub-genus: 3-segmented urosome in female and elongated, distal segment of right P5 in male. Yhe most distinctive character to distinguish T. dextrilobatus from T. derjugini is the development of the dorsolateral process on the 3rd urosomal somite of the female (the process is considerably developed in T. dextrilobatus but weakly in T. derjugini). The dorsolateral process of female T. derjugini had been overlooked in the previous studies (Brodsky, 1950; Chen & Zhang, 1965). It is difficult to discriminate male in the two species. Chen & Zhang (1965) mentioned that there were several differences between these two males in the shapes of cephalosome, abdomen and P5; however, the reexamination of male T. desjugini from Japanese, Korean and Chinese waters revealed that these differences are not useful enough to distinguish between these two males. There are differences in the structure of right P5: the chela is much smaller in T. dextrilobatus than in T. derjugini; the 2nd segment is larger and relatively wider in T. derjugini than in T. dextrilobatus.
Introduced in San Francisco by ship's ballast waters.
According to Cordell & al., 2008 (p.753) the 1st record of this species in San Francisco Estuary is 1993.
After Hooff & Bollens (2004, p.167), despite a ''dramatic'' increase in the introduction of non-indigenous estuarine zooplankton as T. dextrilobatus in the San Francisco Bay from China in recent decades, the trophic implications of such introduction has been quantified and shows it can play a significant role, at least seasonally or episodically, in the secondary production. However prey size selectivity for all developmental stages of T. dextilobatus remains largely unknown. Additionally
, more thorough understanding of species-specific excape responses and vertical distributions of smaller copepods (such as the seasonally abundant, introduced cyclopoid Limnoithona tetraspina [S.M. Bollens unpubl. data] and larger potential prey items (>850 µm prosome length), will be important aspects for clarifying prey selection behavior by this predator. | | | Last update : 03/03/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 10, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




 dextrilobatus - Plate 1 of morphological figures',750,500);)
 dextrilobatus - Plate 2 of morphological figures',750,500);)
 dextrilobatus - Plate 3 of morphological figures',750,500);)
 dextrilobatus - Plate 4 of morphological figures',750,500);)
 dextrilobatus - Plate 5 of morphological figures',750,500);)
 dextrilobatus - Plate 6 of morphological figures',750,500);)
 dextrilobatus - Plate 7 of morphological figures',750,500);)

 dextrilobatus - Distribution map 2',750,500);)
 dextrilobatus - Distribution map 3',750,500);)
 dextrilobatus - Distribution map 4',750,500);)
 dextrilobatus - Distribution map 5',750,500);)
 dextrilobatus - Distribution map 6',750,500);)
 dextrilobatus - Distribution map 7',750,500);)
 dextrilobatus - Distribution map 8',750,500);)
 dextrilobatus - Distribution map 9',750,500);)
 dextrilobatus - Distribution map 10',750,500);)
 dextrilobatus - Distribution map 11',750,500);)
 dextrilobatus - Plate 1 of morphological figures){kind=link}
 dextrilobatus - Plate 2 of morphological figures){kind=link}
 dextrilobatus - Plate 3 of morphological figures){kind=link}
 dextrilobatus - Plate 4 of morphological figures){kind=link}
 dextrilobatus - Plate 5 of morphological figures){kind=link}
 dextrilobatus - Plate 6 of morphological figures){kind=link}
 dextrilobatus - Plate 7 of morphological figures){kind=link}
 dextrilobatus - Distribution map 2){kind=link}
 dextrilobatus - Distribution map 3){kind=link}
 dextrilobatus - Distribution map 4){kind=link}
 dextrilobatus - Distribution map 5){kind=link}
 dextrilobatus - Distribution map 6){kind=link}
 dextrilobatus - Distribution map 7){kind=link}
 dextrilobatus - Distribution map 8){kind=link}
 dextrilobatus - Distribution map 9){kind=link}
 dextrilobatus - Distribution map 10){kind=link}
 dextrilobatus - Distribution map 11){kind=link}
