|
|
 |
|
Monstrilloida ( Order ) |
|
|
|
Monstrillidae ( Family ) |
|
|
|
Monstrilla ( Genus ) |
|
|
| |
Monstrilla grandis Giesbrecht, 1891 (F,M) | |
| | | | | | | Syn.: | ? Monstrilla grandis : T. Scott, 1904 (p.243, figs.F,M);
Monstrilla intermedia Kricchagin, 1877;
? Strilloma scotti: Isaac,1975 (n°144/145, p.6, 7, 9, figs.F,M, Rem.);
Monostrilla grandis : Shih & Young, 1995 (p.75) | | | | Ref.: | | | Giesbrecht, 1892 (p.586, 590, figs.F,M); Malaquin, 1901 (p.111); ? T. Scott, 1904 (p.243, figs.F,M); van Breemen, 1908 a (p.207, figs.F,M); Gurney, 1927 (p.168, figs.M, Rem.); Candeias, 1932 (p.8, figs.F:?); Rose, 1933 a (p.344, figs.F,M); Dolgopol'skaya, 1948 (p.177, figs.F,M); Shen & Bai, 1956 (p.233, figs.F,M); Fish, 1962 (p.30); Ramirez, 1971 b (p.379, figs.F,M, Rem.); Isaac, 1974 (p.128, figs.F, Rem.F,M); 1974 a (p.131); 1975 (p.9); Zheng Zhong & al.,1984 (1989) (p.272, figs.F,M); Huys & Boxshall, 1991 (p.155, 156, 158, 464, figs.F); Suarez-Morales, 1994 (p.265: Rem.); Grygier, 1995 a (p.9, 48, 66); Dias de O., 1996 (p.255, Rem.); Suarez-Morales, 2000 (p.107, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.836, figs.F); Vives & Shmeleva, 2010 (p.177, figs.F,M, Rem.); Suarez-Morales, 2011 (p.9, 10); Suarez-Morales & al., 2013 (p.620, Descr.M, figs.M, Rem.); Chang C.Y., 2014 (p.207, Redescr. F, figs F, Rem.; Suarez-Morales & al., 2017 (p.1822, Rem., figs.F, M) | 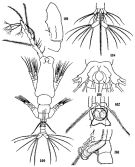 issued from : C.-j. Shen & S.-o. Bai in Acta Zool. sin., 1956, 8 (2). [Pl.XIII, Figs.100-105]. Male (from Chefoo, China): 100, habitus (dorsal); 101, A1 (segment ?); 102, P5 and genital segment; 103, idem (lateral left side). Female: 104, urosome (ventral); 105, thoracic segment 5 (ventral).
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution, The Ray Society, 1991. [p.158, Fig. 2.5.1]. Female (from Norfolk, England): A, habitus (lateral); B, cephalothorax (ventral); C, urosome (ventral). Nota: P5 female is biramous but both rami are fused to the protopodal part of the limb. The 3 setae on the exopod and the 2 setae on the endopod represent the maximum setation found in the Monstrilloida.
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution, The Ray Society, 1991. [p.159, Fig. 2.5.2]. Female: A, P2 (anterior view); B, genital double somite (ventral); C, caudal ramus (dorsal); D, P5 and genital double somite (lateral). Copulatory pores arrowed in B and D.
|
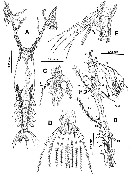 issued from : E. Suarez-Morales in Bull. Inst. R. Sci. nat. Belgique, Biologie, 70, 2000. [p.109, Fig.2]. Female (from Toulon Harbor, Mediterranean Sea): A, habitus (dorsal); B, right A1; C, head (lateral); D, P5 (anterior view); E, P5 and genital somite (lateral; digitiform processes arrowed). Nota: Cephalothorax (incorporating 1st pedigerous somite) accounting for 61 % of total body length. Forehead depressed in dorsal view. Anterior part of cephalothorax with ventral, rounded convex protuberance; another cuticulatr process, formed by a row of 8-11 rounded denticles is located adjacent to the first one forming a linear polip-like structure. Oral papilla lying midventrally 0.52 of way back along cephalothorax. Naupliar eye present. No cuticular ornamentation was observed on the surface of cephalothorax. A1 3-segmented (purported segments 3-5 fused); ratio of length segments 12.5:16.9:70.6 = 100.; A1 slightly longer than 53 % of total body length. P5 bilobed, fused medially; outer lobe with 3 long, plumose seta; inner lobe with 2 setae; all setae biserially setulated; a single finger-like process on distal part of inner margin of outer lobe. Urosome consisting of 5th pedigerous somite, genital double somite, and 2 free abdominal somites; Ratio of length of urosomal somites 31.4:38.5:19.1:11.1 = 100. Caudal rami 1.8 times longer than wide, moderately divergent, bearing 3 terminal, 1 dorsal, and 2 inner setae Nota: Giesbrecht (1891) original drawings does not observe on the outer lobe of P5 a finger-like process.
