 List of the studied authors
List of the studied authors
Sewell, 1948 (p.322); Yamazi, 1958 (p.153, Rem.); Hanaoka, 1977 (p.267, 301, abundance); Chen Q-c, 1980 (p.794); Greenwood, 1981 (p.K591, fig.1: co-occurrence); Hirota 1981 (p.19, Table 1, length-weight-CHN); Uye & al., 1984 (p.390, resting eggs-survival); Madhupratap & Haridas, 1986 (p.105, tab.2); Othman & al., 1990 (p.561, 564, Table 1); Yoo, 1991 (tab.1); Dai & al., 1991 (tab.1); Hattori, 1991 (tab.1, Appendix); Yoo & al., 1991 (p.259); Myung & al., 1994 (tab.1); Shih & Young, 1995 (p.66); Ohno & al., 1996 (p.231, fig.1); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Park & Choi, 1997 (Appendix); Ramaiah & Nair, 1997 (tab.1); McKinnon & Klumpp, 1998 (tab.1); Mauchline, 1998 (tab.40); Hsieh & Chiu, 1998 (tab.2); Noda & al., 1998 (p.55, Table 3, occurrence); Nair & Ramaiah, 1998 (p.274); Achuthankutty & al., 1998 (p.1, Table 2, fig.5, seasonal abundance vs monsoon); Dolganova & al., 1999 (p.13, tab.1); Hsiao & al., 2004 (p.325, tab.1); Rezai & al., 2004 (p.486, tab.2, 3, abundance); Chang & Fang, 2004 (p.456, tab.1); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Lan & al., 2004 (p.332, tab.1); Lo & al., 2004 (p.89, tab.1); Jiang X. & al., 2004 (p.89, resting eggs vs sediment depth); Choi & al., 2005 (p.710: Tab.III); Rezai & al., 2005 (p.157, Table 2: spatial & temporal variations); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: stations group); Hwang & al., 2006 (p.943, tab.I); Dur & al., 2007 (p.197, Table IV); Madhu & al., 2007 (p.54, Table 4, abundance vs monsoon); Jitlang & al., 2008 (p.65, Table 1); McKinnon & al., 2008 (p.843: Table. I, p.848: Tab. IV); Ohtsuka & al., 2008 (p.115, Table 5); Moon & al., 2008 (p.61); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Lan & al., 2009 (p.1, Table 2, % vs hydrographiv conditions); Jiang Z.-B. & al., 2009 (p.196, Table 1, 2); Zhang G.-T. & al., 2010 (p.492, Table 1, 2); Cornils & al., 2010 (p.2076, Table 3); Zengling & al., 2010 (p.233, UV-radiation response); Sun & al., 2010 (p.1006, Table 2); Fazeli & al., 2010 (p.153, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I); Guo & al., 2011 (p.597, Table 2, fig.9, indicator, abundance); Maiphae & Sa-ardrit, 2011 (p.641, Table 2); Sun X.-H. & al., 2011 (p.741, figs. 5, 3, 4, tab.I, II, production); Zhang D. & al., 2011 (p.86, pH effects); Hsiao & al., 2011 (p.232, Table 1: abundance, %, Table 2: trace metal concentration); Beltrao & al., 2011 (p.47, Table 1, density vs time); Kang J.-H., 2011 (p.219, occurrences, inter-annual variability vs t° & Sal., Chl.a, Rem.: p.224); Hsiao & al., 2011 (p.317, Table 2, indicator of seasonal change); Johan & al., 2012 (2013) (p.1, Table 1); Mulyadi & Rumengan, 2012 (p.202, Rem.: p.204); Yoshida & al., 2012 (p.644, fig.1, 3, Table 1, egg development time and hatching vs temperature); Lee D.B. & al., 2012 (p.88, co-occurrence, abundance, feeding); Huo & al., 2012 (p.1, Table 1: dominance); Yoshida & al., 2012 (p.251, feeding, egg production, Table 2: other data); Lee J. & al., 2012 (p.95, Table 2: abundance, annual & seasonal distribution); Zhang H. & al., 2013 (p.57, Rem. p.58); Suzuki, K.W. & al., 2013 (p.15, Table 2, 3, 4, estuaries, annual occurrence); Seo & al., 2013 (p.448, Table 1, occurrence); Oh H-J. & al., 2013 (p.192, Table 1, occurrence); Nakajima & al., 2013 (p.27, cooccurence); Nakajima & al., 2015 (p.19, Table 3: abundance); Trottet & al., 2017 (p. 7, Table 3: resting stage); Ohtsuka & Nishida, 2017 (p.565, Table 22.1)


