|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Tortanidae ( Family ) |
|
|
|
Tortanus ( Genus ) |
|
|
|
Eutortanus ( Sub-Genus ) |
|
|
| |
Tortanus (Eutortanus) komachi Itoh, Ohtsuka & Sato, 2001 (F,M) | |
| | | | | | | Ref.: | | | Itoh & al., 2001 (p.60, figs.F,M); Andronov, 2014 (p.36, fig. A2) |  issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.61, Fig.1]. Female (from off S Akita): A-B, habitus (dorsal and lateral, respectively); C-D, prosomal end and urosome (dorsal and ventral, respectively); E-J, variability of prosomal end (dorsal view); K-P, idem (left lateral view); Q-V, idem (right lateral view); W, genital operculum.
|
 issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.62, Fig.2]. Female: A, A1; B, A2; C, Md; D, Md (mandibular cutting edge); E, Mx1 (minute setule indicated by arrow); F, Mx2; G, Mxp.
|
 issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.63, Fig.3]. Female: A-E, P1 to P5 (anterior surface).
|
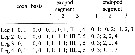 issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.60]. Female : Armament of the swimming legs P1 to P4. Roman numeral = spine; arabic numeral = seta.
|
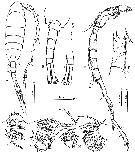 issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.64, Fig.4]. Male: A, habitus (dorsal); B, prosomal end and urosome (dorsal); C, 4th and 5th urosomites, anal somite, and caudal rami (ventral); D, right A1 (ventral view); E, right A1: segments XI-XV (dorsal view); F-I, variability of maxillular praecoxal arthrite (variable portion indicated by arrows).
|
 issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.65, Fig.5]. Male: A, P5 (anterior view); B, right P5 (posterior view).
|
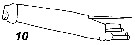 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.36, Fig.9, 10]. Tortanus komachi after Itoh & al., 2001. 10, exopod of A2.
| | | | | NZ: | 1 | | |
|
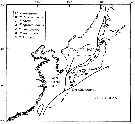
Distribution map of Tortanus (Eutortanus) komachi by geographical zones
|
| | |  issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.66, Fig.6]. issued from : H. Itoh, S. Ohtsuka & T. Sato in Plankton Biol. Ecol., 2001, 48 (1); [p.66, Fig.6].
horizontal distribution of species of the subgenus Tortanus (Eutortanus) around the Japan Sea, redrawn after Ohtsuka (1999); current system modified from Uda (1934).
In T. terminalis and T. komachi symbol indicates collected site, whereas in others the distributions are schematically exhibited.
The speciation of the six known species might have occured in the ancient East China Sea Gulf with low salinity water during the Middle Miocene to the Pleistocene, and relatively recently members of the subgenus may have secondarily expanded their distributions in the neighboring waters of the East China Sea. |
| | | | Loc: | | | Sea of Japan (NW Japan ) | | | | N: | 1 | | | | Lg.: | | | (923) F: 2,78-3,28; M: 2,43-2,50; {F: 2,78-3,28; M: 2,43-2,50} | | | | Rem.: | Found in the stomac contents of juvenile chum salmon (Onchorhynchus keta) and also found in plankton samples taken simultaneously when the salmon were captured. Depth: 20 m. Coast.
This species seems to be confined to the Sea of Japan and neighboring waters influenced by the Tsushima Current. Hirakawa (1984) reported a species as T. (E.) derjugini? from Funka Bay and the morphological characters of the species are very similar to those of T. (E.) komachi (Hirakawa, pers. comm.). The new species has several advanced characters compared with T. (E.) derjugini: 1- 3rd urosomite and caudal rami of the female are completely fused (incompletely in derjugini); 2- the right endopod of male P5, represented by a remarkably inward produced process, is similar in shape to that of the 5th copepodid stage of derjugini (neoteny).
These morphological characters suggest that the new species might have been split off from derjugini after the species was introduced into the Sea of Japan in the late Pleistocene. The speciation of the new species is suggested to have occured since 20.000-60.000 years ago
In the Calanoida speciation rates are estimated to fall between 10.000-1.000.000 years (Fleminger, 1986). The speciation rates of the subgeneric members seem to be relatively fast (cf. Nishimura, 1980). | | | Last update : 18/04/2016 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 21, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




 komachi - Plate 1 of morphological figures',750,500);)
 komachi - Plate 2 of morphological figures',750,500);)
 komachi - Plate 3 of morphological figures',750,500);)
 komachi - Plate 5 of morphological figures',750,500);)
 komachi - Plate 6 of morphological figures',750,500);)
 komachi - Distribution map 2',750,500);)
 komachi - Plate 1 of morphological figures){kind=link}
 komachi - Plate 2 of morphological figures){kind=link}
 komachi - Plate 3 of morphological figures){kind=link}
 komachi - Plate 4 of morphological figures){kind=link}
 komachi - Plate 5 of morphological figures){kind=link}
 komachi - Plate 6 of morphological figures){kind=link}
 komachi - Plate 7 of morphological figures){kind=link}
 komachi - Distribution map 2){kind=link}