|
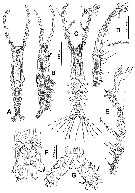 issued from : E. Suarez-Morales in Bull. Inst. R. Sci. nat. Belgique, Biologie, 70, 2000. [p.110, Fig.3]. Male (from Toulon Harbor, Mediterranean Sea): A, habitus (dorsal); B, idem (lateral; showing cuticular nipple-like process, arrowed). Nota: Cephalothorax representing about 47 % of total body length. Oral papilla located at about midway of way back along ventral surface of cephalothorax. Dorsal ocelli present, pigment cups relatively large, separated by the distance of two ocelli diameters, poorly developed, almost unpigmented, rounded in dorsal view. Two strongly chitinized, nipple-like cuticular processes located ventrally anterior to oral papilla (arrowed in fig.B). A1 5-segmented (segment 4 longest). 1st pedigerous somite incorporatd into cephalothorax. P5 represented by a pair of long seta each inserted on a basal lobe. Genital lappets with small subterminal rounded process, posteriorly directed.Urosome consisting of 4 segments (1st pedigerous somite, genital somite, and 2 free somites); ratio of lengths of genital somite and 2 free posterior somites 51.6: 25.8: 23.1 = 100. Caudal rami about 1.8 times longer than wide; each ramus with 2 protuberant processes (1 on outer margin, second on inner margin (arrowed in fig.G); 3 well developed terminal plus 2 additional setae inserted on the outer margin.setae
|
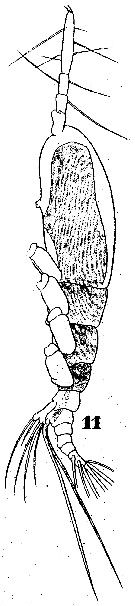 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.11]. Female: 11, habitus (lateral);
|
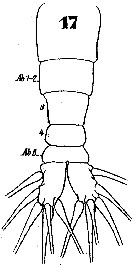 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.17]. Female: 17, urosome (dorsal).
|
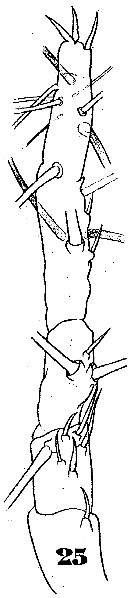 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.25]. Female: 25, A1 (anterior view).
|
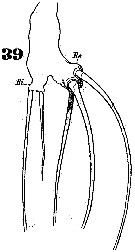 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.39]. Female: 39, P5.
|
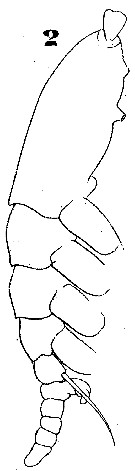 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.2]. Male: 2, habitus (lateral).
|
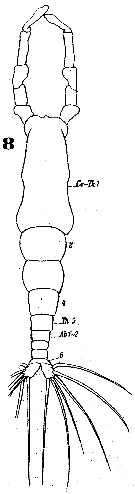 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.8]. Male: 8, habitus (dorsal).
|
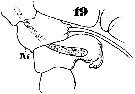 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.19]. Male: 19, urosome (lateral)
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.24]. Male: 24, A1 (dorsal view).
|
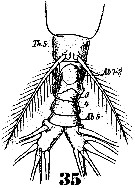 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.46, Fig.35]. Male: 35, urosome (ventral).
|
 issued from : M.J. Isaac in J. mar. biol. Ass. U.K., 1974, 54. [p.129, Fig.1]. Female: A, P5 of British and Chinese specimens; B, P5 of Naples specimens (after Giesbrecht).
