|
|
 |
|
Cyclopoida ( Order ) |
|
|
|
Oithonidae ( Family ) |
|
|
|
Oithona ( Genus ) |
|
|
| |
Oithona brevicornis Giesbrecht, 1891 (F,M) | |
| | | | | | | Syn.: | no Oithona brevicornis f. arostrata Früchtl, 1923 (p.454);
? O. brevicornis : Rosendorn, 1917 (p.303); 1917 a (p.34, figs.F);
no O. brevicornis f. aruensis Früchtl, 1923 (p.454);
no O. brevicornis : Wilson, 1932 (p.46); 1932 a (p.315); Deevey, 1948; 1952; ? Deevey, 1960 (tab.II, annual abundance, Rem.: fig.18, 19); Grice, 1956 (p.59); 1960 a (p.487); Anraku, 1964 (p.49); ? Faber, 1966 (p.191, 197, figs.N); ? 1966 a (p.419);
? no O. brevicornis (small form) : Wellershaus, 1969 (p.282, figs.F: 107-108);
O. spinulosa Lindberg, 1947 a (p.129, figs.F); 1950 e (p.17, figs.F); Kos, 1976 (Vol.II, figs. F, Rem.); Shuvalov, 1976 (in Kos, 1976, Vol. II, figs.F, Rem.); 1980 (p.143, figs.F); Ferrari, 1981 (p.1246, figs.F,M); part.
O. brevicornis f. minor Nishida & al., 1977 (p.131, figs. F,M, Rem.);
no O. brevicornis : Uchima, 1979 (p.59, figs. Juv., F,M)
? Oithona brevicornis : Al-Yamani & al., 2011 (p.96, 98, figs.F, Rem.).
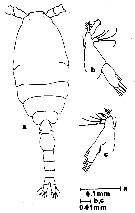
Oithona brevicornis brevicornis : Shiganova & al., 2012 (p.61, fig.3, 4, 5, 6) | | | | Ref.: | | | Giesbrecht, 1892 (p.538, 549, 774, figs.F); Rosendorn, 1917 a (p.34, figs.F,M); Pesta, 1920 (p.549); Sewell, 1924 (p.792, figs.F,M); Früchtl, 1924 b (p.68, Rem.); Kiefer, 1929 g (p.8, Rem.F,M); Rose, 1933 a (p.280, Rem.F,M, figs.F); Sewell, 1933 (p.30); 1934 (p.82); Dakin & Colefax, 1940 (p.116, fig.F); Sewell, 1947 (p.253); Marques, 1949 (p.24, figs.F); 1951 (p.16); Lindberg, 1955 a (p.466: Rem.); Marques, 1958 a (p.134); 1961 (p.46); Kasturirangan, 1963 (p.76, 77, figs.F,M); Marques, 1973 (p.246); Herbst, 1964 (p.155, Rem.); Vilela, 1965 (p.11, figs.F); Gonzalez & Bowman, 1965 (p.271, Rem.); Wellershaus, 1969 (p.279, figs.F,M); 1970 (p.479); Chen & al., 1974 (p.36, figs.F,M, p.74: Rem.); Fonseca & Björnberg, 1975 (1976) (p.130); Bowman, 1975 (Rem.: p.134, figs.F); Shuvalov, 1976 (in Kos, 1976, Vol. II, figs.F, M, Rem.); 1980 (p.136, figs.F,M); Nishida & Ferrari, 1983 (p.72, figs.F,M, Rem.); Ferrari & Orsi, 1984 (p.119); Nishida, 1985 a (p.30, figs.F,M, Rem.: p.141); 1986 a (p.914); Dussart & Defaye,1985 (p.10); Chihara & Murano, 1997 (p.936, Pl. 199, 201: F,M); Yoo & Lim, 1993 (p.91, 93, 98, Table 1, fig.2, Bradford-Grieve & al., 1999 (p.886, 965, figs.F); Al-Yamani & Prusova, 2003 (p.103, figs.F; Rem.: p.105); Vives & Shmeleva, 2010 (p.52, figs.F,M, Rem.; Temnykh & Nishida, 2012 (p.425, fig.2, Table 1, Rem.); Cornils & Heckmann, 2015 (p.243, Fig.2, molecular biology). | 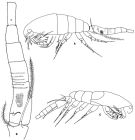 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1245, Fig.1 A-C]. As Oithona spinulosa. Female (from Hong Kong): A, habitus (lateralleft side); B, urosome (lateral left side). Nota: Prosome/Urosome = 1.3. The urosomal segments and furca in the proportional