|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Pseudodiaptomidae ( Family ) |
|
|
|
Calanipeda ( Genus ) |
|
|
| |
Calanipeda aquaedulcis Kritschagin, 1873 (F,M) | |
| | | | | | | Syn.: | Poppella guernei Richard, 1888 (p.43); de Guerne & Richard, 1889 (p.149, figs.); Giesbrecht & Schmeil, 1898 (p.63, fig.M) | | | | Ref.: | | | Dussart, 1967 a (p.86, figs.F,M); Kiefer, 1978 d (p.57, figs.F,M); Grindley, 1984 (p.218, fig.M, Rem.); Dussart, 1989 (p.13, figs.F,M); Boxshall & Halsey, 2004 (p.174) | 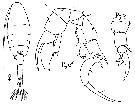 issued from : B. Dussart in Ls Copépodes des eaux continentales. Edit. N. Boubée & Cie, 1967, Tome I: Calanoïdes et Harpacticoïdes. [p.87, Fig.22]. Female (from Camargue, France): habitus (dorsal); P5. Nota: A1 25-segmented (25th very little), reaching about the 2nd urosomal segment. Urosome 4-segmented. Genital segment slightly shorter than the urosomal segments 2 and 3 together; showing a hook, curved backwards with a sensitive spine. P5 without endopodite; exopodite 3-segmentedexopoal segment 2 with 1 inner, a process distal spiniform strong spine, and at base 1 little, strong, spine; 3rd exopodal segment a thick claw-like, curved inward, with 1 strong inner spine and a little, short process at base. After Schachter (1952) there is a slightly dissymetry in exopodite segment 2 between the right and left legs. Caudal rami 6 -7 times longer than wide, inner marginal frange, with 5 plumose setae. Male: P5. Nota: Right A1 prehensile.
| | | | | Compl. Ref.: | | | Rose & Vaissière, 1952 a (p.121); Cannicci, 1962 (p.349, Table 3); Castel & Lasserre, 1977 (p.129, fig.8); Grindley, 1990 (p.240, Rem.); Cavallo & Belmonte, 1995 (p.56, tab.1); Mouelhi & al., 1999 (p.61, Rem.: p.68); Azeiteiro & al., 1999 (p.23, annual cycle); Bressan & Moro, 2002 (p.58, tab.2); Kovalev, 2003 (p.47); Vieira & al., 2003 (p.S163, Table 2, abundance); S.C. Marques & al., 2006 (p.297, tab.III); S.C. Marques & al., 2007 (p.213, fig.6); Marques S.C. & al., 2007 (p.725, Table 1, fig.4, climate variability); S.C. Marques & al., 2011 (p.59, Table 1); Svetlichny & al., 2012 (p.15, mortality, respiration egg mass density vs salinity); Uriarte & al., 2016 (p.718, fig.5: abundance vs years 1998-2011) ; Svetlichny & al., 2016 (p.77, fig.8: respiration rate, Rem.); Belmonte, 2018 (p.273, Table I: Italian zones) | | | | NZ: | 2 | | |
|
Distribution map of Calanipeda aquaedulcis by geographical zones
|
| | | | | |  Issued from : I. Uriarte, F. Villate & A. Iriarte in J. Plankton Res., 2016, 38 (3). [p.724]. Issued from : I. Uriarte, F. Villate & A. Iriarte in J. Plankton Res., 2016, 38 (3). [p.724].
Monthly time-series from 1998 to 2011 of Acartia tonsa (black) and PCPCalanus (mostly Calanipeda aquaedulcis (grey) in the inner estuary of Bilbao (NW Spain). Thicker lines are moving averages. Ro is the Spearman's rank correlation coefficient (***P<0.001).
Nota: Calanipeda aquaedulcis showed positive correlation with temperature and Secchi dissk depths and negative correlation with streamflow.
After Uriarte & al. (2016, p.728) this copepod contribues significantly to the increase in the abundance of total copepods in the estuary of Bilbao (NW Spain) in the last period of the time series. Despite it being often considered as a marine species, it has been observed from freshwater to salt lake systems, being an extremely euryhaline osmoconformer (see Svetlichny & al., 2012). The species is commonly found in the upper reaches of other nearby Basque coast estuaries such as Plentzia., but it was first observed in the estuary of Bilbao in 2001 (see Albaina & al., 2009). The species showede a very large increase in abundance during the last 2 years of the time-series (2010-2011), linked promarily to an increase in water transparency, and under conditions of lowered chlorophyll a concentrations and of certain stability in the improved levels of DOS (dissolved oxygen saturation). It is considered as eurythermal, but usually with a preference for cooler temperature (see Frisch & al., 2006). Accordingly, in the estuary of Bilbao, it was found that it could attain high abundance already by March, although peak abundaces could be attained in July. |
| | | | Loc: | | | Portugal (Mondego estuary); Bilbao estuary; Canal du Midi (France), Salse & Canet lagoons, around mediterranean Sea, Tunisia, Sicily, , Sardinia, Apulia, Appenine, Po Valley, Adriatic (S, N), Black Sea (estuaries), Azov Sea, Caspian Sea, Aral Sea | | | | N: | 22, Data incomplete | | | | Lg.: | | | F: 1,30-1,45; M: 1,0 (from Camargue); {F: 1,30-1,45; M: 1,0} | | | | Rem.: | freshwater species, sometimes in river mouths and brackish lagoons.
Not considered in the marine environment.
Incomplete data.
Diameter of subitaneous egg: 108 ±7 µm.; mass density: 1.074 ± 0.009; for salinity 18.2 psu (after Svetlichny & al., 2012) | | | Last update : 20/10/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed January 02, 2026] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)

;)
{kind=link}
{kind=link}