|
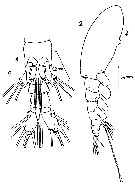 issued from : F.C. Ramirez in Physis, 1971, XXX, 81. [p.382, Lam.II, Figs.2, 4]. Female (from Argentina shelf: 38°30'S, 57°W): 2, habitus (lateral); 4, urosome (ventral), showing P5.
|
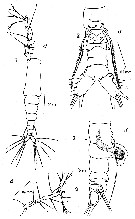 issued from : F.C. Ramirez in Physis, 1971, XXX, 81. [p.378, Lam.I]. Female: 4, A1. Male: 1, habitus (dorsal); 2, urosome (ventral, showing P5; 3, same (lateral).
|
 issued from : M.J. Grygier & S. Ohtsuka in Zool. J. Linnean Soc., 2008, 152. [p.499, Fig.29]. Female: Dorsal and lateral pore and pit seta patterns, from rear of cephalothorax through genital compound somite. Symbols: dots (three sizes) = pores; larger circles = pits of pit setae. Pattern based on SEM and light microscopical examination.
|
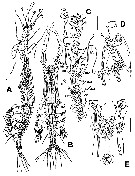 Issued from : E. Suarez-Morales, A. Carillo & A. Morales-Ramirez in J. Nat. Hist., 2013, 47 (5-12). [p.622, Fig.2]. As Monstrilla aff. grandis. Male (from Cahuita, Costa Rica): A-B, habitus (lateral and dorsal, respectively); C, left A1 (dorsal); D, cephalic area showing oral papilla and ventral protuberance (arrowed), lateral view); E, same (ventral). Scale bars: 100 µm (A, B); 50 µm (C-E). Nota: Cephalothorax representing up to 44.2% of total body length. Oral papilla protuberant, located at about midway (0.46) back along ventral surface of cephalothorax. Pair of relatively small lateral pigment cups well developed, separated by length of less than one eye diameter, moderately pigmented; ventral cup slightly larger than lateral cups. Wide-based, low rounded process on ventral surface protruding between bases of antennules (arrowed in fig.2 D). 2 pairs of nipple-like cuticular processes on anterior ventral surface between A1 bases and oral papilla (arrowed in fig.2D); additional triplet of nipple-like processes posterior to latter pairs (fig.2D,E). A1 elongate representing 55% of the body lenth and c.120% as long as cephalothorax, 5-segmented (segment 4 longest). Incorporated first pedigerous somite and succeding 3 three pedigerous somites. P5 represented by a pair of long biserially setulated setae each arising from small basal lobe on ventral surface of 5th pedigerous somite; 5th leg's setae (when stretchted backwards) reaching posterior end of anal somite. Genital apparatus strongly built, with enlarged base protruding ventrally; genital apparatus widely cylindrical, distally branched to form 2 short, rounded lappets each bearing subdistal rounded process posteriorly directed. Urosme consisting of 5 somites (5th pedigerous somite, genital somite, 2 free postgenital somites, and anal somite. Ratio of lengths of urosomites 25 : 16.2 : 21.6 : 19.3 : 17.8 = 100. Caudal rami subrectangular, moderately divergent, about twice as long as wide, each bearing 6 setae (3 terminal, 2 outer, and 1 on inner margin; dorsal medial seta shorter and more slender than other setae. Remarks: The Cahuita's specimen from Costa Rica (Caribbean Sea) was identified as M. grandis based on its possession of an elogate antennule which is close to 60% the length of the body, the presence of a P5 with a fused basal segment armed with a single long plumose seta, the shape and distal process of the genital apparatus and the presence of 6 caudal setae. There are some characters present in the male Costa Rican specimen that differ slifhtly from those in other known records: The relative length of A1 with respect to the total body length (48.5%: Naples; 54.8%: Argentina; 56.4%: China; 57%: Black Sea; 61.2%: Toulon); the oral papilla is located in slightly different position (44% : Naples; 49%: China; 53%: Toulon; 46%: Cahuita); the relative lengths of 4th antennular segment differ slightly (25.9%: Naples; 20%: Black Sea; 23%: China; 26.6%: Toulon; 24.4%: Argentina; 26.9%: Cahuita); the body size is also somewhat variable, less than 0.8 mm (Naples, Black Sea, Cahuita) up to more than 1.5 mm (Naples, Barbados, Argentina, Brazil, Scotland); the length of the singleP5 seta is similar in all cases, reaching the midlength of the caudal rami (when stretched backwards); in all cases the dorsal caudal seta (VII in Huys & Boxshall, 1991) is thinner and shorter than the others.