lengths 13:29:15:14:13:16. Genital segment with a distinct patch of long hairs, midlaterally, both sides; beyond 2 rows of small hairs dorsolaterally. Urosomal segment 3 with 3 or 4 rows of small hairs. Caudal ramus: length/width = 3/1. Male: C, habitus (lateral right side). Nota: Prosome/Urosome = 1.6. Rostrum absent. Cephalosome flap extending beyond posterior margin of pediger 1. \\\"Pore signature\\\" of hebes type. Genital flap with 1 thick seta. Caudal ramus: length/width = 2/1
|
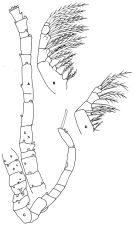 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1247, Fig.2 A-D]. As Oithona spinulosa. Female: A, A1; B, Mx2. Male: C, A1; D, Mx2.
|
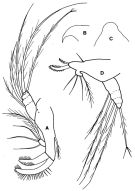 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1248, Fig.3 A-D]. As Oithona spinulosa. Female: A, Md; B, forehead (lateral); C, idem (dorsal). Male: D, Md.
|
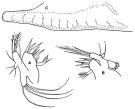 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1249, Fig.4 A-C]. As Oithona spinulosa. Female: A, Mx1. Male: B, Mx1; C, posterior part of cephalosome and flap (\\\"pore signature\\\", right side)..
|
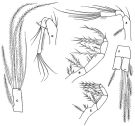 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1250, Fig.5 A-F]. As Oithona spinulosa. Female: A, A2; B, Mxp; C, caudal ramus (dorsal). Male: D, A2; E, Mxp; F, caudal ramus (dorsal).
|
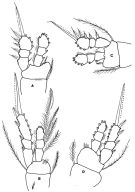 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1251, Fig.6 A-D]. As Oithona spinulosa. Female: A, P1; B, P4. Male: C, P1; D, P4.
|
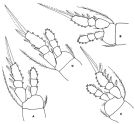 issued from : F.D. Ferrari in Proc. Biol. Soc. Washington, 1981, 94 (4). [p.1252, Fig.7 A-D]. As Oithona spinulosa. Female: A, P2; B, P3. Male: C, P2; D, P3.
|
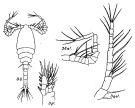 issued from : R.B.S. Sewell in Fauna of the Chilka Lake, 1924, 12. [Pl. XLVI, Fig.3]. Female and Male (from Chika Lake).
|
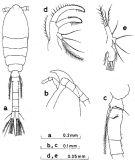 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.31, Fig.11]. As Oithona brevicornis typical form. Female: a, habitus (dorsal); b, forehead (lateral); c, last thoracic segment 5 and genital segment (lateral left side); d, Md (mandinular palp); e, Mx1 (inner lobe not illustrated).
|
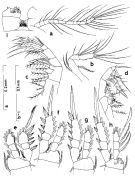 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.32, Fig.12]. As Oithona brevicornis typical form. Female: a, A1; b, A2; c, Mx2; d, Mxp; e, P1; f, P2; g, P3; h, P4; i, connecting plate of P4 (posterior surface). Nota after C.R.: Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 3; 1, 1, 4. P2: 1, 1, 3; 1, 1, 5 P3: 1, 1, 3; 1, 1, 5. P4: 1, 1, 2; 1, 1, 5.
|
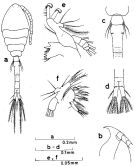 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.34, Fig.13]. As Oithona brevicornis smaller form. Female (from Persian Gulf): a, habitus (dorsal); b, forehead (lateral); c, genital segment (dorsal); d, anal segment and caudal rami (dorsal); e, Md; f, Mx1.
