|
|
 |
|
Calanoida ( Order ) |
|
|
|
Calanoidea ( Superfamily ) |
|
|
|
Calanidae ( Family ) |
|
|
|
Mesocalanus ( Genus ) |
|
|
| |
Mesocalanus tenuicornis (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Calanus tenuicornis Dana,1849; Giesbrecht, 1892 (p.90, 129, figs.F,M); Giesbrecht & Schmeil, 1898 (p.18); Cleve, 1904 a (p.186); Esterly, 1905 (p.127, figs.M, Rem.F,M); Pearson, 1906 (p.5); Farran, 1908 b (p.20); A. Scott, 1909 (p.8); Sewell, 1914 a (p.193); Pesta, 1920 (p.496); Farran, 1926 (p.227); 1929 (p.207, 217); Sewell, 1929 (p.20); Mori, 1929 (p.170, Figs.F,M); Rose, 1929 (p.8); 1933 a (p.62, figs.F,M); Dakin & Colefax, 1933 (p.204); Farran, 1936 a (p.77); Mori, 1937 (1964) (p.16, figs.F,M); Dakin & Colefax, 1940 (p.87, figs.F,M); Vervoort, 1946 (p.22, Rem.); Sewell, 1948 (p.347, 513, 519, 544, 555, 565); Brodsky, 1950 (1967) (p.96, figs.F,M); C.B. Wilson, 1950 (p.269); Vervoort, 1951 (n°32, p.3, figs.F,M); Bowman, 1955 (p.413, figs.F,M, Rem.); Tanaka, 1956 (p.255); Chiba & al., 1957 (p.306); 1957 a (p.11); Yamazi, 1958 (p.146); Gaudy, 1962 (p.93, 99, Rem.: p.101); Grice & Hart, 1962 (p.287, table 3); Grice, 1962 (p.175, Rem.); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); V.N. Greze, 1963 a (tabl.2); Shmeleva, 1963 (p.141); Björnberg, 1963 (p.10, Rem.); Kasturirangan, 1963 (p.10, figs.F,M); Unterüberbacher, 1964 (p.13); De Decker, 1964 (p.14, 18, 29); De Decker & Mombeck, 1964 (p.11); Grice & Hulsemann, 1965 (p.223); Chen & Zhang, 1965 (p.28, figs.F,M); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Pavlova, 1966 (p.43); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio/kuroshio transitional area, Table 7, 10); Fleminger, 1967 a (tabl.1); Ehrhardt, 1967 (p.737, 887, figs.51-53, geographic distribution), Rem.); Grice & Hulsemann, 1967 (p.13); Delalo, 1968 (p.137); Vilela, 1968 (p.7, fig.M); Vidal, 1968 (p.14, figs.F); De Decker, 1968 (p.45); Park, 1968 (p.532, Rem.); Mullin, 1969 (p.438, biogéo); Park, 1970 (p.474); Morris, 1970 (p.2300); Corral Estrada, 1970 (p.64, figs.F,M); Dowidar & El-Maghraby, 1970 (p.267); Deevey, 1971 (p.224); Marshall & Orr, 1972 (p.8); Brodsky, 1972 (1975) (p.9, 69, 85, 121, figs.); Bradford, 1972 (p.31, fig.F); Razouls, 1972 (p.91, Tableau XXVI, XXVII, Annexe: p.5, figs.F,M); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night); Boucher & Thiriot, 1972 (p.47, Tableau 4); Grice, 1972 (p.175); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 a (p.318); Heinrich, 1973 (p.95); Björnberg, 1973 (p.287, 384); Ikeda, 1974 (p.1, respiration ammonia excretion rate); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Harding, 1974 (p.141, tab.3, gut contents); Peterson & Miller, 1975 (p.642, 650, Table 3, interannual abundance); Vives & al., 1975 (p.35, tab.II, III, IV); Peterson & Miller, 1976 (p.14, Table 1, 2, 3, abundance vs interannual variations); Vyshkvartzeva, 1976 (p.14); 1977 a (p.97, figs.); Carter, 1977 (1978) (p.35); Peterson & Miller, 1977 (p.717, Table 1, seasonal occurrence); Deevey & Brooks, 1977 (p.256, Table 2, Station "S"); Dawson & Knatz, 1980 (p.5, F,M); Sreekumaran Nair & al., 