|
 Issued from : E. Suarez-Morales, A. Carillo & A. Morales-Ramirez in J. Nat. Hist., 2013, 47 (5-12). [p.624, Fig.3]. As Monstrilla aff. grandis. Male: A, P1 (dorsal); B, P2 (ventral); C, P3 (dorsal); D, P4 (dorsal); E, urosome (lateral view), showing P5 and features of genital apparatus, caudal rami cut short; F, same (ventral).
|
 Issued from : E. Suarez-Morales, A. Carillo & A. Morales-Ramirez in J. Nat. Hist., 2013, 47 (5-12). [p.625, Table 1]. Female: Formula armature of swimming legs P1 to P4. Spines denoted by Roman numerals, setae by Arabic numerals from outer margin of any segment given first, separated by a hyphen from the inner margin.
|
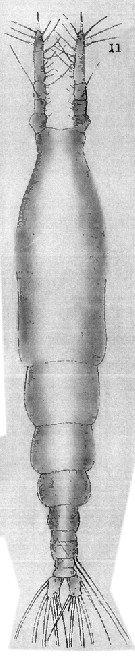 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XIII, fig.11]. With doubt. Female (from Loch Fyne: First of Clyde): 11, habitus (dorsal). Nota: For Rose (1933, p.345), the species depicted by T. Scott should be an another form.
|
 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XiV, fig.9-11]. With doubt Female: 9, A1; 10, P5; 11, Abdomen and caudal rami. Nota : A1 moderately stout, scarcely equal to ¼ the length of the body. P5 sub-cylindrical, about 2 times as long as wide, and with a somewhat bi-lobed extremity. The exterior lobe bears 3 setae, the inner one small but the other 2 elongated. The inner lobe appears to be furnished with only 1 apical seta but the dissection shows what appears to be the base of a seta on the inner margin of this lobe, the seta itself having probably been broken off (the position of this set ais indicated on the drawing by dotted lines. Nota : Abdomen 3-segmented, but the first, which is larger than the next two combined, divided into two portions by a pseudo-articulation ; the 2nd and 3rd segments sub-equal and together much smaller than the first segment. Caudal rami each provided with 6 setae, one being situated on the outer edge near the base of the segment while the others spring from the apex ; 1 of the apical setae near the inner edge very small.
|
 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XIII, fig.12]. With doubt. Male: 12, habitus (dorsal). Nota : A1 proportionally rather longer than those of the female, 5-segmented, the last segment being hinged to the preceding one. 1st and 2nd segments of abdomen not coalescent as in female. Armature of the caudal rami similar to that of the female. P5 resemble very closely to that of M. longicornis
|
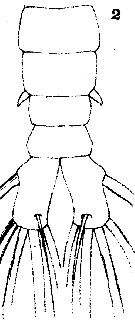 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XV, fig.2]. With doubt. Male: 2, abdomen and caudal rami. lapsus calami in the plate XV, fig.2 indicated as female.
|
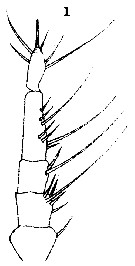 Issued from : T. Scott in Rep. Fishery Bd Scotl., 1904, 12 (3). [Pl. XV, fig.1]. With doubt. Male: A1.
|
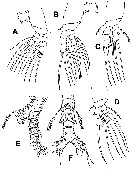
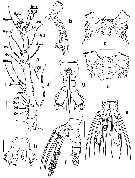 Issued from : E. Suarez-Morales, A. Goruppi, A. de Olazabal & V. Tirelli inJ. Nat. Hist., 2017, 51 (31-32) [p.1828, Fig.12]. Females specimens 'A' and 'B' from Trieste: Female specime 'A': a, right A1 with armature; b, cephalic area showing cuticular ornamentation (lateral view); c, same (dorsal view); e, urosome with P5 (ventral view); f, same (lateral view); g, same (ventral view, P5 omitted); h, P5 (ventral view, thumb-like processes arrowed) Female specimen 'B': d, cephalic area showing ventral ornamentation. Scale bars: 100 µm (a, e-g); 50 µm (b-d, h).