|
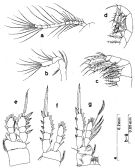 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.35, Fig.14]. As Oithona brevicornis smaller form. Female: a, A1; b, A2; c, Mx2; d, Mxp; e, P1; f, P2; g, P4
|
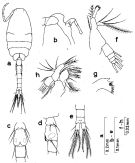 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.36, Fig.15]. As Oithona brevicornis smaller form. Male: a, habitus (dorsal); b, forehead (lateral); c, genital segment (ventral); d, idem (lateral left side); e, last urosomal segments and caudal rami (dorsal); f, Md (mandibular palp; g, Md (biting edge); h, Mx1.
|
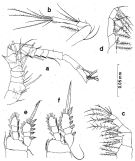 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.37, Fig.16].. As Oithona brevicornis smaller form. Male: a, A1; b, A2; c, Mx2; d, Mxp; e, P1; f, P2. Nota after C.R.: Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 3; 1, 1, 4. P2: 1, 1, 3; 1, 1, 5 After Vives & Shmeleva (2010, p.53): P3: 1, 1, 3; 1, 1, 5. P4: 1, 1, 2; 1, 1, 5.
|
 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.4, Figs.1-8]. Female (from China Seas): 1, habitus (dorsal); 2, forehead (lateral); 3, P1; 4, P2; 5, P3; 6, P4; 7, habitus (another specimen). Male: 8, habitus (dorsal)
|
 issued from : F.Y. Al-Yamani & I. Prusova in Common Copepods Northwestern Arabian Gulf : Identification Guide. Kuwait Institute for Scientific Research, 2003. [p.104, Fig.38]. With doubt. Female: A, habitus (dorsal); B, forehead (lateral); C, A2; D, Md (mandibular palp); E, P1; F, P2; G, P3; H, P4.
|
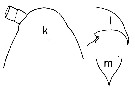 issued from : T.E. Bowman in Chesapeake Sc., 1975, 16 (1). [p.135, Fig.1, k-m]. Female (from Hong Kong): k, forehead (dorsal); l, idem (lateral); m, rostrum (ventral); Nota: O. brevicornis, to which O. colcarva has been assigned erroneously, has among other differences, a much longer and narrower rostrum and 4 setae on the endopod of Md.
|
 issued from : K. Lindberg in Rec. Indian Mus., 1947, 45. [p.130, Fig.1]. As Oithona spinulosa. Female (from coast of Madras, India): a, forehead (lateral); b, thoracic segment 5 and abdominal segments 1 and 2 (ventral); c, Md. Nota: Formula of spines on the outer edge from P1 to P4: 1--1-3; 1-1-3; 1-1-3; 1-1-2, respectively. Nota: For the author, this species is close to O. brevicornis but differs by the genital segment, the length and structure of caudal setae, and the swimming legs and P5.
|
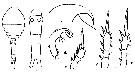 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. \\\"Valdiviella\\\", 1917, 23. [p.34, Fig.19]. Female: a, habitus (dorsal); b, urosome; c, forehead; d, Md; e, exopod of P1; f, exopod of P2; g, exopod of P4. Nota: Proportion of lengths (p.cent) Prosome : 53.85, Urosome : 46.15 . Relative lengths of urosomal segments and caudal rami: 10 : 30 : 14 : 13 : 11 : 13. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae ; Si = inner setae), P1 : 1, 1, 3 Se ; 0, 1, 4 Si ; P2 : 1, 1, 3 Se ; 0, 1, 5 Si ; P3 : 1, 1, 3 Se ; 0, 1, 5 Si ; P4 : 1, 1, 2 Se ; 0, 1, 5 Si .
