|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Pseudodiaptomidae ( Family ) |
|
|
|
Pseudodiaptomus ( Genus ) |
|
|
| |
Pseudodiaptomus pankajus Madhupratap & Haridas, 1992 (F,M) | |
| | | | | | | Ref.: | | | Madhupratap & Haridas, 1992 (p.555, figs.F,M) | 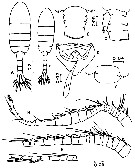 Issued from : M. Madhupratap & P. Harida in J. Plankton Res., 1992, 14 (4). [p.556, Fig.1].
Female (from Gulf of Kutch): A, habitus (dorsal); B, urosomite 1 (dorsal); C, urosomite 1 (left lateral view); D, ventral genital field with spermatophore; E, A1.
Male: F, habitus (dorsal); G, urosomite 1 (dorsal); H, right A1.
Scale bars in mm.
Female:
- Head and 1st pediger separate, 4th and 5th fused.
- Posterior corners of metasome rounded with 1 pair of posteriorly directed dorsal spines and spinules between spines and urosome insertion.
- Urosomite 1 symmetrical with 3 rows of lateral spinules extending incompletely dorsal terminating in a seta. Genital field with a row of proximal spinules.
- Urosomites 1-3 with posterodorsal spine rows (spines on Ur3 largest).
- Caudal rami 3 times longer than wide with 5 setae and a dorsal setule; middle seta slightly swollen.
- Urosomites segments and caudal rami with proportions 29 : 17 : 16 : 15 : 23 = 100.
- A1 22 segmented with modified barbed seta on segment 20.
Male:
- Prosome - urosome ratio: 1.6 : 1.
- Posterior corners of metasome with 1 pair of small dorsal spines.
- Urosomite 1 with incomplmete dorsal spinule row at midlength.
- Urosomites 2-4 with postrodorsal spinule rows.
- Caudal rami 3 times longer than wide with setulation similar to female except that middle seta is not swollen.
- Urosme segments and caudal rami with proportions 12 : 19 : 17 : 17 : 15 : 20 = 100.
- Right geniculate A1 21-segmented; geniculation between segments 17 and 19 with flanges; segment 18 with comb plate of spinules.
- Other mouth parts and P1-P4 similar to female.
|
 Issued from : M. Madhupratap & P. Harida in J. Plankton Res., 1992, 14 (4). [p.557, Fig.2].
Female: A, A2; B, Md; C, tip of the gnathal lobe of Md; D, Mx1; E, Mx2; F, Mxp.
Scale bars in mm.
Nota:
- Gnathal lobe of Md with about 16 teeth and seta.
- 4 modified bristles present on inner lobes of Mxp.
|
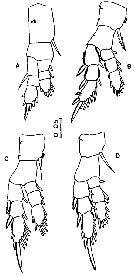 Issued from : M. Madhupratap & P. Harida in J. Plankton Res., 1992, 14 (4). [p.558, Fig.3].
Female : A, P1; B, P2; C, P3; D, P4.
Scale bar in mm.
|
 Issued from : M. Madhupratap & P. Harida in J. Plankton Res., 1992, 14 (4). [p.559, Fig.4 A].
Female; A, P5 (anterior view).
Nota: P5 asymmetrical. coxa (B1) with proximal spinule row, basis (B2) with large posterior surface seta, very fine proximomedial spinule patch and a distal spinule row continuing on to lateral surface. Left basis (B2) extending over proximal corner of exopodal segment 1 with large bulbous ptojection at distomedial corner; right basis (B2) also extending over exopodal segment 1 with irregular medial margin. Exopodal segment 1 wit naked seta, exopodal segment 2 with stout medial spiriform process, both margins hirsute and small naked seta. Exopodal segment 3 with proximomedial spiniform process slightly lo,nger than exopodal segment 2 spiniform ptocess and appears divided distal to origin of basal process.
|
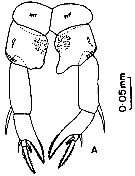
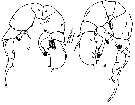 Issued from : M. Madhupratap & P. Harida in J. Plankton Res., 1992, 14 (4). [p.559, Fig.4].
Male: B, P5 (posterior view); C, P5 (anterior view).
Scale bar in mm.