|
 Issued from : E. Suarez-Morales, A. Goruppi, A. de Olazabal & V. Tirelli in J. Nat. Hist., 2017, 51 (31-32) [p.1829, Fig.13]. Female specimen 'C' from Trieste: a, habitus (dorsal) (1 caudal ramus omitted); b, right A1 with armature (dorsal view); c, cephalic region (lateral view); d, urosome with P5 (lateral view); e, same (ventral view); f, same (dorsal); g, frontal area (dorsal); h, urosome of specimen 'D' with P5 (ventral view); i, same (dorsal view). Scale bars: 200 µm (a); 100 µm (b, d-f, h, i); 50 µm (c, g).
|
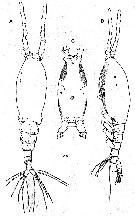 Issued from : C.Y. Chang in Anim. Syst. Evol. Divers., 2014, 30 (3). [p.208, Fig.1]. Female (from S. Korea): A, habitus (dorsal); B, same (lateral); C, cephalothorax (ventral). Scale bar = 200 µm. - Cephalothorax incorporating the 1st pedigerous somite, accounting for about 58 % of total body length. - Forehead slightly concave medially in dorsal view. - Anteriormost part of ventral surface with small, round protuberance furnished with 8-10 minute denticles. - All 3 cups of nauplius eye round, not well developed. - Oral papilla situated slighly posterior to midlength of cephalothorax (52.4 %), slightly protruding midventrally. 2 pairs of sensory pores ahead of oral papilla. - Urosome of 4 urosomites, together accounting for 21.7 % of total body length (excluding caudal setae). - Genital double-somite slightly swollen laterally, bearing paired, long ovigerous spines, inserted on middle of ventral surface, basally separated, representing 37.5 % of total body length, nearly twice as long as urosome, with tips pointed, not swollen, extending far beyond tips of caudal setae. - Anal somite trapezoidal, lacking wrinkles or striae both on dorsal and ventral surfaces; lateral margin nearly smooth without apparent notch. - Caudal rami a little divergent, about 1.8 times longer than wide, with small cuticular protuberance at inner distal corner of ramus and anterior to bases of lateral caudal setae; armed with 6 well-developed caudal setae, of which 3 distal, 2 lateral and 1 dorsal. - A1 long and slender, slightly longer than 48 % of total body length, about 73 % as long as cephalothorax; 3-segmented (ratio of lengths of segments: 12.4 : 17.2 : 70.4 = 100), with distal segment barely showing traces of fusion of 3 ancestral segments (purported segments 3-5). - P5 bilobed, both legs fused medially; outer lobe elongate, lateral margin curved outward, armed with 3 long, plumose setae; thumb-like process present on distal part of inner margin of outer lobe; inner lobe oblong, about 2 times as long as wide, its tip not quite reaching distal end of outer lobe, furnished with 2 apical setae of subequal length. Chang (2014, p.210) notes that Giesbrecht (1891, 1893) does not indicate the presence of a thumb-like or knob-like process on the outer lobe of P5 in the female. Isaac (1974) mentioned another difference between his material and Giesbrecht's (1892) figure: the 2 apical setae on the inner lobe of P5 were nearly equal in length in Isaac's specimens, but the inner seta was much shorter than the outer in Giesbrecht's figure. All the Korean specimens agree with Isaac's description. After Grygier (pers. comm.), it seems unlikely that the original material of M. grandis from the Southern Hemisphere (49° S, 65° W) could truly be conspecific with European and East Asian populations in the Northern Hemisphere
|
 Issued from : C.Y. Chang in Anim. Syst. Evol. Divers., 2014, 30 (3). [p.209, Fig.2]. Female: A, A1; B, P1; C, P4; D, urosome (ventral); R, 1st to 3rd urosomites (lateral). Scale bars: A = 200 µm; B-E = 100 µm.
|
 Issued from : C.Y. Chang in Anim. Syst. Evol. Divers., 2014, 30 (3). [p.210]. Female: Armature of swimming legs P1 to P4.