|
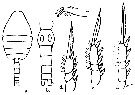 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. \\\"Valdiviella\\\", 1917, 23. [p.35, Fig.20]. Male: a, habitus (dorsal); b, urosome; c, Md; d, P2; e, P3; f, P4. Nota: Proportion of lengths (p.cent) Prosome : 63.83, Urosome : 36.17 . Relative lengths of urosomal segments and caudal rami8 : 9 : 30 : 15 : 14 : 12 : 12. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae ; Si = inner setae), P1 : 1, 1, 3 Se ; 0, 1, 4 Si ; P2 : 1, 1, 3 Se ; 0, 1, 5 Si ; P3 : 1, 1, 3 Se ; 0, 1, 5 Si ; P4 : 1, 1, 2 Se ; 0, 1, 5 Si .
|
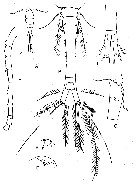 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.144, Fig.39]. As Oithona spinulosa. After Lindberg, 1960. Female: A, habitus (dorsal); B, urosome (dorsal); C, posterior part of urosome and caudal rami (dorsal); D, rostrum (lateral); E, same (frontal view; F, A1; G, 5th thoracic segment with P5 and genital segment (lateral).
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.145, Fig.40]. As Oithona spinulosa. After Lindberg, 1960. Female: A, A2; B, Md; C, P1; D, P4. Setal formulae on exopod of P1 and P4 do not agree with Nishida (1985).
|
 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.137, Fig.35]. As Oithona brevicornis. Female: A-B, habitus (lateral and dorsal, respectively); C-D, cephalon (dorsal and lateral, respectively); E-F, rostrum (lateral, right and left side respectively); G, Md; H, P1; I, P3; J, P4; k, 4 th and 5th thoracic segment, genital segment and 2nd urosomal segment (dorsal); L, posterior part of urosome. Male: M, habitus (habitus (dorsal); N, forehead (lateral).
|
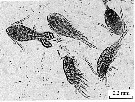 issued from : Zh.P. Selifonova in inInland Water Biol., 2009, 1. [p;31, Fig.2]. Samples collected in the Novorossiysk Harbor (NE Black Sea).
|
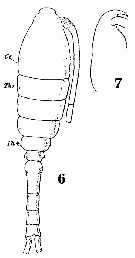 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 34, Figs.6, 7]. Female: 6, habitus (dorsal); 7, forehead (lateral).
|
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.130, Fig.4]. Female (from Suruga Bay and adjacent waters) forma minor, doubtful: a, habitus (dorsal), b, forehead (lateral). forma typica: c, caudal rami; d, Md; e, P1; f, P2; g, P3; h, P4. Nota: Setae formula of exopod P1 to P4 (outer setae in first, inner setae in second). P1: 1,1,3; 0,1,4. P2: 1,1,3; 1,1,5. P3: 1,1,3; 1,1,5. P4: 1,1,2; 1,1,5. Setae of P5 and caudal rami with abundant hair. Prosome length/urosome length = 1.16-1.37 (forma typica); 1.11-1.38 (forma minor
|
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.131, Fig.5]. Male (from Suruga Bay and adjacent waters) forma minor, doubtful: a, habitus (dorsal), c, Md. Male forma typicatypica: b, Md. Nota: Setae formula of exopod P1 to P4 (outer setae in first, inner setae in second).P1: 1,1,3; 1,1,4. P2: 1,1,3; 1,1,5. P3: 1,1,3; 1,1,5. P4: 1,1,2; 1,1,5. Prosome length/urosome length = 1.43-1.68 (forma typica; 1.50-1.79 (forma minor)
|
 Issued from : A. Temnykh & S. Nishida in Aquatic Inv., 2012, 7 (3). [p.427, Fig.2]. Character comparison between Oithona brevicornis (A-D), O. aruensis (E-G), and O. davisae (H-K). A, E, H: rostrum; B, F, I: Md; C, G, J: Mx1, indicating distal-most spine (thin arrow) of 1st inner lobe, endopod (arrowhead), seta on 2nd inner lobe (dotted arrow, absent in J) and seta on 2nd basal segment (thick arrow, absent in J); D: genital double-somite and next somite, lateral view, indicating rows of hairs (arrows) in O. brevicornis; K: genital double-somite, lateral view. Not at scale. After Nishida (1985: A-D), Nishida & Ferrari (1983: E-G) and Ferrari & Orsi (1984: H-K).
|
 Issued from : A. Temnykh & S. Nishida in Aquatic Inv., 2012, 7 (3). [p.428, Table 1]. Characteristics in O. brevicornis adult female. Nota: Compare with the species O. wellershausi, O. aruensis, O. davisae, and O. davisae from the Black Sea.
|
 Issued from K.-I. & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2). [Key of female]: ]Morphological characters of Oithona brevicornis female in Korean waters :
1 - Anterior part of prosome rounded in dorsal view.