1981 (p.493), Björnberg & al., 1981 (p.616, figs.F,M); Brenning, 1981 (p.1, Rem.: p.4); Kovalev & Shmeleva, 1982 (p.82); Alvarez-Marques, 1984 (p.112, figs.F,M, annual abundance); De Decker, 1984 (p.315, 332: carte); Scotto di Carlo & al., 1984 (p.1042); Cummings, 1984 (p.163, Table 2); Brenning, 1985 a (p.3, Table 2); Regner, 1985 (p.11, Rem.: p.22); Brinton & al., 1986 (p.228, Table 1); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Lozano Soldevilla & al., 1988 (p.57); Jimenez-Perez & Lara-Lara, 1988; Hernandez-Trujillo, 1989 a (tab.1); Suarez & al., 1990 (tab.2); Valdes & al., 1990 (tab.2); Scotto di Carlo & al., 1991 (p.271); Yoo, 1991 (tab.1); Suarez & Gasca, 1991 (tab.2); Morales C.E. & al., 1991 (p.455, Table I, grazing); Rosales & Sepulveda, 1992 (p.38); Ayukai & Hattori, 1992 (p.163, Table 5, fecal pellet production rate); Suarez, 1992 (App.1); Ashjian & Wishner, 1993 (p.483, abundance, species group distributions); Morales C.E. & al., 1993 (p.185); Seguin & al., 1993 (p.23, Table 2: abundance, %); ); Landry & al., 1994 (p.55, abundance, grazing); Hays & al., 1994 (tab.1); Suarez-Morales, 1998 (p.345, Table 1); Harvey & al., 1999 (p.1, 49: Appendix 5, in ballast water vessel); Gomez-Gutiérrez & Peterson, 1999 (p.637, Table II, abundance); Moraitou-Apostolopoulou & al., 2000 (tab.I); Sautour & al., 2000 (p.531, Table II, abundance); Weikert & al., 2001 (p.227, tab.4, Rem.); Pakhomov & al., 2000 (p.1663, Table 2, transect Cape Town-SANAE antarctic base); Hernandez-Trujillo & Esqueda-Escarcega, 2002 (in Appendix); Vukanic, 2003 (p.139, tab.1); Vukanic & Vukanic, 2004 (p.9, tab. 2, 3); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Maiphae & Sa-ardrit, 2011 (p.641, Table 2); Sobrinho-Gonçalves & al., 2013 (p.713, Table 2, fig.8, seasonal abundance vs environmental conditions);
Neocalanus tenuicornis : Sars, 1925 (p.9); Wilson, 1942 a (p.195); Lysholm & al., 1945 (p.7); C.B. Wilson, 1950 (p.269); Brodsky, 1962 c (p.98); Fagetti, 1962 (p.7); Chen & Zhang, 1965 (p.28, figs.F,M); Mazza, 1966 (p.69); Brodsky, 1972 (1975) (p.116); Vives & al., 1975 (p.35, tab.II); Zheng & al., 1982 (p.7, figs.F,M); Brodsky & al., 1983 (p.185, figs.F,M); Guangshan & Honglin, 1984 (p.118, tab.); Kouwenberg, 1994 (tab.1); Go & al., 1997 (tab.1); Park & Choi, 1997 (Appendix); Hure & Krsinic, 1998 (p.15, 99); Padmavati & al., 1998 (p.349); Hwang & al., 1998 (tab.II); Mauchline, 1998 (tab.30); Bragina, 1999 (p.195); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Ashjian & al., 2005 (p.1380: tab.2); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Pagano, 2009 (p.115);
Neocalanus tenuicornis : Noda & al., 1998 (p.55, Table 3, occurrence); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence) | | | | Ref.: | | | Bradford & Jillett, 1974 (p.12, figs.F,M); Gardner & Szabo, 1982 (p.148, figs.F,M); Mackas & Sefton, 1982 (p.1179); Fleminger, 1985 (p.276, 285, Table 1, 4, fig.M, Rem.: A1); Bradford,1988 (p.74, 76); Nishida, 1989 (p.173, fig. 3, table 1, 2, 3: dorsal hump); Bradford-Grieve, 1994 (p.38, figs.F,M, Rem., fig.98); Chihara & Murano, 1997 (p.741, Pl.69: