|
|
 |
|
Calanoida ( Order ) |
|
|
|
Clausocalanoidea ( Superfamily ) |
|
|
|
Aetideidae ( Family ) |
|
|
|
Gaetanus ( Genus ) |
|
|
| |
Gaetanus tenuispinus (Sars, 1900) (F,M) | |
| | | | | | | Syn.: | Chiridius tenuispinus Sars, 1900 (p.67, Descr.F, figs.F); 1902 (1903) (p.30, figs.F, Rem.); Frontier, 1977 a (p.14);
Gaidius boreale Wolfenden, 1902 (p.365);
Gaidius similis : A. Scott, 1909 (p.51, figs.M, Rem.F,M); Sewell, 1948 (p.516);
Gaidius glacialis Brady, 1918 (p.19, figs.juv.M,
no F);
Gaidius tenuis : Dessier, 1979 (p.204);
Gaidius tenuispinus : Mràzek, 1902 (p.512, 521, figs.F, Rem.); Sars, 1903 (p.162, Descr.M, figs.M); Farran, 1905 (p.33, Rem.F); Pearson, 1906 (p.13, Rem.); Vanhöffen, 1907 (p.521, figs.F); Farran, 1908 b (p.32); Esterly, 1911 (p.316, figs.M); Wolfenden, 1911 (p.223, figs.F); With, 1915 (p.89, figs.F,M); Sars, 1925 (p.46); ? Farran, 1926 (p.249: juv.); 1929 (p.208, 231, Rem.); Sewell, 1929 (p.100, Rem.); Wilson, 1932 a (p.53, figs.F,M); Rose, 1933 a (p.98, figs.F); Jespersen, 1934 (p.58, fig.14); Hardy & Gunther, 1935 (p.158); Jespersen, 1939 (p.48, Rem., Table 29, 30); 1940 (p.19); Vervoort, 1949 (p.15, figs.F); Brodsky, 1950 (1967) (p.162, figs.F,M); Vervoort, 1951 (p.81, Rem.); 1952 d (n°45, p.3, figs.F,M); Østvedt, 1955 (p.14: Table 3, p.63); Vervoort, 1957 (p.6, 58, Rem.); Tanaka, 1957 a (p.60, figs.F, non M); Minoda, 1958 (p.253, Table 1, 2, abundance); Grice & Hart, 1962 (p.287, table 3); Grice, 1963 a (p.495); M.W. Johnson, 1963 (p.89, Table 1, 2); Owre & Foyo, 1967 (p.43, figs.F,M); Maclellan D.C., 1967 (p.101, 102: occurrence); Ramirez, 1969 (p.58, figs.F, Rem.); Tanaka & Omori, 1970 (p.124, figs.F,M); Wheeler, 1970 (p.10, figs.M); Shih & al., 1971 (p.32, 201); Vidal, 1971 a (p.15, 26, 114, figs.F,M); Bradford, 1971 b (p.21, fig.M); 1972 (p.38, figs.F, Rem.); Park, 1975 a (p.11); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Park, 1978 (p.127, figs.F,M); Pipe & Coombs, 1980 (p.223, figs. 1, 2, table 1, vertical distribution); Björnberg & al., 1981 (p.630, figs.F,M); Gardner & Szabo, 1982 (p.220); Cummings, 1984 (p.163, Table 2); Ward, 1989 (tab.2); Suarez, 1992 (App.1); Zmijewska, 1987 (tab.2a); Razouls, 1994 (p.76, figs.F, M, Rem.); Richter, 1994 (tab.4.1a); Chihara & Murano, 1997 (p.688, Pl.45,46: F,M); Mauchline, 1998 (tab.30, 33, 64);
Ref. compl.: Damas & Koefoed, 1907 (p.398, tab.II, III); Lysholm & Nordgaard, 1921 (p.13); Wilson, 1942 a (p.188); Lysholm & al., 1945 (p.13); Sewell, 1948 (p.348, 496, 499, 516, 526, 545, 549, 556, 559, 566); C.B. Wilson, 1950 (p.234); Fagetti, 1962 (p.18); De Decker & Mombeck, 1964 (p.12); Grice & Hulsemann, 1965 (p.223); Mazza, 1966 (p.70); Harding, 1966 (p.17, 66); Furuhashi, 1966 a (p.295, vertical distribution in Kuroshio region, Table 9, 10) ; Grice & Hulsemann, 1967 (p.15); Dunbar & Harding, 1968 (p.318); Dowidar & El-Maghraby, 1970 (p.268); Park, 1970 (p.476); Deevey, 1971 (p.224); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 a (p.335); Rudyakov, 1972 (p.886, Table 1: sinking rate); Björnberg, 1973 (p.323, 386); Vives & al., 1975 (p.41, tab.II, XII); Vaissière & Séguin, 1980 (p.23, tab.3); Buchanan & Sekerak, 1982 (p.41, vertical distribution); Kovalev & Schmeleva, 1982 (p.83); Huntley & al., 1983 (p.143, Table 