2 - Rostrum pointed ventrally. 3 - Length of A1 shorter than that of prosome. 4 - Length of 1st spine in inner lobe 1 of Mx1 same as that of other spines.
|
 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.280, Fig.103-119]. Female (from Cochin Backwater, S. India): 103, habitus (dorsal; A1 segmented not correct); 106, urosome (dorsal); ciliation on all setae yellow); 109, forehead (lateral); 111, P4; 112, P1; 113-115, Md (different aspects); 119, A2. Male: 104, forehead (from the side andventral); 110, A1 (n = notch); 116, Md; 118, A2.
Scale : single line = 0.1 mm ; double line = 0.05 mm. Nota Female (great form): - Anal segment broader than long. - Caudal rami: length : broadth = 2.5 : 1. The insertion of Se divides the lateral margin into a proximal and a distal portion, the relations of which are 2 : 7; Si ciliatedon the outer side; Si naked, length= abdominal segments 3-5 + caudal rami. - P5 with 1 naked basal seta on a papilla, it is bent and shows upwards; there is1 terminal thick seta, the latter reaches the posterior margin of abdominal segments 1-2 and is ciliated; - Se, all St and the terminal seta of P5 are ciliated with strong yellow cilia. - A1 reaches the posterir margin of yjoracic segment 3 or 2. - Rostrum acute, bent, downwards. - Genital segment with a yuft of tender hairs on the sides just behind the genital swelling. - Small spinules on the sides of abdominal segment1-2 and 3 absent (or only very weak in some specimens) In some specimens abdomonal segment 5 has fine spinules on the ventral side, and sometimes also on the dorsal aspect of the posterior margin) Abdominal 1-2 - 4 are naked. - Md has 2 conspicuous hooks at the end of basis; the median hook is longer and bears on the median side a row of about 10 strong spines; both hooks bear at the end on each side a row of fine cilia, as a whole resembling two membranes; the hooks ebd in a short spine each; endopod bears 4 hairs of which the proximal is stronger and finely ciliated, on the opposite side there is a ciliated seta. Nota Male: - Ratio prosome : urosome = 1.56. - Forehead slightly bent downwards; withoit a rostral horn. Ratio prosome (cephalon-4th thoracic segment) : urosome (5th thoracic segment-caudal rami) = 1.56. - Md has a somewhat different shape: the median hook of basis is shorter, almost straight, with spines on both sides; the lateral hook resembles that of the female, only, there is an additional spine on either side. - Abdominal segment 5 (anal) has fine spinules on the ventral side of the posterior margin. - In some males the setae of the caudal rami have the same yellow ciliation as in the female.
|
 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.281]. Female I (great form): Armature of the swimming legs P1 to P4 . Nota: Proportion of the exopod of P4 terminal spine : whole exopod = 8 : 9. Endopod segment 3 of P4 bears a strong seta on the inner margin in the most proximal position (only seen in the mature female). Older copepodite have a thick and yellow ciliated seta on P6, like the terminal seta on P5. - All copepodites bear a typical rostrum as the females.