F,M); Bradford-Grieve & al., 1999 (p.877, 907, figs.F,M); Bucklin & al., 2003 (p.335, tab.2, fig. 4, Biomol.); Conway & al., 2003 (p.147, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.81: fig.M); Avancini & al., 2006 (p.60, Pl. 29, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.898, figs.F,M, Rem.); Blanco-Bercial & al., 2014 (p.5: Rem.: problematical for barcoding) | 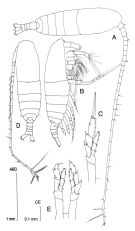 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). New Zealand Oceanographic Institute Memoir, 102, 1994. [p.39, Fig.14]. Female: A, habitus (dorsal); B, idem (right lateral side); C, P5. Nota: Li 1 of Mx2 with 4 setae. Endopodite 1 of P1 without inner seta. Endopodite of P5 with 7 setae. Male: D, habitus (dorsal); E, P5. Nota: Left P5 slightly modified, not prehensile. Exopodite 1 of P2 without recurved spine (hook) adjacent to outer spine. Coxa of P5 with inner edge without teeth. Both endopodite of P5 with 7 setae each.
|
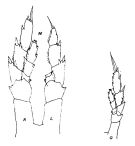 issued from : J.M. Bradford & J.B. Jillett in Crustaceana, 1974, 27 (1). [p.14, Fig.2,G,M]. Female: G, P5. Male: M, P5 (L = left, R = right).
|
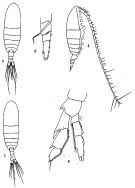 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.2, 4-8]. As Neocalanus tenuicornis. Female (from E China Sea): 4, habitus (dorsal); 5, idem (lateral right side); 6, endopod of P1 (anterior). Male: 7, habitus (dorsal); 8, left P5 (posterior).
|
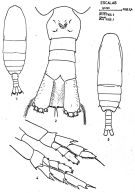 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.4]. As Calanus tenuicornis. Female (from Canarias Is.): 1, habitus (dorsal); 2, urosome (ventral). Male: 3, habitus (dorsal); 4, P5.
|
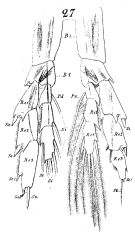 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.27]. As Calanus tenuicornis. Male: 27, P5 (anterior surface).
|
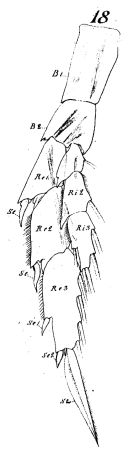 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.18]. As Calanus gracilis. Female; 18, P5 (anterior surface). B1 = basipodite 1 (= Coxa); B2 = basiopodite 2 (= Basis): Re = exopodite; Ri = endopodite.
|
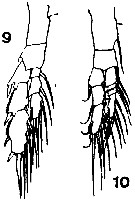 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.3, Figs.9-10]. As Calanus tenuicornis. Female: 9, P5 (posterior); 10, P1 (anterior).
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.4, Figs.1-3]. As Calanus tenuicornis. Female: 2, habitus (lateral). Male: 1, habitus (dorsal); 3, P5.
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.8, Fig.2]. As Neocalanus tenuicornis. Female: a-b, habitus (dorsal and lateral, respectively); c, P1; d, P3; e, P5. Male: f, habitus (dorsal); g, urosome (dorsal); h, head (lateral); i, P1; j, P5. Scale bars in mm.
|
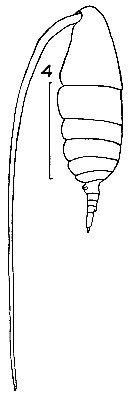 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.33, Fig.4 (4)]. Female (from Kaikoura, New Zealand): 4, habitus (lateral). Nota: Proportional lengths of abdominal segments 2 and 3: 4.8:3.8. Scale bar: 1 mm.
|
 issued from T. Mori inZool. Mag. Tokyo, 1929, 41 (486-487). [Pl. III, Figs.20-23]. As Calanus tenuicornis. Female (from Chosen Strait, Korea-Japan): 20, urosome; 21, P2; 22, P3; 24, P5.