2); Sameoto, 1984 (p.213, Table 1); Cummings, 1984 (p.176); Gordon & al., 1985 (p.89, Table 2, fish diet); Groendahl & Hernroth, 1986 (tab.1); Lozano Soldevilla & al., 1988 (p.58); Kosobokova, 1989 (p.26); Suarez & Gasca, 1991 (tab.2); Mumm, 1993 (tab.1, fig.2); Ward & al., 1995 (p.195, Table 2); Errhif & al., 1997 (p.422); Hanssen, 1997 (tab.3.1); Voronina & Kolosova, 1999 (p.71); Atkinson & Sinclair, 2000 (p.46, 50, 51, 53, 55, zonal distribution); Lapernat, 2000 (tabl. 3, 4); Razouls & al., 2000 (p.343, tab. 5, Appendix); Ward & Shreeve, 2001 (p.50, tab.3); Yamaguchi al., 2002 (p.1007, tab.1); Sameoto & al., 2002 (p.12); G. Harding, 2004 (p.43, figs.F); Ikeda & al., 2006 (p.1791,Table 2); Ward & al., 2008 (p.241, Appendix II ); Morales-Ramirez & Suarez-Morales, 2008 (p.517); Darnis & al., 2008 (p.994, Table 1); Homma & Yamaguchi, 2010 (p.965, Table 2); Homma & al., 2011 (p.29, Table 2, 3, abundance, feeding pattern: suspension feeders); Marrari & al., 2011 (p.1599, table 2, abundance, composition, interannual variation); 2011 a (p.1614, Table 2, Fig.2A, 5, 6);
Gaidius pungens : Bradford & Jillett, 1980, (p.61, figs.M, no F); Matsuno & al., 2012 (Table 2); Michels & al., 2012 (p.369, Table 1, occurrence frequency) | | | | Ref.: | | | Park, 1975 a (p.30, figs.F,M); Vives, 1982 (p.291); Roe, 1984 (p.357); Hopkins, 1985 (p.167); Markhaseva & Razzhivin, 1992 (1993) (p.611); Sirenko & al., 1996 (p.348); Markhaseva, 1996 (p.221, figs.F,M); Bradford-Grieve & al., 1999 (p.879, 922, figs.F,M); Vives & Shmeleva, 2007 (p.586, figs.F,M, Rem.) |  issued from : T. Park in Antarct. Res. Ser. Washington, 1978, 27. [p.128, Fig.15]. As Gaidius tenuispinus. Female: A, habitus (dorsal view); B, idem (left lateral side); C, posterior part of metasome and urosome (dorsal view); D, idem (left lateral side); E, genital segment (ventral view); F, forehead (left lateral side); G, rostrum (anterior view); H, A2; I, Md.
|
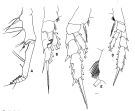 issued from : T. Park in Antarct. Res. Ser. Washington, 1978, 27. [p.129, Fig.16]. As Gaidius tenuispinus. Female: A, Mxp., B, P1 (anterior view); C, P2 (posterior view); D, P4 (posterior); E, medial part of coxa of P4 (posterior view).
|
 issued from : T. Park in Antarct. Res. Ser. Washington, 1978, 27. [p.130, Fig.17]. As Gaidius tenuispinus. Male: A, habitus (right lateral side); B, forehead (left lateral side); C, A2; D, P1 (anterior view); E, P5 (posterior view).
|
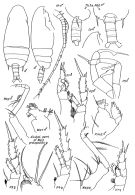
 issued from : E.L. Markhaseva in Proc. Zool. Inst. RAN, St. Petersburg, 1996, 268. [p.223, Fig.177]. Female. Ce: forehead; P.md: mandibular palp.; (a): NW Pacif.: Kuril Trench; (b): Arctic Basin (Central part); other figures: Arctic Basin (Central part). Nota: Posterior corners of last thoracic segment with spines always exceeding the midlength of genital segment (usually covering 3 to 4/5), sometimes reaching its posterior border. A1 reaching the midlength of urosome, or the end of caudal rami. Endopodal segment 1 of Md with 2 setae (1 of them very small, poorly visible); segment 2 with 9 terminal setae. P1 with indistinct subdivision between exopodal segments 1 and 2 (some authors noted complete division). P4 coxopodite with group of about 20 thin spines.