|
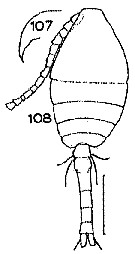 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.280, Fig. 107, 108]. With doubt. Female (small form): 107, rostrum (lateral); 108, habitus (dorsal)/ Scale bar = 100 µm. Nota: - Anal segment broader than long. - Caudal rami: length : breadth = 2 : 1; proximal : distal portion of the lateral margin = 2 : 3; length of the external seta (Se) = 1.3 x caudal rami; length of the inner seta (Si) = abdominal segments 3-5 + caudal rami. Caudal setae with fine, colourless and transparent cilia. - P5 as in the species type. - A1 reaches the posterior margin of the 2nd thoracic segment. - Rostrum acute, bent downwards. - Genital segment without hairs on the sides. - Small spinules absent on the abdomen, all abdominal segments - Md similar to samples from Cochin Backwaters (great form), however, measures 1 naked./3 in length. - Exopodal segment 3 of P4 bears 1 strong seta similar to samples from Cochin Backwater (great form). - Both forms occurred in water with more than 22 p.1000 salinity, although sometimes found in a 4 p.1000 sample. The small form closely resembles O. brevicornis forma aruensis Früchtl, 1924 from Aru Islands (Indonesia) in brackish water. The only difference is the shape of the terminal hooks of Md; they are sharply pointed in the forma aruensis, whereas in the small Cochin animals they resemble those of the great form.
Both the small and the great form occurred in the same samples in the Cochin Backwater, but intermediate forms were also seen.
|
 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.279]. Female (great form): Body length, ratio prosome : urosome and ratio of the urosomal segments.
|
 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.282]. Female (small form): Body length, ratio prosome : urosome and ratio of the urosomal segments.
|
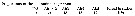 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.283]. Female (from Hong Kong Harbour): Body length, ratio prosome : urosome and ratio of the urosomal segments. Nota: Body length: 0.66; 0.67; 0.70 mm. - Anal segment broader than long. - Caudal rami: length : breadth = 3.3 : 1; proximal : distal portion = 2 : 6. External seta (Se)longer than caudal rami; inner seta (Si) naked, length as in the Cochin Backwater typical form; in some females the setae are yellow as in Cochin animals. - A1 reaches thoracic segment 2 or 3. - Rostrum acute, bent downwards, however, in general, there seems to be a variation in the shape of the rostrum, so that it cannot used for differentiation between the forms. - Tender hairs on the genital segment. - Small spinules on the side of abdominal segment s1-2 and 3 were never seen.. No spinules seen on Anal segment. - Endopodal segment 3 of P4 bears a similar strong seta as in the Cochin animal (fig.111). - Md as for the Cochin animals Male: Md as figured for the Cochin animal (fig.116).
|
 Issued from : S. Wellershaus in Veröff. Inst. Meeresforsch. Bremerh., 1969, 11 (2). [p.283, Table 4]. Comparative list of characters in Oithona brevicornis female (in addition to Herbst, 1964).
|
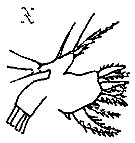 Issued from : K.-I. Yoo & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2) [p.99, Fig.2, N] Female (from Korean Waters): N, Mx1.
|
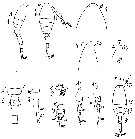 Issued from : M.S. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. II, 1976. original from V.S. Shuvalov, 1976. Female: 1-2, habitus (lateral and dorsal, respectively); 3-4, forehead (dorsal and lateral view, respectively); 6, posterior thoracic and genital segments (dorsal); 7, posterior abdominal segments (dorsal); 8, Md; 9, P1; 10, P3; 11, P4. Male: 5, forehead (lateral view); 12, habitus (dorsal). Spine formulae: P1: 1-1-3; P2: 1-1-3; P3: 1-1-II3; P4: 1-1-2.
