|
|
 |
|
Cyclopoida ( Order ) |
|
|
|
Oncaeidae ( Family ) |
|
|
|
Triconia ( Genus ) |
|
|
| |
Triconia dentipes (Giesbrecht, 1891) (F,M) | |
| | | | | | | Syn.: | Oncäa dentipes Giesbrecht, 1891; 1892 (p.591, 603, 774, Descr.F, figs.F);
Oncaea dentipes : Farran, 1929 (p.210, 286); Rose, 1933 a (p.300,figs.F); ? Wilson, 1942 a (p.198, fig.F); Sewell, 1948 (p.392); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Neto & Paiva, 1966 (p.31, Table III); Vilela, 1968 (p.32); Vinogradov, 1968 (1970) (p.268); ? Corral Estrada, 1970 (p.222, Descr.M, figs.M); Razouls, 1972 (p.95, Annexe: p.115, figs.F); ? Chen & al., 1974 (p.42, figs.F,M); ? Razouls, 1974 b (p.239, figs.F); Dawson & Knatz, 1980 (p.9, 10, figs.F,M); Björnberg & al., 1981 (p.668, figs.F); Malt, 1983 a (p.6, figs.F, Rem.); Zheng & al., 1984 (p.104, figs.F); Böttger-Schnack, 1990 (p.869, tab.III); Ohtsuka & al., 1996 a (p.91); Chihara & Murano, 1997 (p.980, Pl.224: F,M); Boxshall, 1998 (p.226)
Ref. compl.: V.N. Greze, 1963 a (tabl.2); Shmeleva, 1964 a (p.1068); De Decker & Mombeck, 1964 (p.13); Mazza, 1966 (p.73); Pavlova, 1966 (p.45); Deevey, 1971 (p.224); Björnberg, 1973 (p.364, 388); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Vives & al., 1975 (p.56, 57, tab.II, IV, XII); Vives, 1976 (p.104); Peterson & Miller, 1976 (p.14, Table 1, 3, abundance vs interannual variations); 1977 (p.717, Table 1, seasonal occurrence); Dessier, 1979 (p.207); Vaissière Séguin, 1980 (p.23, tab.2); Vives, 1982 (p.295); Kovalev & Shmeleva, 1982 (p.85); Scotto di Carlo & al., 1984 (p.1044); Regner, 1985 (p.11, Rem.: p.41); Lozano Soldevilla & al., 1988 (p.61); Böttger-Schnack & al., 1989 (p.1089) ; Böttger-Schnack, 1992 (p.304); Seguin & al., 1993 (p.23, Table 2: abundance, %); Böttger-Schnack, 1995 (p.92); Shih Young, 1995 (p.77); Böttger-Schnack, 1996 (p.1086); 1997 (p.409); Hure & Krsinic, 1998 (p.104); Krsinic, 1998 (p.1051); Siokou-Frangou, 1999 (p.479); Seridji & Hafferssas, 2000 (tab.1); Vukanic, 2003 (p.139, tab.1); Prusova & Smith, 2005 (p.76); Zervoudaki & al., 2006 (p.149, Table I); Hwang & al., 2007 (p.25); Humphrey, 2008 (p.85: Appendix A); Hafferssas & al., 2010 (p.1281, Table III, abundance vs spatial distribution); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Zaafa & al., 2014 (p.67, Table I, occurrence); | | | | Ref.: | | | Böttger-Schnack, 1999 (p.43, 60, figs.F,M, Redescr., Rem.); Böttger-Schnack & al., 2004 (p.1130, tab.1, Rem.); Di Capua & Boxshall, 2008 (p.1409, figs.F); Vives & Shmeleva, 2010 (p.341, figs.F,M, Rem.); Böttger-Schnack & Machida, 2011 (p.111, Table 1, 2, fig.2, 3, DNA sequences, phylogeny); Wi & al., 2012 (p.852: Table 1, p.853: Table 1, 2) ; Cho & al., 2013 (p.14: Rem.) | 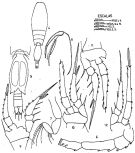 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.55, figs.4-12]. As Oncaea dentipes. With doubt. Male (from Canarias Is.): 4, habitus (dorsal); 5, urosome (ventral); 6, A1; 7, A2; 8, Mxp; 9, P1; 10, P2; 11, P3; 12, P4. Nota: Prosome length/wide = 2.6 times. Abdomen 5-segmented, the abdominal segments and furca in the proportional lengths 61:5:4:6:13:11 = 100.