|
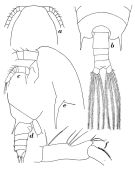
 issued from : E.L. Markhaseva in Proc. Zool. Inst. RAN, St. Petersburg, 1996, 268. [p.224, Fig.178]. Female (from Arctic Basin: Central part). Male (from Norwegian Sea)
|
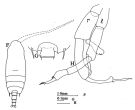 issued from : J.M. Bradford & J.B. Jillett in Mem. N.Z. Oceanogr. Inst., 86, 1980. [p.62, Fig.41, F-H]. As Gaidius pungens. Male: F, habitus (dorsal); G, distal part of metasome and genital segment (dorsal); H, P5 (r = right leg; l = left leg).
|
 Issued from : W. Vervoort in Zool. Verh., Leiden, 1949, 5. [p. 16, Fig.7].. As Gaidius tenuispinus. Female (from Moluccan Sea): a, forehead (dorsal); b, distal thoracic segments and urosome (dorsal); c, forehead (lateral); d, last thoracic segment and urosome (lateral right side); e, spine of the left lateral thoracic margin; f, 1st basal segment of Mxp. Nota: Head and 1st thoracic segment fused, 4th and 5th fused. The abdominal segments and furca in the proportional lengths 42:16:14:12:16 = 100. A1 (24 free-segments) is as long as the cepphalothorax. A2, mouth parts and legs differ scarcely from descriptions given by Sars (1900, 1902, 1903) and With (1915). The basipod of Mxp has no trace of a lamella, the terminal lobe, besides carrying a tubercle, has 3 setae (only 2 are figured by With). The 1st basal segment of P4 Nota: The 1st basal segmant of P4 carries 5 to 6 rows of hair like tubes on the posterior surface in the distal corner. The Indo-Malaysian specimens differ scarcely from Atlantic specimens.
|
 issued from : G.O. Sars in An Account of the Crustacea of Norway. Vol. IV. Copepoda Calanoida. Published by the Bergen Museum, 1903. [Pl. XVIII]. As Chiridius tenuispinus. mp2 = Mxp. Female.
|
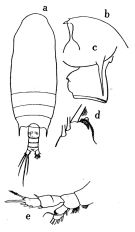 issued from : O. Tanaka in Publ. Seto Mar. Biol. Lab., 1957, VI (1). [p.61, Fig.37]. As Gaidius tenuispinus. Female (from Sagami, Japan): a, habitus (dorsal); b, forehead (lateral); c, last thoracic segment and genital somite (lateral left side); d, distal margin of 1st basal segment of Mxp; e, P1. Nota: Proportional lengths of urosomites and furca 41:18:13:10:18 = 100. A1 (24-segmented) estends about to distal margin of 2nd urosomal segment.
|
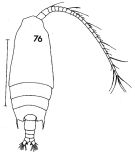 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.62, Lam. XI, fig.76 ]. As Gaidius tenuispinus. Female (from off Mar del Plata): 76, habitus (dorsal). Scale bar in mm: 1.5.
|
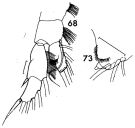 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.60, Lam. X, figs.68, 73 ]. As Gaidius tenuispinus. Female (from off Mar del Plata): 68, P1; 73, distal margin of basipodal segment 1 of Mxp. Scale bars in mm: 0.05 (68).
|
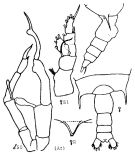 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.162, Fig.75]. As Gaidius tenuispinus. Female (from Arctic.): lest thoracic segment and urosome (dorsal); idem (lateral right side); R, rostrum; S1, P1. Nota: Exopod of P1 with partially separated 1st and 2nd segment. Male: S5, P5.