| | | | | Compl. Ref.: | | | Carazzi & Grandori, 1912 (p.15, 38); ? Wilson, 1942 a (p.196); Fagetti, 1962 (p.38); Gaudy, 1963 (p.30, Rem.); Mazza, 1966 (p.73); Pavlova, 1966 (p.44: ?); Ehrhardt, 1967 (p.742, geographic distribution, Rem.); Delalo, 1968 (p.138); Heinle, 1969 (p.186, fig.4, 5, Table 1, 2, 4); Palet, 1975 (p.660); Kos, 1976 (Vol.II, figs.F;M, Rem.); Madhu Pratap & al., 1977 (p.138, Table 3: abundance vs. stations); Hirota, 1979 a (p.33, Table 2, fig.5); Chen Q-c, 1980 (p.795); Vaissière & Séguin, 1980 (p.23, tab.2); Hirota 1981 (p.19, Table 1, length-weight-CHN); Uye S-i., 1982 (p.149, relation length-weight-C-N); Vives, 1982 (p.295); Kovalev & Shmeleva, 1982 (p.85); Dussart, 1984 (p.25, 64: occurrence): Lozano Soldevilla & al., 1988 (p.61); Yoo, 1991 (tab.1); Dai & al., 1991 (tab.1); Kim & al., 1993 (p.271); Godhantaraman, 1994 (tab.5, 6); Böttger-Schnack, 1995 (p.92); Shih & Young, 1995 (p.74); Böttger-Schnack, 1996 (p.1088); Park & Choi, 1997 (Appendix); Noda & al., 1998 (p.55, Table 3, occurrence); Hure & Krsinic, 1998 (p.103); Suarez-Morales & Gasca, 1998 a (p.111); Ueda & al., 2000 (tab.1); Seridji & Hafferssas, 2000 (tab.1); Hsieh & al., 2004 (p.398, tab.1); Rezai & al., 2004 (p.486, tab.2, 3, abundance); Lo & al., 2004 (p.468, tab.2); Zuenko & Nadtochii, 2004 (p.526, tab.1, fig.8); Hwang & al., 2005 (p.106); non Gubanova & Altukhov, 2007 (p.407); Madhu & al., 2007 (p.54, Table 4, abundance vs monsoon); Khelifi-Touhami & al., 2007 (p.327, Table 1); Fernandes, 2008 (p.465, Tabl.2); non Selifonova & al., 2008 (p.305, Tabl. 2); Shmeleva & al., 2008 (p.31, Table 1); Ohtsuka & al., 2008 (p.115, Table 4, 5, Rem.: in Japan Harbors); non Selifovona, 2009 (p.30); Hwang & al., 2010 (p.220, Table 2, fig.3, 4); Mazzocchi & Di Capua, 2010 (p.428); Fazeli & al., 2010 (p.153, Table 1); non Selifonova, 2011 (p.227, Rem: anthropic effects); Shanthi & Ramanibai, 2011 (p.132, Table 1); Chew & Chong, 2011 (p.127, Table 3, abundance vs location); Zhang G.-T. & Wong, 2011 (p.277, fig.6, 7, 8, abundance, indicator); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea, Rem.: p.78); Beltrao & al., 2011 (p.47, Table 1, density vs time); DiBacco & al., 2012 (p.483, Table S1, as O. brevicornis brevicornis, ballast water transport); Etilé & al., 2012 (p.627, spatio-temporal distribution, fecundity, production); Salah S. & al., 2012 (p.155, Tableau 1); Gubanova & al., 2013 (in press, Rem.: p.4); Mihneva & Stefanova, 2013 (Rem.: p.120); Zamora Terol, 2013 (p.174: Table 5); Jagadeesan & al., 2013 (p.27, Table 3, seasonal variation); Lidvanov & al., 2013 (p.290, Table 2, % composition) ; Pansera & al., 2014 (p.221, Table 2, fig.4, annual abundance) ; Zaafa & al., 2014 (p.67, Table I, occurrence); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Ohtsuka & Nishida, 2017 (p.565, Table 22.1); El Arraj & al., 2017 (p.272, table 2, spatial distribution); | | | | NZ: | 11 | | |
|
Distribution map of Oithona brevicornis by geographical zones
|
| | | | | | | | | | | |  Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.141, Fig.92]. Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.141, Fig.92].