|
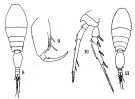 issued from : Q.-c Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.7, Figs.8-11]. As Oncaea dentipes. With doubt. Female (from China Seas): 8, habitus (dorsal); 9, Mxp; 10, P4. Male: 11, habitus (dorsal). Nota: According Böttger-Schnack (1999, p.70) this form is more similar to T.giesbrechti
|
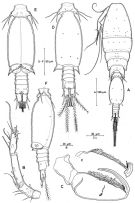
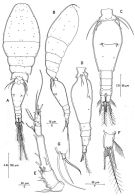 issued from : R. Böttger-Schnack in Mitt. hamb. zool. Mus. Inst., 1999, 96 [p.61, Fig.9]. Female (from Red Sea): A, habitus (dorsal); B, idem (lateral right side); C, urosome (dorsal); D, idem (lateral left side); E, A1; F, caudal ramus (dorsal, long terminal seta omitted); G, P5 (dorsal). Nota: Proportional lengths (%) of urosomites and caudal rami 8.9: 56.1:6.8: 8.0:11.0:9.3. A1 6-segmented, relative lengths (%) of segments measured along posterior non-setigerous margin 6.8:20.5:46.8:11.7:2.9:11.2. P6 represented by operculum closing off each genital aperture, armed with long spine and minute blunt spinule. Egg-sacs paired, oblong, each sec containing 4 eggsin a row (diameter about 0.040 mm).
|
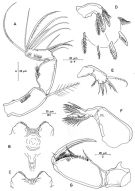 issued from : R. Böttger-Schnack in Mitt. hamb. zool. Mus. Inst., 1999, 96 [p.62, Fig.10]. Female: A, A2 (anterior); B, labrum (anterior); C, idem (posterior); D, Md (showing individual elements); E, Mx1; F, Mx2; G, Mxp.
|
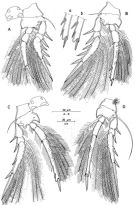 issued from : R. Böttger-Schnack in Mitt. hamb. zool. Mus. Inst., 1999, 96 [p.64, Fig.11]. Female: A, P1 (anterior); B, P2 (anterior, intercoxal sclerite omitted) [a, b, terminal part of endopod, showing variation in spine length]; C, P3 (anterior); D, P4 (anterior, intercoxal omitted).
|
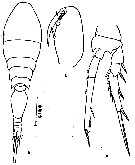
 issued from : R. Böttger-Schnack in Mitt. hamb. zool. Mus. Inst., 1999, 96 [p.67, Fig.13]. Female (from Adriatic Sea): A, urosme (dorsal, long inner terminal seta omitted); B, P5 (dorsal); C, caudal ramus (dorsal, long terminal seta omitted); D, endopod of P2 (anterior); E, endopod of P4 (anterior).
|
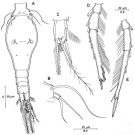
 issued from : R. Böttger-Schnack in Mitt. hamb. zool. Mus. Inst., 1999, 96 [p.66, Fig.12]. Male (from Red Sea): A, habitus (dorsal); B, A1; C, Mxp (anterior) [c, inner margin of basis and proximal part of endopodal claw, showing fused pectinate spine); D, urosome (dorsal); E, idem (ventral, spermatophores fully developped); F, idem (lateral left side). Nota: Proportional lengths (%) of urosomites and caudal rami 8.2:54.9:5.1: 5.1:6.7:10.2:9.8. A1 4-segmented, relative lengths (%) of segments measured along posterior non-setigerous margin 9.2:19.5:43.1:28.2.
|
 issued from : R. Böttger-Schnack in Mitt. hamb. zool. Mus. Inst., 1999, 96 [p.68, Table 2]. Comparison of morphological characters of T. dentipes and related species from the Red and Mediterranean Seas. ODS = outer distal spine; OSDS = outer subdistal spine; no = number; ? = uncertain.
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.104, Fig.62]. As Oncaea dentipes. Female: a, habitus (dorsal); b, Mxp; c, P4. Scale bar in mm.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Figs.7, 17]. As Oncäa dentipes. Female: 7, urosome (dorsal); 17, habitus (lateral).
|
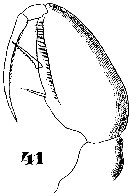 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Fig.41]. As Oncäa dentipes. Female: 41, Mxp.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.47, Figs.51, 52]. As Oncäa dentipes. Female: 51, P1; 52, P4.
|
 issued from : C. Razouls in Th. Doc. Etat Fac. Sc. Paris VI, 1972, Annexe. [Fig.75]. Female (from Banyuls, G. of Lion): A, urosome; B, A1; C, A2; D, Mxp; E, Mx2; F, P1; G, P3; H, P4; I, P2.