|
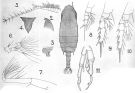 issued from : A. Scott in Siboga-Expedition, 1909, XIX a. [Plate VII, Figs.1-11]. As Gaidius similis. Male (from Indonesia-Malaysia): 1, habitus (dorsal); 2, forehead (lateral); 3, last thoracic and genital segments (left side); 4, rostrum; 5, A1; 6, A2; 7, Mxp; 8, P1; 9, P2; 10, P4; 11, P5. Nota: The 1st exopodal segment of the right P5 has a lamella-like process on the middle of the inner margin, which may be easily overlooked.
|
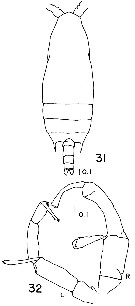 Issued from : E.H. Wheeler, Jr in Smiths. Contr. Zool., 1970, 55. [p.20, Figs.31-32]. As Gaidius tenuispinus. Male: 31, habitus (dorsal); 32, P5 (L = left leg; R = right leg).
|
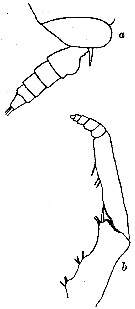 issued from : R.N. Wolfenden in Die Marinen Copepoden der Deutschen Südpolar-Expedition 1901-1903, 1911. [p.223, Fig.11]. As Gaidius tenuispinus. Female: a, posterior part cephalothorax and urosome; b, Mxp.
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1911, 6 (14). [Pl.28, Fig.26]. As Gaidius tenuispinus. Male (from San Diego Region): 26, forehead (lateral).
|
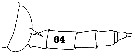 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1911, 6 (14). [Pl.29, Fig.64]. As Gaidius tenuispinus. Male: last thoracic segment and urosome (lateral, left side). nota: While the San Diego specimen does not agree in all respects with the description given by Sars (1903), there is no apparent reason for making a separate species; P5 are exactly as shown by Sars (1903, Suppl. Pl 6).
|
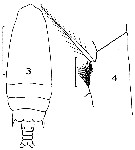 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.39, Fig.7, (3-4]. As Gaidius tenuispinus. With doubt. Female (from Kaikoura, New Zealand): 3, habitus (dorsal); 4, basipod segments of P4. Nota: This rare species may be distinguished from G. pungens Giesbrecht, by the two-segmented endopod of P2.
|
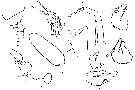 issued from : C. With in The Danish Ingolf-Expedition, 1915, III (4). [p.90, Text-Fig.23]. As Gaidius tenuispinus. Female (from N Atlantic): a, last thoracic segment and urosome (lateral); b, left Mx2 (posterior); c, basipod of left P4 (posterior); d, left P5; e, right Mx2 (anterior, showing position of parasites); f, g, paraites; h-j, contorted bodies attached to the body-wall. Nota: Caudal rami 1.6 as long as wide and a little wider than the anal segment is long. A1 reach to about the end of the 3rd urosomal segment ; 2nd segment 1.2 as long as 8-9 which is almost 1.3 as long as segment 13 and only a little shorter than segment 20 ; segments 19 and 22 almost of equal length, and almost 1.1 shorter than segment 2 ; the posterior seta of the segment 23 ringed, and about 3 times as long as the segments 24 and 25, which are well separated. A2 : exopod 1.3 times as long as endopod. Mx1 : inner lobe 1 with about 10 faily strong teeth on the anterior surface, and on the posterior surface about 10 similar ones ; the inner lobe 2 has 4 setae, the inner lobe 3 with 5 + a sensory lobe and anteriorly a fairly strong spinulation, the basipode 3 has 4 posterior setae and 1 anterior seta but no spinulation anteriorly ; the 1st endopodite has 3 anterior setae + 1 posterior seta, 2nd endopodite with 3 anterior setae + 1 posterior seta and the 3rd endopodite with 5 setae ; the exopodite has 11 setae, the outer lobe with 7 long + 2 shorter setae. Mx2 very characteristic by a strongly and suddenly conxex outer margin of the basipod, with a short concavity in the middle. P4 : 3rd exopodite 3 times as long as wide and the terminal seta with 25 teeth, partly armed with accessory ones ; the 1st basipodite with 30-40 stiff thin lamellous bristles, which are placed in about 5 rows along the inner margin, and in 2-3 parallel ones on posterior surface . In several adult females curiously twisted bodies (igs.h-j) to the number of 1 or 2 were found; they are by a delicate stalk fastened to the skin of the body wall between the insertion of the Mx1 and Mx2; their content is finely granular.
