|
|
 |
|
Cyclopoida ( Order ) |
|
|
|
Oithonidae ( Family ) |
|
|
|
Oithona ( Genus ) |
|
|
| |
Oithona simplex Farran, 1913 (F,M) | |
| | | | | | | Ref.: | | | Farran, 1913 a (p.187, figs.F); Rosendorn, 1917 (p.303); 1917 a (p.44, Rem.F,M, figs.M); Früchtl, 1924 b (p.73, Rem.); Gurney, 1927 (p.160, Rem.); Kiefer, 1929 g (p.9, Rem.F,M); Sewell, 1948 (p.461); Lindberg, 1955 a (p.466: Rem.); Marques, 1957 (p.41); Tanaka, 1960 (p.64, figs.F,M); Grice, 1960 a (p.488, figs.12-18); Rem); Herbst, 1964 (p.157, Rem.); Gonzalez & Bowman, 1965 (p.274, figs.F, Rem.); Wellershaus, 1970 (p.477, 478); Björnberg, 1972 (p.83, figs., Rem.N); Kos, 1972 (Vol.I, figs. F, Rem.); Chen & al., 1974 (p.34, figs.F,M, p.74: Rem.); Nishida & al., 1977 (p.151, figs. F,M, Rem.); Shuvalov, 1980 (p.162, figs.F, Rem.M); Ferrari & Bowman, 1980 (p.19, figs.F,M, Rem.); Sazhina, 1985 (p.92, figs.N); Nishida, 1985 a (p.88, Redescr.F, figs.F, Rem.F,M, 2 forms, p.140); Nishida, 1986 (p.386); Valentin & al., 1987 (p.1202); Yoo & Lim, 1993 (p.91, 97, 100, Table 1); Gaughan & Potter, 1995 (p.121); Chihara & Murano, 1997 (p.939, Pl.197,201: F,M); Bradford-Grieve & al., 1999 (p.886, 967, figs.F); McKinnon, 2000 (p.108, Rem.); Conway & al., 2003 (p.205, figs.F,M, Rem.); Vives & Shmeleva, 2010 (p.78, figs.F,M, Rem.); Cornils & Heckmann, 2015 (p.243, Fig.2, molecular biology). | 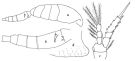 issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.20, Fig.12]. Female (from Caribbean area): a, Habitus (lateral right side); b, urosome (lateralright side); c, P4.Br>Nota: Prosome /Urosome = 1.6. Endopodal segments 2 and 3 of P4 with internal setae without curvature of flange. Knob near genital opening with small, thin spine medially, a smaller spine ventrally, and a tiny spine dorsally. Male: d, posterior part of cephalosome with \"pore signature\". Nota: Prosome/Urosome = 1.5. Cephalosome similar to female, without flap.
|
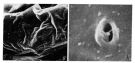 issued from : F.D. Ferrari & T.E. Bowman in Smiths. Contr. Zool., 1980, 312. [p.24, Fig.15 g-h]. Male (SEM): g, \"pore signature\" (right side, X1800); h, single pore. Nota: Individual pores without well-developed ridges; integument slightly thickened anteriorly and posteriorly.
|
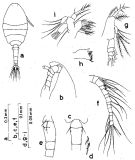 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.92, Fig.52]. As Oithona simplex typical form. Female: a, habitus (dorsal); b, forehead (lateral); c, thoracic segment 5 and genital segment (dorsal); d, P6; e, thoracic segment 5 and genital segment (lateral right side); f, A1; g, Md (mandibular palp); h, Md (biting edge); i, Mx1. Nota: Proportional lengths of urosome segments and caudal ramus 12:36:16:16:8:11.
|
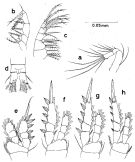 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.93, Fig.53]. As Oithona simplex typical form. Female: a, A2; b, Mxp; c, Mx2; d, anal segment and caudal rami (dorsal); e, P1; f, P2; g, P3; h, P4. Nota after Vives & Shmeleva (2010, p.79): Setal formula on outer margin (in first) and inner margin (in second) of exopod segments (from proximal to distal) of P1 to P4: P1: 1, 1, 3; 1, 1, 4. P2: 1, 1, 3; 1, 1, 5 P3: 1, 1, 3; 1, 1, 5. P4: 1, 1, 3; 1, 1, 5.
|
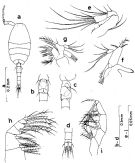 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.94, Fig.54]. As Oithona simplex typical form. Male: a, habitus (dorsal); b, thoracic segment 5 and genital segment (ventral); c, idem (lateral left side); d, last urosomal segments and caudal rami (dorsal); e, A2; f, Md; g, Mx1; h, Mx2; i, Mxp.
