|
|
 |
|
Cyclopoida ( Order ) |
|
|
|
Corycaeidae ( Family ) |
|
|
|
Corycaeus ( Genus ) |
|
|
|
Ditrichocorycaeus ( Sub-Genus ) |
|
|
| |
Corycaeus (Ditrichocorycaeus) affinis McMurrich, 1916 (F,M) | |
| | | | | | | Syn.: | Corycaeus japonicus Mori, 1937 (1964) (p.138, figs.F,M); Shen & Bai, 1956 (p.229, figs.F,M); Shen & Lee, 1963 (p.589); Delalo, 1968 (p.139); Palomares-Garcia & Vera, 1995 (tab.1); Shih & Young, 1995 (p.76); Suarez-Morales & Gasca, 1998 a (p.112);
Ditrichocorycaeus affinis : Boxshall & Halsey, 2004 (p.495); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrographic conditions); Ohtsuka & al., 2015 (p.123, Table 2); Zakaria & al., 2016 (p.1, Table 1, Rem.); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence) | | | | Ref.: | | | Campbell, 1929 (p.327, Rem.M); Davis, 1949 (p.75, figs.F,M); Tanaka, 1957 (p.92, figs.F,M, Rem.); Chen & al., 1974 (p.58, figs.F,M); Zheng Zhong & al., 1984 (1989) (p.268, figs.F,M); Kang & al., 1990 (p.53, figs.F,M); Kim & al., 1993 (p.270); Chihara & Murano, 1997 (p.964, Pl.213,214: F,M); Bradford-Grieve & al., 1999 (p.888, 975, figs.M); Marquez-Rojas & al., 2014 (p.9, Descr.F,M, figs. F,M, Rem.); | 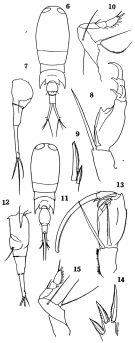 issued from : O. Tanaka in J. Fac. Agricult. Kyushu Univ., 1957, 11 (1). [Pl.8, Figs.6-15]. Female (from Japanese waters): 6, habitus (dorsal); 7, urosome (lateral right side); 8, A2; 9, terminal spine of exopod of P2; 10, P4. Nota: The abdominal segments and furca in the proportional lengths 36:26:38; anterior segment1.8 times as long as wide. Genital segment 9/4 as wide as long. Anal segment little wider at the proximal, and less than 2 times as long as it is wide at the distal (11:6). Furcal rami 1.7 times as long as anal segment (17:10), and about 9 times as long as wide at the proximal (17:2), rami divergent. Male: 11, habitus (dorsal); 12, urosome (lateral left side); 13, A2; 14, terminal spines of exopod of P1; 15, P4. Nota: The abdominal segments and furca in the proportional lengths 50:22:28. Eyes separated by about 1/3 the greatest diameter. 3rd thoracic segment has sharp backward pointing wings which extend to proximal 1/3 of the genital segment. The demarcation between 3rd and 4th thoracic segment indistinct. 4th thoracic segment produced into small pointed processes. Genital segment1.7 times as long as wide (20:12), ventral surface with a ventrally pointing hook. Anal segment 1/3 the length of the distal margin of the segment. Furcal rami straight, about 6 times as long as wide (11:2).
|
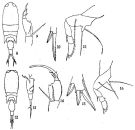 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.17, Figs.8-16]. Female (from China Seas): 8, habitus (dorsal); 9, A2; 10, terminal spines of exopod of P2; 11, P4. Male: 12, habitus (dorsal); 13, urosome (lateral right side); 14, A2; 15, terminal spines of exopod of P1; 16, P4.
|
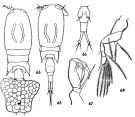 issued from : C.-j. Shen & S.-o. Bai in Acta Zool. sin., 1956, 8 (2). [Pl.IX, Figs.65-69]. As Corycaeus affinis. Female (from Chefoo): 65, habitus with egg sac (dorsal); 66, urosome (dorsal); 67, A2. Nota: Head and 1st thoracic segment separated. Caudal rami divergent posteriorly. Male: 68, habitus (dorsal); 69, P4. Head and 1st thoracic segment fused. Caudal rami parallel to each other.
|
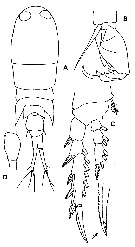 issued from : Y.S. Kang, S.-H. Huh & S.S. Lee in J. Ocanol. Soc. Korea, 1990, 25 (2). [Pl. 3, Figs.A-D]. Female (from Korean waters): A, habitus (dorsal); B, A2; C, P2; D, genital segment (lateral, left side). Nota: Caudal rami divergent, and approximately 7-8 times longer than width. The seta on basipod 1 of A2 3 times as long as the seta on basipod 1 of A2. Apical spine of exopod of P2 with 2-3 teeth on the inner anterior side. Genital segment as long as the caudal rami, and the genital segment with one ventral process.