|
 issued from : C. With in The Danish Ingolf-Expedition, 1915, III (4). [Pl. III, Figs2, a-n]. As Gaidius tenuispinus. Female: ea, rostrum (dorsal view); 2b, basal segments of left A2; 2c, left Mxp lobe 4 of basipod 2 (anterior); 2d, terminal seta of the last exopodal segment ofP1; 2e, oral surface of the labrum. Male: 2f, forehead (lateral); 2g, rostrum (ventral); ; 2h, Mx2; 2i, terminal seta of the last exopodal segment of P1; 2j, exopodite 2 of left P5 (in exterior view); 2k, tip of exopodite 3 of left P5; 2l, endopod of right P5 (seen from left side); 2m, exopod 1-2 of left P5; 2n, right P5 (3rd segment of exopodite from inner side). Nota : Rostrum fairly long and shows, terminally, trace of bifurcation. A1 segments 8-9 partly fused with 10, and segments 12 and 13 with 14 ; segments 20 and 21 and 24 and 25 completely fused ; segment 22 is 1.1 as long as segment 17 and 1.2 as long as segment 2. Md : Manducatory potrtion has rudimentary teeth ; basipodite 3 has no setae . Mx1 : Fairly long inner lobe 2 with a number of rounded sensory organs. Mx2 : Small, thin-skinned organs ; exterior margin of basipodite witout convexity ; the inner surface possesses an indistinct number of short partly fused lobes, the setae of which are more or less transformed into thin-skinned more or less setiformed appendages with a swollen base ; endopod with a few setae. P5 : on the right leg the endopod is distally swollen and contains clear alveolar structures ; exopodite 1 (1-2 ?) has in most specimens a lateral protuberance, and shows trace of segmentation (fig.2m) ; exopodite 2 has inwards a lamelliform process which surrounds a shallow cup facing backwards and outwards (fig.2n) ; exopodite 3 elongate and curved with a terminal hooked hairy process. Left leg (Text-fig.22d) appears more slender than in Sars figure ; the endopod shows a trace of segmentation is long and slender, distally somewhat enlarged and bearing a short hair ; the exopodite 2 has inwards a hairy protuberance (fig.2j), and the exopodite 3 is very elongate with a distal fairly long outer seta and a terminal seta (fig.2k).
|
 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.43]. As Gaidius tenuispinus. Female. Nota:Female: last thoracic segment with posterior spines long, reaching past the middle of the genital segment. Exopodal segments 1 and 2 of P1 partially separated (arrowed). Coxa of P4 (B1) with 5 to 6 rows of hair-like tubes (arrowed).
|
 Issued from : E. Vanhöffen in Zool. Jb., Syst., 1907, 25 (4) [Pl. 22, Figs.33]. Female (from 70°25'N, 51°10'W): 33, A1.
|
 Issued from : E. Vanhöffen in Zool. Jb., Syst., 1907, 25 (4) [Pl. 21, Figs.27, 27 a]. Female: 27, terminal seta of distal exopoal segment of swimming legs.
|
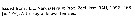 Gaetanus tenuispinus female: 1 - Cephalon without frontal spine. 2 - Posterior corners of last thoracic segment with long spines. 3 - Exopodal segment 1 of A2 without setae; segment 2 with 2 setae. 2nd internal lobe of Mx1 with 4 setae. Exopod of P1 3-segmented with 2 external spines. 4 - Mxp protopodite without lateral plate. 5 - Md palp base with 2 setae. 6 - Spines of last thoracic segment long, with reaching the midlength of genital segment.
|
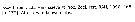 Gaetanus tenuispinus Gaetanus tenuispinus male: 1 - Cephalon without frontal spine. 2 - Cephalon without crest in anterior part. 3 - Exopodal segment 3 of left P5 stylet-like. Body size < 5 mm. 4 - Posterior corners of last thoracic segment with spines. 5 - Spines exceeding posterior border of genital segment.
|
 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.20, Figs.61, 62]. As Gaidius tenuispinus. Female (from Ross Sea): 61, coxa of P4; 62, basipod of Mxp. Scale bar: 100 µm.
|
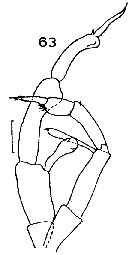 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.21, Fig.63]. As Gaidius tenuispinus. Male (from Ross Sea): 63, P5. Scale bar: 1 mm. Nota: Posterior thoracic borders extended into narrow spines which reach midway down the urosomal segment 2. Tanaka's male has a shorter left endopod.