|
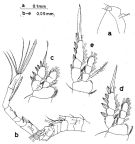 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.95, Fig.55]. As Oithona simplex typical form. Male: a, forehead; b, A1; c, P1; d, P2; e, P4.
|
 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.97, Fig.56]. As Oithona simplex long form. Female: a, habitus (dorsal); b, forehead); c, anal segment and caudal rami (dorsal); d, idem (ventral); e, A2; f, Md (mandibular palp); g, Md (biting edge); h, Mx1. Nota: Proportional lengths of urosome segments and caudal ramus 11:33:15:16:11:13.
|
 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.98, Fig.57]. As Oithona simplex long form. Female: a, A1; b, Mx2; c, Mxp; d, P1; e, P2; f, P3; g, P4. Nota: Mxp as in typical form but endopodal segment with 1 more minute seta. 2
|
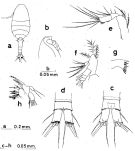
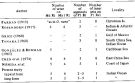 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.96, Table 4]. Structure of Md, Mx1 and P1 of female Oithona simplex reported by workers. Ri = endopodite; Re = exopodite.
|
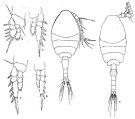 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.3, Figs.4-9]. Female (from China Seas): 4, habitus (dorsal); 5, P1; 6, P2; 7, P3; 8, P4. Male: 9, habitus (dorsal).
|
 issued from : G.P. Farran in Proc. zool. Soc. Lond., 1913. [Plate XXIX, Figs.10-14]. Female (from Christmas Island, Indian): 10, forehead (lateral); 11, habitus (dorsal); 12, urosome (dorsal); 13, P3; 14, P4.
|
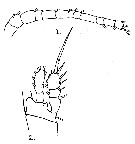 issued from : G.P. Farran in Proc. zool. Soc. Lond., 1913. [Plate XXX, Figs.1-2]. Female: 1, A1; 2, P1.
|
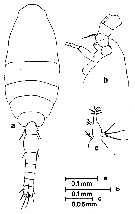
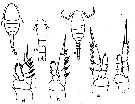 issued from : G.D. Grice in Bull. mar. Sci. Gulf Caribb., 1960, 10 (4). [p.486, Figs.12-18]. Female (from NE Gulf of Mexico: off Alligator Harbor): 12, habitus (dorsal); 13, urosome (dorsal); 14, P1; 15, P2; 16, P3; 17, P4. Nota: Rostrum absent. Exopod segments 1-3 of P4 with 1, 1, 3 spines, respectively. Male: 18, habitus (dorsal). Nota: Exopod segments 1-3 of P1 with 1, 1, 3 spines, respectively. Exopod segments 1-3 of P4 with 1, 1, 3 spines, respectively. Remarks: The specimens of the Gulf of Mexico diverge slightly from those described by Farran (1913) in two respects. The 1st segment of the endopod of P1 has 1 seta and the 2nd segment of the exopod of P4 has 1 inner seta. These segments in Farran have 0 and 2 setae, respectively.. Rosendorn (1917) stated that the endopod of P1 consists of 3 segments and not 2 as shown by Farran. There is also some confusion regarding the number of setae on the 3rd endopodal segment of P4; Rosendorn apparently misinterpreted Farran's method of presenting the number and position of the setae on the various endopodal segments of the swimming legs.
|
 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. \"Valdiviella\", 1917, 23. [p.45, Fig.26]. Male: a, habitus (dorsal); b, urosome (dorsal); c, Md; d, Mx1; e, exopod of P1; f, exopod of P2. Nota: Proportion of lengths (p.cent) Prosome : 62.16, Urosome : 37.84 . Relative lengths of urosomal segments and caudal rami: 6 : 9 : 5 : 5.5 : 5 : 2.5 : 4. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae), P1 : 1, 1, 3 Se ; P2 : 1, 1, 3 Se; P3 : 1, 1, 3 Se ; P4 : 1, 1, 3 Se. Female: Proportion of lengths (p.cent) Prosome : 65.85, Urosome : 34.15 . Relative lengths of urosomal segments and caudal rami: 6 : 20 : 9 : 10 : 4.5 : 9. Setal formula of the exopod swimming legs P1 to P4 (Se = outer setae), P1 : 1, 1, 3 Se; P2 : 1, 1, 3 Se; P3 : 1, 1, 3 Se; P4 : 1, 1, 3 Se.