|
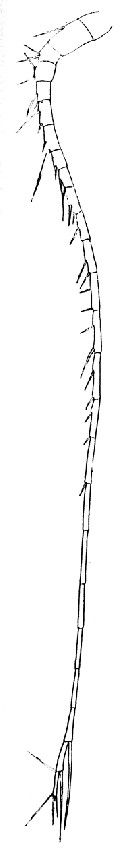 issued from T. Mori inZool. Mag. Tokyo, 1929, 41 (486-487). [Pl. IV, Fig.1]. As Calanus tenuicornis. Female: 1, A1.
|
 issued from : A. Fleminger in Mar. Biol., 1985, 88. [p.284, Fig.6, G]. Male (NE, SE Pacific): Left A1 proximal segments (ventral view); Nota: see remarks in Calanus s.l. pacificus californicus (Fleminger, 1985, p.275) concerning the dimorphism in the female A1.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.6, Fig.13]. As Calanus tenuicornis. Female: 13, habitus (dorsal). Ce = cephalothorax; Th = thoracic segment; Ab1-2 = urosomal segment 1 (genital segment: = genital double-somite).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.6, Fig.12]. As Calanus tenuicornisMale: 12, habitus (dorsal). Ce = cephalothorax; Th1 = thoracic segment 1; Ab 5 = anal segment.
|
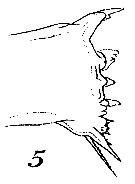 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.7, Fig.5]. As Calanus tenuicornisFemale: 5, masticatory edge of Md (anterior view).
|
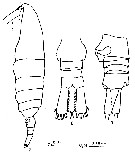 Issued from : F. Alvarez-Marques in Rev. Biol. Univ. Oviedo, 1984, 2. [p.115, Pl. I, Figs.7, 8, 9]. Female (from off Gijon, NW Spain): 7, habitus (lateral); 8, urosome (dorsal); 9, urosome (lateral) with spermatheca)
|
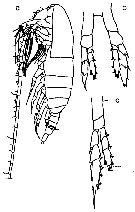 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.748, Pl. 69, fig.90 a-c]. Female: c, P5. Male: a, habitus (lateral); b, P5.
|
 ssued from : K.A. Brodsky, N.V. Vyshkvartzeva, M.S. Kos & E.L. Markhaseva in Opred. Faune SSSR, 1983, 135. [p.186, Fig.81]. As Neocalanus tenuicornis. Female and Male (after Brodsky, 1948, 1950, 1972).
|
 Issued from M. Rose in Résult. Camp. scient. Prince Albert I, 1929, 78, p.7. Remarques concernant Neocalanus tenuicornis ( = Mesocalanus tenuicornis) identifié aux cours des campagnes de 1911 et 1912.
|
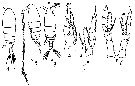 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. As Calanus (Neocalanus) tenuicornis. After Brodsky, 1950. Female : 1, habitus (dorsal); 2, same with A1 (lateral); 5, P5. Male: 3, habitus (dorsal); 4, P1; 6, P5.