|
 Issued from : C. Séret inThesis 3ème Cycle UPMC, Paris VI, 1979. [Pl. XII, Figs.71-73]. Female (from off Kerguelen Is.): 71-72, habitus (dorsal and lateral, respectively); 73, right Mxp.
|
 Issued from : C. Razouls in Ann. Inst. océanogr., Paris, 1994, 70 (1). [p.76]. Caractéristiques morphologiques de Gaetanus tenuispinus as Gaidius tenuispinus femelle et mâle adultes. Terminologie et abbréviations: voir à Calanus propinquus. Un individu femelle provenant de la station kerfix (Kerguelen présente les caractères suivants: longueur totale de 3, 55 mm. Pr/Ur = 3,74; Endo de P2 à deux segments très nets sur les deux faces; B1 de P4 présente sur le bord interne une seule rangée de poils raides (qui tourne distalement vers l'autre face) ; B1 de Mxp présente de très petites dents distales en ''coussinet'' (visible seulement au fort grossissement du microscope); les pointes postérieures de Th5 atteignent ± la fin du segment génital (selon l'état de contraction du segment génital par rapport au métasome). le Bi de Mxp ne correspond pas exactement à la figure 41D, p.62 in Bradford & Jillett (1980).. Le B1 de P4 ne présente pas 5-6 rangs de poils raides comme l'indique Vervoort (1957, p.59), qui malheureusement n'en donne pas de dessin.
| | | | | Compl. Ref.: | | | Tremblay & Anderson, 1984 (p.3); Hopkins, 1985 (p.197, Table 1, gut contents); Dearborn & al., 1986 (p.1, predation by benthic star, as tenuispinis); Hopkins & Torres, 1988 (p.82, tab.2); Ferrari & Dearborn, 1989 (p.1315, predator); Rau & al., 1991 (p.1, isotopic forms vs feeding); Donnelly & al., 1994 (p.171, chemical composition); Kosobokova & al., 1998 (tab.2); Suarez-Morales & Gasca, 1998 a (p108); Mumm & al., 1998 (p.189, Fig.6); Auel, 1999 (tab.2); Lapernat, 2000 (tabl.3, 4); Kosobokova & Hirche, 2000 (p.2029, tab.2); Holmes, 2001 (p.50); Auel & Hagen, 2002 (p.1013, tab.2); Berasategui al., 2005 (p.313, fig.2); Hopcroft & al., 2005 (p.198, table 2); Hop & al., 2006 (p.182, Table 4); Kosobokova & al., 2007 (p.919, Tab.2, 4, 6, 7, fig.2, eggs); Blachowiak-Samolyk & al., 2007 (p.2716, Table 2); Blachowiak-Samolyk & al., 2008 (p.2210, Table 3, biomass); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Muelbert & al., 2008 (p.1662, Table 1); Galbraith, 2009 (pers. comm.); Laakmann & al., 2009 (p.741, fig.2, vertical distribution, lipids analysis); Laakmann & al., 2009 (p.679, fig.2, 4, Table 2); Park & Ferrari, 2009 (p.143, Table 4, 7: common deep water species, Appendix 1, biogeography); Kosobokova & Hopcroft, 2010 (p.96, Table 1, fig.7); Hidalgo & al., 2010 (p.2089, Table 2); Mazzocchi & Di Capua, 2010 (p.423); Bucklin & al., 2010 (p.40, Table 1, Biol mol.); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); 2011 a (p.1231, Table 2: abundance, biomass, as Gaidius renuispinosus); Hirche & Kosobokova, 2011 (p.2359, Table 3, abundance, biomass %); Kosobokova & al., 2011 (p.29, Table 2, figs.4, 8, Rem.: Arctic Basins); Pepin & al., 2011 (p.273, Table 2, seasonal abundance); Pomerleau & al., 2011 (p.1779, Table V); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Salah S. & al., 2012 (p.155, Tableau 1); Laakmann & al., 2012 (p.535, Table 1, fig.2, Rem.: mol. Biol.); Teuber & al., 2013 (p.1, Table 1, abundance vs oxygen minimum zone); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Lidvanov & al., 2013 (p.290, Table 2, % composition); Ward & al., 2014 (p.305, Table 7, seasonal and abundance in the ''Discovery'' Investigations in the 1930s); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Bode & al., 2015 (p.268, Table 1, chemical components, trophic level, geographic zone); Smoot & Hopcroft, 2016 (p.1, fig.7, vertical distribution); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Belmonte, 2018 (p.273, Table I: Italian zones); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 22 + 1 doubtful | | |
|
Distribution map of Gaetanus tenuispinus by geographical zones
|
| | | | | | | | | | | | | | | | | | 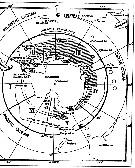 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Reports - Ser. B, Vol. III, 1957 [Fig.40]. As Gaidius tenuispinus. issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Reports - Ser. B, Vol. III, 1957 [Fig.40]. As Gaidius tenuispinus.