Indo-Pacific geographical distribution of Oithona brevicornis. |
| | | | Loc: | | | Angola, Congo (mouth), San Tomé and Principe Is., Ivory Coast (Grand-Lahou lagoon); Guinea , Bahia de Mochima (Venezuela), ? G. of Mexico, ? Delaware Bay, Canary Is., off Morocco-Mazuritania, Cap Ghir, Portugal, off W Tangier, Medit. (M'Diq, Gulf of Annaba, Ligurian Sea, Tyrrhenian Sea, Lake Faro (Sicily), Adriatic Sea, Venice, Ionian Sea, Aegean Sea, W Lebanon Basin), [non Black Sea, Sebastopol Bay, Novossiysk & Tuapse harbors)], Suez Canal, Red Sea, Gulf of Oman, Arabian Sea, Arabian Gulf, ? Kuwait, Agatti Is (offside & very more abundant in lagoon), India (Saurashtra coast, Kerala, S, Coom & Adyar, Madras, Gulf of Mannar, Palk Bay, Chilka Lake, Calcutta Salt Lakes), Bay of Bengal, Malaysia (Kurau River), Verlaten Is., Straits of Malacca, Sangga estuary (mainly nearshore and offshore), G. of Thailand, Sunda Straits, China Seas (Bohai Sea, Yellow Sea, East China Sea, South China Sea, Hong Kong, Xiamen Harbour), Taiwan (W, Tapong Bay), Korea (E, S & W), Japan Sea, Japan ( Kuchinoerabu Is., Seto Inland Sea, Setonaikai, Ariakae Bay, Kyushu: Shijiki Bay), E Siberia, S Kuril Is., off Chile | | | | N: | 99 | | | | Lg.: | | | (46) F: 0,7; (104) F: 0,79; (109) F: 0,62-0,55; M: 0,52-0,45; (172) F: 0,58-1,70 ?; M: 0,65; (280) F: 0,741-0,646; (334) F: 0,6; M: 0,55; (336) F: 0,62-0,57; M: 0,57-0,49; (373) F: 0,67-0,48; M: 0,63-0,46; (433) F: 0,58-0,51; M: 0,53-0,43; (449) F: 0,7; (627) F: 0,62-0,58; 0,5-0,48; M: 0,47-0,46; (630) F: 0,63-0,58; M: 0,57-0,52; (631) F: 0,51; 0,48; M: 0,46; 0,42; (649) F: 0,52; M: 0,47; (761) F: 0,741-0,646; (798) F: 0,57-0,64; M: 0,53-0,57; (880) F: 0,65-0,74; 0,48-0,70; M: 0,40-0,47; (937) F: 0,53-0,58; (1006) F: 0,646-0,741; ? (1085) F: 0,5-0,75; (1239) F: 0,64-0,73; (1232): F: 0,48-0,7; M: 0,40-0,47; {F: 0,48-0,79; M: 0,40-0,65} | | | | Rem.: | epipelagic; marine & brackish.
For Nishida (1985, p.38) the species is closely allied O. aruensis and O. wellershausi.
[According to Gubanova & Altukhov (2007, p.409) this species is probably brought into Sevastopol Bay in the ballast water of a ship, but this species corresponds to Oithona davisae].
For Wellershaus (1969, p.284) the specimens described by Lindberg do not show striking differences; at the most spinulosa can be considered as a sub-species based on the lateral spinules in abdominal segments 1-2 and 3. See Yoo & Lim (1993, p.94) in remarks concerning morphological characters between O. brevicornis and O. davisae.
For Al-Yamani & Prusova (2003, p.105) the specimens agree with O. brevicornis Giesbrecht, 1891, but differ from the latter in proportions of prosome length and width, and in caudal ramus lateral seta being plumose and extending beyond posterior margin of caudal ramus. In structure of lateral seta the present specimens correspond to O. spinulosa Lindberg,1950. However Nishida (1985) considers O. spinulosa Lindberg,1950 as a junior of O. brevicornis Giesbrecht,1891. Shuvalov (1980) believed that O. brevicornis s.l. could be a complex of closely related species.
After Temnykh & Nishida (2012, p.429) O. brvicornis that has been reported to be recently introduced into the Black Sea is indeed O. davisae. For Gubanova & al. (2013, p.5)O. brevicornis has been routinely observed in the Black Sea since the mid 2000s but recently, the species was reidentified as O. davisae. | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 28, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