|
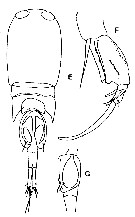 issued from : Y.S. Kang, S.-H. Huh & S.S. Lee in J. Ocanol. Soc. Korea, 1990, 25 (2). [Pl. 3, Figs.E-G]. Male: E, habitus (dorsal); F, A2; G, genital segment (lateral, left side). Nota: Lateral points of 3rd thoracic segment extended to 1/3 the genital segment. Genital segment with one ventral process like a hook. Caudal rami about 7 times longer than width. Basipod 2 of A2 with one row of strong teeth on the outer distal and one row of fine teeth on the central part.
|
 issued from : T. Mori in The Pelagic copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.76, Figs.3-5, 8-9, 11]. As Corycaeus japonicus. Female: 3-4, habitus (dorsal and lateral, respectively); 5, A2; 8, P2; 9, P3; 11, P4.
|
 issued from : T. Mori in The Pelagic copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.76, Figs.1-2, 6-7, 10]. As Corycaeus japonicus. Male: 1-2, habitus (lateral and dorsal, respectively); 6, A2; 7, P1; 10, P4.
|
 Issued from : B. Marquez-Rojas, O. Diaz-Diaz, L. Troccoli, J. Morales & L.M. Marcano in Métod. Ecol. Sistemat., 2014, 9 (3). [p.20, Fig.2, a-f]. Temporary under the taxon Corycaeus affinis before other animals examined.
Female (from Cariaco Gulf, Venezuela): a, habitus (dorsal); b, A2; c, P4.
Male: d, habitus (dorsal); e, A2: f, P4.
Nota female:
- Head and 1st pedigerous somite partly fused.
- posterior points of 3rd and 4th thoracic somites teach 1/2 of the genital somite.
- Urosome 2-segmented, about longer than half of metasome.
- Genital somite and caudal rami, approximately of equal length.
- Anal somite slightly more than half the length of the genital, somite.
- Genital somite with a medial hook, directed posteriorly on the proximal part on the ventral side.
- Basipodite 1 of A2 with one seta 3 times longer than the seta on the basipodite 2.
- Endopod P4 with one or two small segments showing 2 setae.
Nota male: Body similar than in female, excepted the sexual dimorphism.
- Lateral points of the 3rd thoracic somites extending at 1/3 of genital segment.
- genital segment with a ventral process like a hook.
- Basipodite 2 of A2 with a row of strong teeth on the outer distal border and a line of fine teeth in the central portion.
- Anal somite and caudal rami, together, of the same length that the genital segment.
- Caudal rami longer than the anal somite.
| | | | | Compl. Ref.: | | | Yamazi, 1958 (p.155, Rem.); Northcote & al., 1964 (p.1069, Table II); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio/Kuroshio transitional area, Table 7, 10); Shih & al., 1971 (p.155); Hirota & Hara, 1975 (p.115, fig.5); Hanaoka, 1977 (p.267, 301, abundance); Chen Q-c, 1980 (p.795); Hirota 1981 (p.19, Table 1, length-weight-CHN); Hirakawa & al., 1990 (tab.3); Yoo, 1991 (tab.1); Hwang & Choi, 1993 (tab.3); Myung & al., 1994 (tab.1); Shih & Young, 1995 (p.76); Kotani & al., 1996 (tab.2); Park & Choi, 1997 (Appendix); Noda & al., 1998 (p.55, Table 3, occurrence); Hsieh & Chiu, 1998 (tab.2); Suarez-Morales & Gasca, 1998 a (p.112); Dolganova & al., 1999 (p.13, tab.1); Ueda & al., 2000 (tab.1); Lo & al., 2001 (1139, tab.I); Hsieh & al., 2004 (p.398, tab.1, p.399, tab.2); Rezai & al., 2004 (p.490, tab.2); Lan & al., 2004 (p.332, tab.1); Kang & al., 2004 (p.1524); Lo & al., 2004 (p.89, tab.1); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Choi & al., 2005 (p.710: Tab.III); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: stations group); Hwang & al., 2006 (p.943, tabl. I); Lan Y.-C. & al., 2008 (p.61, Table 1, % vs stations, Table 2: indicator species, Rem.: p.71); Tseng L.-C. & al., 2008 (p.153, fig.5, Table 2, occurrence vs geographic distribution, indicator species); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons, fig.8, table 3: indicator species); Ohtsuka & al., 2008 (p.115, Table 4, 5); Chiba & al., 2009 (p.1846, Table 1, occurrence vs temperature change); C.-Y. Lee & al., 2009 (p.151, Tab.2); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Fazeli & al., 2010 (p.153, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Zhang G.-T. & Wong, 2011 (p.277, fig.6, 7, 8, abundance, indicator); Tseng L.