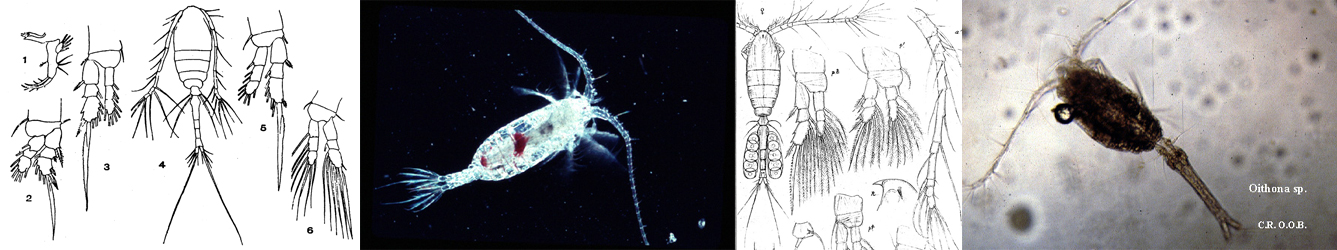

 pacifica - Plate 1 of morphological figures',750,500);)
 pacifica - Plate 2 of morphological figures',750,500);)
 pacifica - Plate 3 of morphological figures',750,500);)
 pacifica - Plate 4 of morphological figures',750,500);)
 pacifica - Plate 5 of morphological figures',750,500);)
 pacifica - Plate 6 of morphological figures',750,500);)
 pacifica - Plate 7 of morphological figures',750,500);)
 pacifica - Plate 8 of morphological figures',750,500);)
 pacifica - Plate 9 of morphological figures',750,500);)
 pacifica - Plate 10 of morphological figures',750,500);)
 pacifica - Plate 11 of morphological figures',750,500);)
 pacifica - Plate 12 of morphological figures',750,500);)
 pacifica - Plate 13 of morphological figures',750,500);)
 pacifica - Plate 15 of morphological figures',750,500);)

 pacifica - Distribution map 4',650,500);)
 pacifica - Distribution map 6',750,500);)
 pacifica - Distribution map 7',750,500);)
 pacifica - Distribution map 8',750,500);)
 pacifica - Distribution map 9',750,500);)
 pacifica - Distribution map 10',750,500);)
 pacifica - Distribution map 11',750,500);)
 pacifica - Distribution map 12',750,500);)
 pacifica - Distribution map 13',750,500);)
 pacifica - Plate 1 of morphological figures){kind=link}
 pacifica - Plate 2 of morphological figures){kind=link}
 pacifica - Plate 3 of morphological figures){kind=link}
 pacifica - Plate 4 of morphological figures){kind=link}
 pacifica - Plate 5 of morphological figures){kind=link}
 pacifica - Plate 6 of morphological figures){kind=link}
 pacifica - Plate 7 of morphological figures){kind=link}
 pacifica - Plate 8 of morphological figures){kind=link}
 pacifica - Plate 9 of morphological figures){kind=link}
 pacifica - Plate 10 of morphological figures){kind=link}
 pacifica - Plate 11 of morphological figures){kind=link}
 pacifica - Plate 12 of morphological figures){kind=link}
 pacifica - Plate 13 of morphological figures){kind=link}
 pacifica - Plate 14 of morphological figures){kind=link}
 pacifica - Plate 15 of morphological figures){kind=link}
 pacifica - Distribution map 4){kind=link}
 pacifica - Distribution map 5){kind=link}
 pacifica - Distribution map 6){kind=link}
 pacifica - Distribution map 7){kind=link}
 pacifica - Distribution map 8){kind=link}
 pacifica - Distribution map 9){kind=link}
 pacifica - Distribution map 10){kind=link}
 pacifica - Distribution map 11){kind=link}
 pacifica - Distribution map 12){kind=link}
 pacifica - Distribution map 13){kind=link}