|
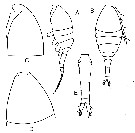 issued from : V.S. Shuvalov in Opred. Faune SSSR, Nauka, Leningrad, 1980, 125. [p.157, Fig.45]. Female: A-B, habitus (lateral and dorsal, respectively); C-D, cephalon (lateral and dorsal, respectively); E, urosome.
|
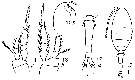 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. \"Valdiviella\", 1917, 23. [p.192, Fig.64, 10-14]. Female: 10, forehead (lateral); 11, habitus (dorsal); 12, urosome (dorsal); 13, P3; 14, P4.
|
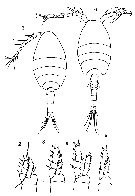
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.152, Fig.23, a-h]. female (off Sagami Bay) and adjacent waters: a, habitus (dorsal); b, forehead (lateral); c, Md; d, Mx1; e, P1; f, P2; g, P3; h, P4. Nota: Formulae of exopod P1 to P4 (outer setae in first, inner setae in second). P1: 1,1,3; 1,1,4. P2: 1,1,3; 1,1,5. P3: 1,1,3; 1,1,5. P4: 1,1,3; 1,1,5; Prosome length/urosome length = 1.82
|
 Issued from : S. Nishida, O. Tanaka & M. Omori in Bull. Plankton Soc. Japan, 1977, 24 (2). [p.152, Fig.23, i-n]. Male (off Sagami Suruga Bay): i, habitus (dorsal); j, forehead (lateral); k, P1; l, P2; m, P3; n, P4. Nota: Prosome length/urosome length = 2.00. P1-P4 armed as in female.
|
 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [Pl. XXVIII, 1-6]. Female (from South China Sea and Indian Ocean): 1, habitus (dorsal); 2, P1; 3, P2; 4, P3; 5, P4. Nota: Prosome and urosome in the proportional lengths 65 to 35. Rostrum absent. Urosomal segments and caudal rami in the proportional lengths 12 : 35 : 15 : 15 : 8 = 100. Caudal rami divergent, 2 times as long as broad. Male: 6, habitus (dorsal). Nota: Prosome and urosome in the proportional lengths 64 to 44. Urosomal segments and caudal rami in proportional lengths 16 : 22 : 17 : 15 : 15 : 5 : 10 = 100. The A1 has the knee-joint between the segments 11 and 12.
|
 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [p. 65]. Female: Proportional lengths of segments of A1. Nota: A1 short, extends to the level of the posterior end of the 2nd thoracic segment.
|
 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [p. 65]. Female: Number of outer marginal spines and inner marginal setae of the exopodal segments of the swimming legs P1 to P4.
|
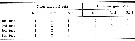 Issued from : O. Tanaka in Spec. Publs. Seto mar. biol. Lab., 10, 1960 [p. 65]. Female: Number of the inner and outer marginal setae on the endopodal segments of the swimming legs P1 to P4.
|
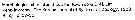 Issued from : K.-I. Yoo & D.-H. Lim in The Korean J. Syst. Zool., 1993, 9 (2) [p.97: key of female] in Korean Waters: Morphological characters of Oithona smplex female :
1 - Anterior part of prosome rounded in dorsal view.
2 - Rostrum blunt. 3 - P5 with 1 seta. 4 - Length of A1 less than 0.7 times of that of prosome.
