|
|
 |
|
Monstrilloida ( Order ) |
|
|
|
Monstrillidae ( Family ) |
|
|
|
Monstrillopsis ( Genus ) |
|
|
| |
Monstrillopsis cahuitae Suarez-Morales & Carillo, 2013 (M) | |
| | | | | | | Ref.: | | | Suarez-Morales & Carillo, 2013 (p.626, Descr.M, figs.M, Rem.) | 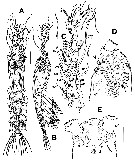 Issued from : E. Suarez-Morales, A. Carillo & A. Morales-Ramirez in J. Nat. Hist., 2013, 47 (5-12). [p.627, Fig.4]. Male (from Cahuita, Costa Rica): A-B, habitus (dorsal and lateral, respectively); C, right A1 (dorsal view); D, cephalic area (lateral), showing position of oral papilla and ventral protuberance (arrowed); E, same (ventral), showing paired nipple-like processes (arrowed). Scale bars: 100 µm (A, B); 50 µm (C-E). Nota: Small species. Cephalothorax representing up to 48.6% of total body length. Oral papilla at 20% of way back along ventral surface of cephalothorax. A1 5-segmented, geniculate, representing 92% of the cephalothorax length. Pair of relatively large pigment cups present, moderately developed, separated by less than half length of eye diameter, weakly pigmented; ventral cup larger than lateral cups. 1st pedigerous somite incorporated into cephalothorax. Urosome 5-segmented: P5 absent Genital apparatus cylindrical, with paired globose processes distally. Caudal rami subrectangular, weakly divergent, about 1.6 times longer than wide, each caudal ramus with 6 setae; inner dorsal seta (seta VII in Huys & Boxshall, 1991) thinnest.
|
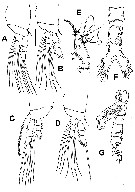 Issued from : E. Suarez-Morales, A. Carillo & A. Morales-Ramirez in J. Nat. Hist., 2013, 47 (5-12). [p.628, Fig.5]. Male: A, P1 (dorsal); B, P2 (dorsal); C, P3 (ventral); D, P4 (dorsal); E, 5th antennular segment showing distinctive inner process (arrowed); F, urosome (ventral), caudal setae cut short; G, urosome (lateral), caudal seta cut short. Scale bars: 50 µm (A-D, F, G); 25 µm (E).
|
 Issued from : E. Suarez-Morales, A. Carillo & A. Morales-Ramirez in J. Nat. Hist., 2013, 47 (5-12). [p.629, Table 2]. Male: Formula armature of swimming legs P1 to P4. Spines denoted by Roman numerals, setae by Arabic numerals from outer margin of any segment given first, separated by a hyphen from the inner margin.
| | | | | NZ: | 1 | | |
|
Distribution map of Monstrillopsis cahuitae by geographical zones
|
| | | | Loc: | | | Caribbean Sea (Cahuita, SE Costa Rica reef) | | | | N: | 1 | | | | Lg.: | | | (1120)* M: 0,55
*: caudal rami excluded. | | | | Rem.: | For Suarez-Morales & al (2013, p. 629) the Cahuitas male specimen is assignable to the genus Monstrillopsis by its possession of a modified 5th antennular segment, with an inner rounded protuberance and an attenuated, sabre-like distal half ; this kind of modification occurs only in males of Monstrillopsis (see Huys & Boxshall, 1991 ; Suarez-Morales a al., 2006). The position of the oral papilla near the anteriormost part of the cephalothorax and fully developed eyes present ;; both characters are regarded as diagnostic of the genus (Sars, 1921 ; Suarez-Morales & al., 2006). The presence of 6 caudal setae in Cahuitas specimen is quite unexpected ; this is usually regarded as a character of the genus Monstrilla and having 4 such setae is associated with Monstrillopsis (Sars, 1921 ; Suarez-Morales & Ivanenko, 2004 ; Suarez-Morales & al., 2006). In this case i twas not considred definitive because the number of caudal setae appears to be more consistent in females than in males ; for instance, both the female and the male of Monstrillopsis zernovi bear 5 caudal setae (Dolgopolskaya, 1948) and represent aberrant cases among the members of this genus (Suarez-Morales & al., 2006).
The decision to assign this male to Monstrillopsis is supported by the strongest character for males of this genus : the unique morphology of the last antennular segment (as described by Huys & Boxshall, 1991). This unique combination of characters is not present in any other known congener of Monstrillopsis and so the specimen from Costa Rica is not present in any other known congener of Monstrillopsis and so the specimen is recognized as a new species.
Another striking character of this species is its small size (body : 0.55 mm long) ; it is among the smallest monstrilloids known. Only Monstrilla pygmaea Suarez-Morales, 2000 (0.43 mm) and Monstrilla minuta Isaac, 1975 (0.49 mm) are smaller. Besides the antennular modification another intresting character shared by males of Monstrilla and Monstrillopsis is the presence of a long, cylindrical genital apparatus with paired terminal globose processes. This kind of structure has been recorded mainly in species of Monstrilla (i.e. Monstrilla reidae, Monstrilla globosa, Monstrilla bahiana, Monstrilla patagonica). It is not exclusive of the genus because it has also been described in species of Monstrillopsis (i.e. Monstrillopsis zernovi, Monstrillopsis fosshageni, Monstrillopsis chathamensis). A different kind of genital apparatus reported in species of Monstrillopsis is that described in Monstrillopsis chilensis with long genital lappets.
The Costa Rica specimen is comparable to Monstrillopsis fosshageni, but differs in being much smaller (0.55 mm versus 1.8 mm), having a terminal spiniform process of the 5th antennular segment less than half the length of the segment versus a longer element in M. fosshageni, representing more than half the length of the segment. The genital apparatus has the same structure and proportions, but the terminal lappets are relatively stronger in the new species ; also, in Monstrillopsis fosshageni, these are smooth, without the blister-like ornamentations described in Monstrillopsis cahuitae.
After Lee J. & al; (2016, p.421), genitalia male with a well developed, long median shaft and relatively short, ovoidal or globose lateral lappets that are more or less separated from the median shaft belongs subgroup ''Type I'' (Suarez-Morales & McKinnon, 2014). | | | Last update : 07/12/2017 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 25, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
{kind=link}
{kind=link}
{kind=link}