|
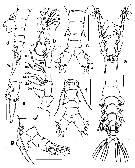 Issued from : E. Suarez-Morales, A. Goruppi, A. de Olazabal & V. Tirelli in J. Nat. Hist., 2017, 51 (31-32) [p.1824, Fig.10]. Male specimen 'A' from Trieste: a, habitus (dorsal); b, right A1; c, urosome (dorsal); Male specimen 'B': d, urosome (lateral, showing modified caudal seta, arrowed); e, detail of modified caudal seta. Male specimen 'C': f, urosome with P5 and genital complex (ventral); g, same (lateral ); Scale bars: 100 µm (a, b-d, f, g); 50 µm (e).
|
 Issued from : E. Suarez-Morales, A. Goruppi, A. de Olazabal & V. Tirelli in J. Nat. Hist., 2017, 51 (31-32) [p.1825, Fig.11]. Male specimen 'C' from Trieste: a, habitus (dorsal); b, cephalic region showing preoral ornamentation and protuberance (lateral); c, same (ventral). Male specimen 'D': d, cephalic region (ventral); e, left A1 with armature (dorsal view); f, P3; g, urosome and genital complex (dordsal); h, same (lateral); i, P5 and genital complex (ventral). Scale bars: 200 µm (a); 100 µm (b-i).
|
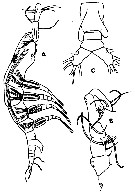 Issued from : R. Gurney in Trans. zool. Soc. London, 1927, 22. [p.168, Fig./27]; Male (from Kabret: Suez Canal): A, habitus (lateral); B, A1; C, abdomen (ventral). Nota: This specimen differs from Giesbrecht's description in the rather more anterior position of the oral tubercle, in the absence of a seta from the P5 (probably broken off), and in the relative lengths of the segments of A1, but agrees in the general proportions of the body and in possessing 6 furcal setae. Giesbrecht's specimens taken in the South Atlantic (65° W, 49° S), also found in the North Sea (Van Breemen, 1908)
| | | | | Compl. Ref.: | | | Kovalev, 2003 (p.47); Marques al., 2006 (p.297, tab.III); Dias & Bonecker, 2007 (p.270, 272, fig.3, tab.II); Oliveira Dias & Costa Bonecker, 2007 (p.282: Rem); Brylinski, 2009 (p.253: Tab.1, p.258: Rem.); S.C. Marques & al., 2011 (p.59, Table 1); El Arraj & al., 2017 (p.272, table 2); | | | | NZ: | 9 | | |
|
Distribution map of Monstrilla grandis by geographical zones
|
| | | | | | | | | | Loc: | | | Patagonia, Cape San Antonio, Brazil (Bahia, Recife, Vitoria)), Barbados Is., Porto Rico, E Costa Rica reef, Portugal, Bristol Channel, Scilly Is., English Channel, off Boulogne coast, North Sea, Portugal (Mondego estuary), Medit. (Algiers, Marseille, Harbour of Trieste, Black Sea), Red Sea, Viet-Nam, China Seas (Bohai Sea, Yellow Sea, East China Sea, Chefoo), S Korea, Hokkaido, Chile
Type locality: 49° S, 65° W | | | | N: | 32 | | | | Lg.: | | | (46) F: 3,75; M: 1,9-1,7; (277) F:1,8; M: 0,75; (466) F: 2,74-2,35; M: 1,31-0,70; (727) F: 4,25; M: 2; (745) F: 2,363; M: 1,637; (785) F: 4,12; M: 1,77; (828)* F: 1,5-0,8; M: 0,65; 0,61; (995) M: 1,06*; (1120)* M: 0,68; (1206)* F : 1,03-1,09; M: 0,64-1,05; (1208) F: 1,15-1,69; {F: 0,80-4,25; M: 0,61-2,00}
*: caudal rami excluded. | | | | Rem.: | In vertical haul 0-22 m.
See remarks for scotti.
After Dias (1996), the specimens off the coastal region of Espirito Santo (Brazil) differ from the description by Ramirez (1971), who found 4 spines and 3 branched setae on the terminal segment of the male's A1. The Dias' specimens had 1 plumose seta, in addition to the ones described by Ramirez. A1 females were described with 1 seta and 4 spines on segment 2; 1 seta and 3 smooth setae on segment 3 and 5 smooth setae, and 3 branched setae and 4 spines on segment 4. The Dias' specimens have 1 smooth setae and 5 spines on segment 2; 2 smooth setae and 1 spine on segment 3; and 5 spines, 5 smooth and 3 branched setae on segment 4. The males are somewhat shorter than previously reported.