|
 Issued from : K. Cho, W.-S. Kim, R. Böttger-Schnack & W. Lee in J. Nat. Hist., 2013. [p.16, Table 2]. Morphological characters of Triconia dentipes of the Triconia dentipes-subgroup from the north-eastern equatorial Pacific. Nota: Med.Sea: Mediterranean Sea; *: Pacific Ocean near Galapagos Islands (3°S, 99°W)Orn: ornamentation; L: length; W: width; Compare with related species and form variants of the Triconia dentipes-subgroup from the north-eastern equatorial Pacific and from other regions: T. pacifica, T. elongata, T. giesbrechti (Pacific form), T. giesbrechti (Red Sea form), T. elongata, T. constricta.
|
 Issued from : K. Cho, W.-S. Kim, R. Böttger-Schnack & W. Lee in J. Nat. Hist., 2013. [p.17, Table 2 (Continued)]. Morphological characters of Triconia dentipes of the Triconia dentipes-subgroup from the north-eastern equatorial Pacific. Nota: Orn: ornamentation; L: length; W: width; OSDS: outer subdistal spine; ODS: outer distal spine; CP: distal conical process Compare with related species and form variants of the Triconia dentipes-subgroup from the north-eastern equatorial Pacific and from other regions: T. elongata, T. pacifica, T. giesbrechti (Pacific form), T. giesbrechti (Red Sea form), T. constricta.
| | | | | Compl. Ref.: | | | Guangshan & Honglin, 1984 (p.118, tab.); Böttger-Schnack & al., 2001 (p.1029, tab.1, 2); Krsinic & Grbec, 2002 (p.127, tab.1); Nishibe & Ikeda, 2004 (p.931, Tab. 2, 5); Nishibe & al., 2009 (p.491, Table 1: seasonal abundance); Böttger-Schnack & Schnack, 2009 (p.131, Table 3); Mazzocchi & Di Capua, 2010 (p.429); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Gubanova & al., 2013 (in press, p.4, Table 2); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Mazzocchi & al., 2014 (p.64, Table 4, 5, abundance); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2, seasonal composition); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence). | | | | NZ: | 14 + 1 doubtful | | |
|
Distribution map of Triconia dentipes by geographical zones
|
| | | | | | | | | | | | | Loc: | | | South Africa (E), SW Atlant. (Brazil), off Bermuda, Angola (Baia Farta), Congo, off Cape Verde Is., off Mauritania, Canary Is., Portugal, Ibero-moroccan Bay, off W Tangier, Medit. (M'Diq, Alboran Sea, Habibas Is., Balearic Basin, Banyuls, Marseille, Ligurian Sea, Tyrrhenian Sea, N & S Adriatic Sea, Ionian Sea, Aegean Sea, Black Sea, Lebanon Basin), G. of Aqaba, Red Sea (N-S), Arabian Sea, SW Indian, China Seas (East China Sea, South China Sea), Taiwan, Japan (Tosa Bay, Kuroshio & Oyashio regions), Pacif. (W equatorial), N New Zealand, Oregon (of Newport), California (San Pedro Bay), off Galapagos, ? Chile (37° S) | | | | N: | 67 ? | | | | Lg.: | | | (35) F: 0,52; (46) F: 0,52-0,51; (109) F: 0,52-0,45; M: 0,57-0,5; (180) F: 0,51; M: 0,39; (327) F: 0,51; (788) F: 0,49-0,44; M: 0,39-0,36; (866) F: 0,42-0,52; M: 0,36-0,38; (1014) F: 0,478; (1023) F: 0,5-0,57; {F: 0,42-0,57; M: 0,36-0,57}
The mean female size is 0.497 mm (n = 14; SD = 0.0390), and the mean male size is 0.421 mm (n = 7; SD = 0.0811). The size ratio (male : female) is 0.89 (n = 3; SD = 0.1771) or ± 89 %. | | | | Rem.: | Mesopelagic. Epipelagic in the Gulf of Naples.
The small sizes of this form (0.36 - 0.57 mm), being a generality for the species of this genus, explain the geographical discontinuities due to the mesh size of the sampling nets.
This species is close to Triconia elongata and Triconia giesbrechti with which it could have been confused, hence certain contestable locality records.
See also Triconia constricta.
After Cho & al. (2013, p.14) this species was originally described by Giesbrecht (1891) based on females collected at 03° S, 99° W.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 5 corresponding to small sac-spawning detritivorous. | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 17, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