| | | | | Compl. Ref.: | | | V.N. Greze, 1963 a (tabl.2); De Decker & Mombeck, 1964 (p.13); Shmeleva, 1964 a (p.1068); 1965 b (p.1350, lengths-volume -weight relation); Pavlova, 1966 (p.44); Delalo, 1968 (p.138, as simplix); Newbury & Bartholomew, 1976 (p.373, production); Ferrari, 1977 (p.410); Dessier, 1979 (p.134, 201, 207); Kovalev & Shmeleva, 1982 (p.85); Dussart, 1984 (p.25, 64: occurrence): Greze & al., 1985 (p.8); Nishida, 1985 (p.125); Turner, 1986 (p.289, fig. 7: pellet-content): Böttger-Schnack al., 1989 (p.1089); Othman & al., 1990 (p.561, 564, Table 1); Yoo, 1991 (tab.1); Dai & al., 1991 (tab.1); McKinnon, 1991 (p.471); Böttger-Schnack, 1992 (p.304); Lopes, 1994 (tab.1); Suarez Morales, 1994 b (tab.1); Gaughan & Potter, 1995 (tab.1); Shih & Young, 1995 (p.75); Gouda & Panigrahy, 1995 (p.206); Böttger-Schnack, 1995 (p.92); 1996 (p.1088); McKinnon & Ayukai, 1996 (p.595); Hopcroft & Roff, 1996 (p.789, diel egg production); Böttger-Schnack, 1997 (p.410); Noda & al., 1998 (p.55, Table 3, occurrence); Hure & Krsinic, 1998 (p.103); Hopcroft & al., 1998 (tab.2); Lopes & al., 1998 (p.195); Suarez-Morales & Gasca, 1998 a (p.112); Razouls & al., 2000 (p.343, Appendix); Ueda & al., 2000 (tab.1); Ueda, 2001 (p.96, occurrence); McKinnon & al., 2003 (p.105, tab.2, fig.11); Hsieh & al., 2004 (p.398, tab.1); Rezai & al., 2004 (p.486, tab.2, 3, abundance, Rem., p.495, tab.8); Lo & al., 2004 (p.468, tab.2); Zuo & al., 2006 (p.159, tab.1, 3, fig.9, spatial abundance, fig.8: stations group); Porri & al., (p.714); Selifonova & al., 2008 (p.305, Tabl. 2); Ohtsuka & al., 2008 (p.115, Table 4, 5, Rem.); Osaka and Kitakyushu Harbors); McKinnon & al., 2008 (p.843: Tab.1, p.846: Tab. II, fig.7); Miyashita & al., 2009 (p.815, Tabl. II); Fazeli & al., 2010 (p.153, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I); Zhang G.-T. & Wong, 2011 (p.277, fig.6, 7, 8, abundance, indicator); Saitoh & al., 2011 (p.85, Table 4, 5); Beltrao & al., 2011 (p.47, Table 1, density vs. time); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Ali & al., 2011 (p.601, fig.11, abundance vs tidal); Chew & Chong, 2011 (p.127, fig.4, 5, Table 2, 3, abundance vs location); Uysal & Shmeleva, 2012 (p.909, Table I); Johan & al., 2012 (p.647, Table 1, 2, fig.2, salinity range); Miyashita & al., 2012 (p.1557, Table 2: occurrence); Johan & al., 2012 (2013) (p.1, Table 1); Zamora Terol, 2013 (p.174: Table 5); Zamora-Terol & Saiz, 2013 (p.376, Rem.: Table 3, egg production); Melo Jùnior & al., 2013 (p.363, Table 2, egg production); Jagadeesan & al., 2013 (p.27, Table 3, seasonal variation); Gubanova & al., 2013 (in press, p.4, Table 2); Jungbluth & Lenz, 2013 (p.630, Table I); Ohtsuka & Nishida, 2017 (p.565, Table 22.1); | | | | NZ: | 14 | | |
|
Distribution map of Oithona simplex by geographical zones
|
| | | | | | | | | | | | | | | 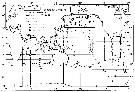 Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.140, Fig.91]. Issued from : S. Nishida in Bull. Ocean Res. Inst., Univ. Tokyo, 1985, No 20. [p.140, Fig.91].
Indo-Pacific geographical distribution of Oithona simplex.
Dotted line: AC, Arctic Convergence; SC, Subtropical Convergence. Arrows indicate the stations where the typical short form occured |
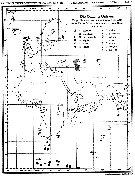 issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. "Valdiviella", 1917, 23. [Taf. I]. issued from : I. Rosendorn in Wiss. Ergebn. dt. Tiefsee-Exped. "Valdiviella", 1917, 23. [Taf. I].
The genus Oithona: Distribution of species sampled during the Deutsche Tiefsee Expedition 1898-99.
The size of symbols indicates the relative quantitty of the species. |
 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 39]. Oithona simplex (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 39]. Oithona simplex (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
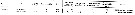 Issued from : S. Zamora Terol in Tesis Doc. Univ. Politèc. Catalunya, 2013 [p.82-83, Table 3]. Issued from : S. Zamora Terol in Tesis Doc. Univ. Politèc. Catalunya, 2013 [p.82-83, Table 3].
Comparison of egg production and clutch sizes among different species of Oithona from field and lab studies.
Also, see: Oithona attenuata, O. colcarva, O. davisae, O. nana, O. plumifera, O. setigera, O. similis, O. aruensis. |
 Issued from : T. Zuo, R. Wang, Ya-qu. Chen, Sh-wu Gao & Ke. Wang in J. Mar. Syst., 2006, 59. [p.169, Fig.9D]. Oithona simplex Issued from : T. Zuo, R. Wang, Ya-qu. Chen, Sh-wu Gao & Ke. Wang in J. Mar. Syst., 2006, 59. [p.169, Fig.9D]. Oithona simplex
Spatial of representative species abundance with isobaths shown in gray.