Chart showing the geographical distribution (white circle) in the seas surrounding the Antarctic continent.
Nota: In this chart the area frequented by whaling vessels has been hatched. The Antarctic circle (66°.5 S) has been drawn as a broken line. The numbers I to VI refer to the sectors into which the Antarctic seas are divided according to Mackintosh (1942) (after Vervoort, 1951). |
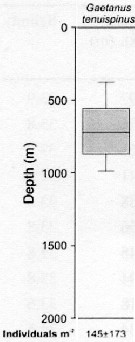 issued from : S. Laakmann, M. Stumpp & H. Auel in Polar Biol., 2009, 32. [p.682, Fig.2]. issued from : S. Laakmann, M. Stumpp & H. Auel in Polar Biol., 2009, 32. [p.682, Fig.2].
Vertical distribution (stages C3 to C6 from Antarctic Polar Front: Atlantic sector).
Error bars encompass the 5th to the 95th percentile. Abundance data are given as mean ± SD. |
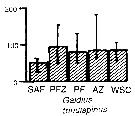 Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3] Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3]
Gaidius tenuispinus (= Gaetanus australis) from Scotia Sea.
Median and interquartile ranges of copepods (nos /m2) in the five water zones; from north to south these are SAF Subantractic Front area, PFZ Polar frontal Zone, PF Polar Front area, AZ Antarctic Zone, WSC Weddell-Scotia Confluence area/ East Wind Drift.
Numbers on the plots are upper interquartiles where these could not be scaled. |
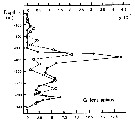 Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.202, Fig.4, B]. As Gaidius tenuispinus. Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.202, Fig.4, B]. As Gaidius tenuispinus.
Abundance (10/m3) and biomass (g dry mass/m3) profiles at the oceanic station from the shelf break in water 4000 m deep off Bird Island, South Georgia (53°04'S, 39°51'W) in January 1990.
Values on the horizontal axes at the base of each profile represent abundance and the one above biomass. Solid line = midday haul; hatched line = midnight haul. |
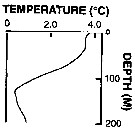 Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.198, Fig.1, B (modified C.R.)]. Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.198, Fig.1, B (modified C.R.)].
Profile temperature-depth at the oceanic stations from the shelf break in water 4000 m deep off Bird Island, South Georgia, in January 1990. |
 Issued from : C. Séret in Thesis 3ème Cycle, UPMC, Paris 6. 1979, Annexe. [p.31]. Issued from : C. Séret in Thesis 3ème Cycle, UPMC, Paris 6. 1979, Annexe. [p.31].
Geographical occurrences of Gaetanus tenuispinus in the Indian Ocean and Antarctic zone. [after publications from: Brady, 1883, 1918; Thompson, 1900; Wolfenden, 1908, 1911; With , 1915; Rosendorn, 1917; Farran, 1929; Sewell, 1929, 1947; Brady & Gunther, 1935; Steuer, 1929, 1392, 1933; Ommaney, 1936; Vervoort, 1957; Tanaka, 1960; Brodsky, 1964; Seno, 1966; Andrews, 1966; Grice & Hulsemann, 1967; Seno, 1966; Frost & Fleminger, 1968; Voronina, 1970; Zverva, 1972].
C. Séret notes the occurrence at stations 56°S, 70°E and 46°S, 64°E. |
 Issued from : S. Laakmann, M. Kochzius & H. Auel in Deep-Sea Res. I, 2009, 56. [p.745, Fig.2 b]. Issued from : S. Laakmann, M. Kochzius & H. Auel in Deep-Sea Res. I, 2009, 56. [p.745, Fig.2 b].
Vertical distribution abundance of copepodite stages C3 to C6 of Gaetanus spp. +: No occurrence; solid line depicts bottom profile (right axis); station 853 over the Yermak Plateau (81°22'N, 6°52'E) is separated from the other stations on the transect; other stations between East Greenland Current, the Fram Strait and the West Spitsbergen Current, into south return Atlantic Current (77°46'N-79°36'N, 6°20'E-7°29'W).