-C. & al., 2011 (p.239, Table 3, abundance %); Beltrao & al., 2011 (p.47, Table 1, fig.3, density vs time); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Huo & al., 2012 (p.1, Table 1: dominance); Lee D.B. & al., 2012 (p.88, co-occurrence); Youn & al., 2012 (p.33, Table 1, 2: seasonal composition); Lee J. & al., 2012 (p.95, Table 2: abundance, annual & seasonal distribution); Moon & al., 2012 (p.1, Table 1); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Palomares-Garcia & al., 2013 (p.1009, Table I, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, pers. comm.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Tseng & al., 2013 (p.507, seasonal abundance); Mendoza Portillo, 2013 (p.37: Fig.7, seasonal dominance); Seo & al., 2013 (p.448, Table 1, occurrence); Oh H-J. & al., 2013 (p.192, Table 1, occurrence); Ohtsuka & Nishida, 2017 (p.565, 578, Table 22.1); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence) | | | | NZ: | 10 + 1 doubtful | | |
|
Distribution map of Corycaeus (Ditrichocorycaeus) affinis by geographical zones
|
| | | | | | | | | | | | | Loc: | | | Namibia (in Stander & De Decker, 1969) , E Medit. (W Egyptian coast), Red Sea, Gulf of Oman; Straits of Malacca, G. of Thailand, Hong Kong, China Seas (Bohai Sea, Yellow Sea, South China Sea, off Minjiang Riv.), Taiwan Strait, Taiwan (E, W, SW, NW, N, Mienhua Canyon, NE), S, E & W Korea, Muan Bay, Chunsu Bay, Asan Bay, Yongsan River estuary, Geumo Is., Korea Strait, Japan Sea, Japan ( Kuchinoerabu Is., Ariake Bay, Seto Inland Sea, Tanabe Bay, Tokyo Bay), Charlotte Queen Is., British Columbia, Vancouver Is., Nitinat Lake, Strait of Georgia, San Francisco Estuary, W Baja California (Magdalena Bay), La Paz, Gulf of California, Bahia Cupica (Colombia), ? Venezuela (Cariaco Gulf) | | | | N: | 94 | | | | Lg.: | | | (59) F: 1,1; M: 0,9-0,83; (91) F: ± 1,1; M: ± 0,9; (107) F: 1,25-1,01; M: 0,83; (109) F: 0,87-0,75; M: 0,77-0,62; (141) M: 0,83; (351) F: 0,92; M: 0,83; {F: 0,75-1,25; M: 0,62-0,90} | | | | Rem.: | C. Davis (1949) states that C. japonicus Mori (1937) is identical with affinis, abundant in the waters near Friday Harbor (Washington State) at certain times of the year.
Gardner & Szabo, 1982 (p.98) consider C. affinis a synonym of C. anglicus.
According to Kang & al. (1990, p.53): Tanaka (1957) reported that the length of the caudal rami of female was 9 times its width and 1.7 times the anal segment. Chen & al (1974) reported that the caudal rami of female was 7 times its width and 1.6 times the anal segment.
Observed in the ballast waters of ships in San Francisco.
Fazeli & al. (p.162: Table 1) note the occurrence for the first time this species in the Indian Ocean at Chabahar Bay, Oman Sea.
Zakaria & al. (2016) note for the first time the occurrence of this species in the East Mediterranean Sea, probably an new lesseptian species.
Ohtsuka & Nishida (2017, p.578) point to the Japanese coasts adjacent to the Kuroshio Current are distinguished by warm-water species C. affinis.
For Boxshall & Halsey (2004, p.491) the subgenus (Dahl, 1912) is considered as full generic status pending phylogenetic revision of the whole family at the generic level. | | | Last update : 09/12/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 15, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




 affinis - Plate 1 of morphological figures',750,500);)
 affinis - Plate 2 of morphological figures',750,500);)
 affinis - Plate 3 of morphological figures',750,500);)
 affinis - Plate 4 of morphological figures',750,500);)
 affinis - Plate 5 of morphological figures',750,500);)
 affinis - Plate 6 of morphological figures',750,500);)
 affinis - Plate 7 of morphological figures',750,500);)
 affinis - Plate 8 of morphological figures',750,500);)

 affinis - Plate 1 of morphological figures){kind=link}
 affinis - Plate 2 of morphological figures){kind=link}
 affinis - Plate 3 of morphological figures){kind=link}
 affinis - Plate 4 of morphological figures){kind=link}
 affinis - Plate 5 of morphological figures){kind=link}
 affinis - Plate 6 of morphological figures){kind=link}
 affinis - Plate 7 of morphological figures){kind=link}
 affinis - Plate 8 of morphological figures){kind=link}