The circles are proportional in diameter to square root value of abundance.
Copepods collected vertically with plankton net (200 µm mesh aperture) from a depth near the bottom to the surface in autumn 2000 (18 October to 21 November).
Body length (mm): 0.37. |
| | | | Loc: | | | sub-Antarct. (SE Atlant.), South Africa (E), Brazil (S, Guarau estuary, Paranagua estuary, Cabo Frio, mouth of Amazon), Congo, Caribbean, Yucatan (Chetumal Bay), Porto Rico, Jamaica, G. of Mexico, off W Mississipi River mouth, Florida, off Cape Finisterre, Medit. (Alboran Sea, NW Basin, Adriatic Sea, Ionian Sea, Aegean Sea, Lebanon Basin, W Black Sea), Suez Canal, Port Taufik, Red Sea, Gulf of Oman, Arabian Sea, Kuwait, Madagascar (Nosy Bé), Indian, E India (Rushikulya estuary, Gulf of Mannar), Christmas Is., Perai River estuary (Penang), Straits of Malacca, Sangga estuary, Indonesia-Malaysia, Sarawak: Bintulu coast, Is. Aru, China Seas (Yellow Sea, East China Sea, South China Sea),Hong Kong, Taiwan (E, W, Tapong Bay), Xiamen Harbour, Okinawa (kabira Bay, lagoons), S Korea (Cheju Is.), S Japan (Kuchinoerabu Is.), Japan (Kyushu: Shijiki Bay, Ariake Bay, Australia (Exmouth Gulf, Wilson Inlet, G. of Carpentaria, Great Barrier), New Caledonia, Samoa Is., Palau Is., Pacif. (tropical), Hawaii, Kaneohe Bay, SE Pacif. | | | | N: | 80 | | | | Lg.: | | | (66) F: 0,44-0,3; M: 0,43-0,375; (109) F: 0,44-0,37; M: 0,47-0,37; (155) F: 0,46-0,43; M: 0,39; (179) F: 0,43-0,36; (434) F: 0,42-0,35; (624) F: 0,41-0,37; (634) F: 0,42-0,38; (649) F: 0,41; M: 0,37; (788) F: 0,44-0,4; M: 0,43-0,4 [grande forme]; 0,35-0,31 [petite forme]; (880) F: 0,30-0,44; M: 0,37-0,40; (991) F: 0,36-0,42; M: 0,39-0,41; (1047) F: 0,45; {F: 0,30-0,46; M: 0,31-0,47} | | | | Rem.: | epipelagic - ?.
This species appears less erratic than its locality records, given by the consulted authors may assume. The species is under-represented in the sampling nets because of the mesh size used.
After Gurney (1927, p.160), this species is characterised by the endopodal 2-segmented of P1, the presence of 3 outer spines on the 3rd exopodal segment of P4; the short anal somite, and its very small size. The Gurney's specimens differ from Farran's description in being of slightly larger size (0.43-0.47 mm) and in having spinules on the outer margins of the exopods in all legs. There is a long slender outer-edge seta on the 2nd basal segment of P3, and on P4 there is on this segment a short stiff seta which projects somewhat backwards and is consequently not visible in front.. In proportion ob body to abdomen, relative length of the segments of A1, and other details, Gurney's specimens agree fully with Farran's description. Gurney (1927) found this species in small numbers at Port Taufik (N Gulf of Suez), but not in the Canal.
Oithona simplex quinquesetosa Früchtl, 1923 (M)
Ref.: Früchtl, 1923 (p.455); 1924 b (p.74); Sewell, 1948 (p.377)
Loc.: Is. Aru
.
Oithona simplex Form typical (short form) Nishida,1985 (F,M)
Ref.: Nishida, 1985 a (p.91, figs.F,M: short form)
Loc.: Indien N, G. Arabe
Lg.: (627) F: 0,39-0,36; M: 0,41-0,39
.
Oithona simplex Large form Nishida,1985 (F)
Ref.: Nishida, 1985 a (p.96, Descr.F, figs.F: large form)
Lg.: (627) F: 0,4-0,38.
See in Oithona robertsoni Table 2 from McKinnon (2000, p.108).
See in DVP Conway & al., 2003 (version 1)
| | | Last update : 14/01/2021 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 04, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