Suarez-Morales (2000 c, p.111) remarks: Isaac (1975) included this species in the unvalid genus Strilloma, which was created as an interpretation of the partial separation of the genital double somite present in some species of Monstrilla; this very variable feature ha been considered to have a questionable taxonomical value. Hence, having 3 or 2 somites between the genital and the caudal rami in the male and the female, this species is correctly placed in the genus Monstrilla.
For Suarez-Morales & al. (2013, p.625) the size and morphometric variations found in M. grandis even in geographically adjacent areas (i.e. The Mediterranean), its wide distribution and the idea supposedly cosmopolitan species among Monstrilloida could represent species complexes highlights the need for a deeper analysis to define whether this nominal species represents a species complex. It will be useful to determine the status of records of M. grandis from other geographic regions than those of the central and south american coasts which could be conspecific.
Suarez-Morales & al. (2017, p.1827) underline a degree of intraspecific variability in the same locality as observed in the Gulf of Trieste. The comparative analysis of the cephalic ornamentation in females shows some variation among specimens from Trieste, always resembling the pattern observed in the males, with an anteriiomost set of bud-like processes arranged in two vertical rows, with 4 or 5 elements on each row (Figs.12c and 13c) or with an asymmetrical alignement of these rows (Fig.12d), the latter pattern depicted also by Huys & Boxshall (1991) and Chang (2014). A similar, but tighter pattern of these elements was observed by Suarez-Morales (2000c) in material from Toulon. The presence of frontal sensilla (Figs.12c and 13g) is also a variable intra-population character, it was absent in some specimens from Trieste (Fig.12d). In all populations compared female M. grandis lack a cephalic ventral protuberance (Fig. 12b and 13c)described in some males of this species.
The female A1 armature in specimens from Trieste showed some coincidences when compared with that of the male in pre-geniculation segments; elements 2v2,3 are the longest of group 2v-d (Figs.12 a and 13b) and element 4v1 is the longest of group 4v-d.
Also, the female genital double-somite has an incomplete suture visible on dorsal view and reaching halfway around the lateral margin (Figs. 12f and 13d) and observed in other populations as well (Dolgopolskaya, 1945; Huys & Boxshall, 1991; Suarez-Morales 2000c; Chang, 2014), but not in Argentinian specimens, in which the suture appears to be more extended around the ventral surface of the somite (Ramirez, 1971, figs.2 and 4).
The female P5 has, in all the populations compared, a thumb-like inner process on the exopodal lobe, it was clearly present in the specimens from Trieste (arrows in Figs. 12h and 13d-f, h,i). Giesbrecht's 1893 [1892] illustration of this appendage could be deemed as incomplete as noticed by Chang (2014)
It is requisite to use molecular evidence to solve taxonomic problem in which morphological patterns are not informative enough.
This presumably widespread species was originally described from the SW Atlantic (49°S, 65°W) by Giesbrecht (1891); the male was depicted from specimens collected at the Gulf of Naples (Giesbrecht, 1893 [1892], later, Suarez-Morales (2000 c) described the male from Toulon Bay. The species has been recorded from different geographic areas of the world, including the Black-Sea (Dolgopolskaya, 1948), the China Seas (Shen & Bai, 1956), the Carribbean area (Fish, 1962; Nutt & Yeaman, 1975), Argentina (Ramirez, 1971, geographically closest to the type locality), the Mediterranean (Isaac, 1974, 1975), Chile (Marin & Antezana, 1985), Costa Rica (Suarez-Morales & al., 2013), Korea (Chang, 2014)
The relative length of the cephalothorax with respect to the total body length: the shortest cephalothorax (43 %) in the Costa Rica population, followed by specimens reported by Giesbrecht (1892) from Naples and by Ramirez (1971) from Argentina (47 %), by Suarez-Morales (2000c) from Toulon (48 %), Dolgopolskaya (1948) from the Black Sea (48.7 %, Shen & Bai (1956) from China (49 %) and a range of 49.2-50.5 % was observed in the male specimens from Trieste.