Deep-sea copepods collected from August 20 to September 16, 2006. |
| | | | Loc: | | | Antarct. (Croker Passage, Peninsula, Weddell Sea, SW & SE Atlant., Indian, SW & SE Pacif., Ross Sea), South Georgia, sub-Antarct. (SW Atlant., Indian, Kerguelen Is., SE Pacif.), South Africa (E), Angola, Congo, G. of Guinea, SW Atlant., off NW Cape Verde Is., off Morocco-Mauritania, Cap Ghir, Canary Is., off Madeira, off W Cape Finisterre, Azores, Uruguay (continental shelf), Brazil-Argentina, off Rio de Janeiro, Caribbean Sea, G. of Mexico, Florida, off Bermuda: Station "S" (32°10'N, 64°30'W), Sargasso Sea, Cape Hatteras, off Woods Hole, G. of St. Lawrence, off E Nova Scotia, Godthaab Fjord (W Greenland), Ungava Bay, Baffin Bay, Karajakfjord, Greenland Sea, Fram Strait, Wyville Thomson Ridge, Spitsbergen, Kongsfjorden, Newfoundland, Iceland, W Ireland, Norwegian Sea, Barents Sea, Franz Josef Land, North Sea, Arct. (all polar Basins, Fletcher's Ice Is., SE Beaufort Sea), Nansen Basin, Laptev Sea, Bering Sea, Canada Basin, Vancouver Is., Bay of Biscay, Ibero-moroccan Bay, Medit. (W Basin, Ligurian Sea, Ionian Sea, Alexandria), S Laccadive Is., Madagascar (Nosy Bé), S Indian (subtropical convergence), Indonesia-Malaysia, Philippines, China Seas, Japan (Izu), Izu-Bonin Trench, Mariana Trench, Hokkaido, Station Knot, Kuril-Kamchatka, S Aleutian Is., Bering Sea, S Aleutian Basin, Chukchi Sea, Canadian abyssal plain, Lomonosov Ridge, E Beaufort Sea, Hawaii, California, W Costa Rica, S Tasmania, ? New Zealand (Kaikoura), Chile (., N). | | | | N: | 159 | | | | Lg.: | | | (5) F: 3; M: 2,87; (7) F: 3,84-3,22; M: 3,01; (10) F: 3,25; (20) F: 3,68-3; M: 3,24-3; (22) F: 3,8-3,5; M: 2; (25) F: 3,51-3,02; M: 3,2-2,97; (29) F: 3,1; (31) F: 3,6-3,4; (35) F: 3,6-3,35; M: 3,25; (37) F: 3,8-3; M: 3,43-2; (39) F: 3,44; (41) F: 3,75-3,6; (45) F: 3,8-3,5; M: 2,25-2; (58) F: 3,2; (65) F: 3,8; M: 2; (102) F: 3,5-3,1; M: 3,1-2,9; (113) F: 3,65-3,3; ? (116) F: 2,92; (134) F: ± 4; (143) M: 3,1; (199) F: 3,12-2,81; M: 2,15-1,82; (201) ; M: 2,9; 2,7; (202) F: 3,5-3,8; M: 2; (203) F: 2,48; 2,02; M: 2,95; (246) F: 3,16-3,33; (254) F: 3; ? (1001) F: 3,59-6,81; {F: 2,02-4,00; M: 1,82-3,43} | | | | Rem.: | epi-bathypelagic. Characteristic in intermediate depths.
Sampling depth (Antarct., sub-Antarct.) : 100-1000+ m. Sargasso Sea (Station "S"): 500-2000 m. 369-616 m At Station T-1 (E Tori Is., E Japan).280-580 m (Pipe & Coombs, 1980, in Arctic).
Bradford & Jillett (1980, p.58, 61) consider a synonymy of this species with Gaidius pungens, a position not followed by Markhaseva (1996, p.217). For Park (1978, p.130) this species and G. pungens are closely similar but the metasomal processes are longer in the former than in the latter. In both female and male G. tenuispinus the first two exopodal segments of P1 are incompletely fused, and the endopod of P2 is 2-segmented. In the female the basis of the palp of Md has 2 setae. In the male the endopod of left P5 reaches about four fifths of the way to the distal end of the 1st exopodal segment. | | | Last update : 28/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 02, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