Nota: P5 posterior view : Coxa (B1) distomedial corners acute. Right leg -, basis (B2) with 1 large surface seta at midlength, 2 distal setae, rows of proximomedial spinules and a row of dorsolateral spinules. Endopodite consists of 2 processes, both oroiginating from the same base at the disyomedial corner of basis and with a hyaline process having an apical cleft in the middle. Medial process of endopodal spinifotrm with seta at the tip, lateral process stout, apex bulbous with irregular margin and a corona of small spinules. Exopodal segment 1 roughly triangular with a circular cluster of spinules at midlength, sculptured with 3 distolateral small ridges, 2 surface setae and distolateral corner ending in a small spine like process; outer seta long, recurved, reaching 3/4 of exopodal segment 2 with proximopmedial margin heavily serrate. Exopodal segment 2 elongate with distomedial knob, surface seta at midlength and outer seta 3/4 the length of segment. Exopodal segment 3 proximomedially swollen with spinules and 2 setae and tip finely serrate.
Left leg basis (B2) with large surface seta, seta on medial margin and a distolateral row of spinules. Large recurved endopod originating from distomedial corner of basis, extending past midlength of exopodal segment 2, base swollen, apex with distomedial spinules and ending in a circular row of long spines. Exopodal segment 1 simple with naked outer seta. Exopodal segment 2 large, flattened and hemispherical with long outer seta, 3 surface setae and 1 terminal seta, medial margin with 3 setae.
Anterior view: coxa (B1) with spinule row. Basis (B2) with row of spinules at midlength continuing on to the lateral sides and distal spinule row. Right exopodal segment 2 with fine distal spinules; left coxa (B1) with proximomedial very fine spinules; left exopodal segment 2 with proximomedial hairy patch extending distally along ridge.
The specific name, pankajus, refers to the lotus-like arrangement of clustered spinules at tips of both endopods and on right exupodal segment 1 (pankaj = lotus, origin-Sanskrit, gender: male).
| | | | | NZ: | 1 | | |
|
Distribution map of Pseudodiaptomus pankajus by geographical zones
|
| | | | | | | Loc: | | | NW India (Singach near Gulf of Kutch: 69°30' N, 22°20' N, 69°30' E) | | | | N: | 1 | | | | Lg.: | | | (1318) F: 1,17-1,27; M: 1,01-1,09; {F: 1,17-1,27; M: 1,01-1,09} | | | | Rem.: | Hyperhalin (salinity between 70-90 p.1000) from shallow water (2-4 m depth); It did not occur in salt ponds with salinity > 90 p.1000.
Incomplete data.
This new species is assignable to the Improcerus species group, after Walter & al. 2006, p.203; based on the presence of processes on the distomedial corners of female P5 basis; however, the right endopod of male P5 is not simple.
For Madhupratap & Haridas (1992, p.560) the species is closely related to P. stuhlmanni (Poppe & Mrazek, 1895). The occurrence of P. pankajus , so far away from its siblings (about 3000-3500 nautical miles) inhabiting waters around Madagascar and southern Africa may have important palaeontological significance. Sewell (1956) attempted to explain the distribution of copepods, particularly pseudodiaptomids, based on continental drift theory and hypothesized that modern species existed right back to the Cretaceous. Grindley (1969) suggested that Pseudodiaptomidae originated in the late Tertiary and speciated and dispersal within the Quaternary; the author (1984) feels that the ancestors of modern Pseudodiaptomidar spread through the Tethys Sea during the Cretaceous or early Tertiary;It seems appropiate to invoke the pre-drift position of the continents to explain the origin of these distancy located, but closely related, species.. It is well-known that the land masses of India and Africa were joined together to form the Gondwanaland (Smith, 1980). The India started sundering apart from the Gondwana in the mid-Triasic and continued its northward drift during the Jurassic until it encounterd the southern margin of Asia during the early Cenozoic (Tertiary). Madagascar seems to have separated from India/Africa during the Cretaceous before drifting to its present position (Krishnan, 1968; Dietz & Holden, 1970). Thus it is very probable that P. pankajus evolved as an allopatric species as a result of these earlier epochs. Such distributions of estuarine or neritic species are difficult to explain based on present current circulation (see Fleminger & Kramer, 1988); except the recent development of the boats and the alien species introduced. | | | Last update : 22/06/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 11, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