The A1 length with respect to the total body length shows some variation: 48 % (Naples), 55 % (Argentina), 56 % (China), 57 % (Black Sea), 60 % (Costa Rica), 61 % (Toulon), 59.3 % - 64.8 % (Trieste)
There are subtle differences on the position of the oral papilla along the cephalothorax: 44 % (Naples), 45-43 % (Trieste), 46 % (Costa Rica), 49 % (China), 53 % (Toulon);
The relarive lengths of the male 4th antennular segment: 20 % (Black Sea), 23 % (China), 24.4 % (Argentina), 26 % (Naples), 26.6 % (Toulon), 27 % (Costa Rica), 28-32 % (Trieste)
The body size is remarkably variable: less than 0.8 mm (Naples, Black Sea, Costa Rica) up to more than 1.5 mm (Naples, Barbados, Argentina, Brazil, Scotland); in the samples from Trieste, the male specimens length ranged between 0.64-1.05 mm (average = 0.86 mm, n = 15).
The cuticular ornamentation of the ventral surface of the cephalothorax was described only in the specimens from Toulon Bay (Suarez-Morales, 2000 c) and the pattern differs from the Costarrican male (Suarez-Morales & al., 2013); in the former specimens, there is only 1 pair of strongly chitinised nipple-like processes vs 3 pairs of such structures (plus an additional medial one) in the Costarrican specimen, these arranged in a relatively tight pattern (Suarez-Morales & al., 2013, fig.2E. In specimens from Trieste the ornamentation resembles that of Costarrican specimens, with 3 pairs of nipple-like processes, but in in our specimens these are arranged in a more separate pattern and, instead of having a single medial papilla-like structure posterior to this group, there are additional bud^like structures grouped either in a transverse row of 3 (arrows in fig.9, b, e, f) or in 2 symmetrical inclined rows each with 2-4 such elements §fig. 11, b, d); these bud-like groups are associated to a ventral cephalic protunerance, which is pronounced in our specimens (arrows in fig. 9 f and 11 b) and in those reported by Giesbrecht (1892), but is weakly developed in the specimens from Toulon (Suarez-Morales, 2000 c, fig. 2C) and Costa Rica §Suarez-Morales & al., 2013, fig. 2D).
Also, the Toulon male specimens have 1 pair of protuberant processes on the inner and outer margins of the caudal rami (Suarez-Morales, 2000 c, fig. 3G); these have not been reported in any other of the examined records, including the males from Trieste (fig. 10 f and 11 g);
The length of the single P5 seta is similar in all cases, reaching midlength of the caudal rami or even slightly beyond it when stretched backwards (fig. 10 f, g and 11 g, i).
Details of the male antennulary armature (Grygier & Ohtsuka, 1995) of our specimens can be compared only with records from Toulon (Suarez-Morales, 2000 c) and Costa Rica (Suarez-Morales & al., 2013): In the 1st segment, element 1 has some variation, even in individuals from Trieste; it can be as long as a segment (fig. 10 b) as in the Toulon specimens (Suarez-Morales, 2000 c, fig.2E) or about half its length (fig. 11 e) as in the Costarrican specimens (Suarez-Morales & al., 2013, fig. 2 C). In the 3 groups compared, elements 2 v2,3 are the longest of group 2 v-d. This is also discernable from illustrations by Giesbrecht (1892) (Naples), Shen & Bai (1956) (China) and Ramirez (1971) (Argentina). On the 4th segment, element 4 v1 is consistently the longest of the setal group 4 v-d in the populations compared (Giesbrecht, Ramirez, Suarez-Morales, 2000 c and Suarez-Morales & al., 2013), but showed some variation in size among the specimens from Trieste (fig. 10 b and 11 e).
At least two of the males from Trieste showed a modified (proximally expanded) caudal seta (fig. 10 d, r), but the other specimens observed have normal caudal setae (fig. 10 a, f and 11 a, g). It is to mention that some females showed an expansion on the 2 caudal setae (arrows in fig. 13i)
In addition, P3 basipodal seta is relatively longer in our specimens (fig. 11 f) than in the specimen from Costa Rica (Suarez-Morales & al., 20013, fig.3 C) | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 27, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