| | | | | Compl. Ref.: | | | Mackas & Sefton, 1982 (p.1173, Table 1); Vives, 1982 (p.290); Dessier, 1983 (p.89, Tableau 1, Rem., %); Tremblay & Anderson, 1984 (p.4, Rem.); Roe, 1984 (p.356); Cooney & Coyle, 1985 (p.310); Wishner & Allison, 1986 (tab.2); Madhupratap & Haridas, 1986 (p.105, tab.1); Hirakawa & al., 1990 (tab.3); Hirakawa, 1991 (p.376: fig.2); Shih & Marhue, 1991 (tab.2, 3); Hattori, 1991 (tab.1, Appendix); Hirakawa & al., 1995 (tab.2); Shih & Young, 1995 (p.68); Kotani & al., 1996 (tab.2); Siokou-Frangou, 1997 (tab.1); Suarez-Morales & Gasca, 1997 (p.1525); Alvarez-Cadena & al., 1998 (t.4); Suarez-Morales & Gasca, 1998 a (p108); Kovalev & Piontkovski, 1998 (p.214, 217, Rem.); Dolganova & al., 1999 (p.13, tab.1); Goldblatt & al., 1999 (p.2619, tabl. 2); Razouls & al., 2000 (p.343, Appendix); Seridji & Hafferssas, 2000 (tab.1); Haury & al., 2000 (p.69, Table 1, Fig.7); Mackas & al., 2001 (p.685, Tab. 3, 6, fig.3: interannual changes in species composition); Rebstock, 2001 (tab.2); 2002 (p.71, Table 3, 5, Fig.2, climatic variability); Holmes, 2001 (p.38); Peterson & al., 2002 (p.353); Peterson & al. 2002 (p.381, Table 2, interannual abundance); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, fig.5, 6); Yamaguchi al., 2002 (p.1007, tab.1); Mackas & Galbraith, 2002 (p.725, tab.2a, 2b); 2002 a (p.423, Table 2); Bode & al., 2003 (p.85, Table 1, abundance); Peterson & Keister, 2003 (p.2499, interannual variability); Keister & Peterson, 2003 (p.341, Table 2, abundance, cluster species vs hydrological events); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.325, tab.1); Hsieh & al. 2004 (p.397,tab. 1); Daly Yahia & al., 2004 (p.366, fig.1); CPR, 2004 (p.56, fig.163); Lo & al., 2004 (p.89, tab.1); Fernandez & al., 2004 (p.501, tab.5); Shimode & al., 2005 (p.113 + poster); Mackas & al., 2004 (p.875, Table 2); Waggett, 2005 (p.17, Table 3.3, behaviour); Mackas & al., 2005 (p.1011, 1021, tab.2, 3); Prusova & Smith, 2005 (p.76); Zuo & al., 2006 (p.162: tab.1); Isari & al., 2006 (p.241, tab.II); Lavaniegos & Jiménez-Pérez, 2006 (p.147, tab.2, 4, Rem.); Mackas & al., 2006 (L22S07, Table 2); Ware & McQueen, 2006 (p.28, Table B1, weight ranges); Zervoudaki & al., 2006 (p.149, Table I); Hooff & Peterson, 2006 (p.2610); Koppelmann & Weikert, 2007 (p.266: tab.3); Alabaina & Irigoien, 2007 (p.435: Tab.1); Valdés & al., 2007 (p.103: tab.1); Cornils & al., 2007 (p.278, Table 2); Mackas & al., 2007 (p.223, climatic change index); Cabal & al., 2008 (289, Table 1); Humphrey, 2008 (p.84: Appendix A); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.518); Fernandes, 2008 (p.465, Tabl.2); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Ohtsuka & al., 2008 (p.115, Table 4); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); Raybaud & al., 2008 (p.1765, Table A1); Waggett & Buskey, 2008 (p.111, fig.2, Table 1); Galbraith, 2009 (pers. comm.); Miyashita & al., 2009 (p.815, Tabl.II); Labat & al., 2009 (p.1746, Table 2); Brugnano & al., 2010 (p.312, Table 3); Hafferssas & Seridji, 2010 (p.353, Table 2); Williamson & McGowan, 2010 (p.273, Table 3, Pacific central gyres: N and S); Cornils & al., 2010 (p.2076, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Mazzocchi & Di Capua, 2010 (p.424); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Nowaczyk & al;, 2011 (p.2159, Table 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Batten & Walne, 2011 (p.1643, Table I, abundance vs temperature interannual variability); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Mazzocchi & al., 2011 (p.1163, fig.6, long-term time-series 1984-2006); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Mazzocchi & al., 2012 (p.135, annual abundance 1984-2006); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Brugnano & al., 2012 (p.207, Table 2); Dorgham & al., 2012 (p.473, Table 4: abundance vs season); Lavaniegos & al., 2012 (p. 11, Appendix); Hidalgo & al., 2012 (p.134, Table 2, 3, figs.6, 8, occurrence vs hydrology); Takahashi M. & al., 2012 (p.393, Table 2, water type index); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Siokou & al., 2013 (p.1313, fig.4, 8, biomass, vertical distribution); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Fernandez de Puelles & al., 2014 (p.82, Table 3, seasonal abundance); Mazzocchi & al., 2014 (p.64, Table 3, 4, 5, spatial & seasonal composition %); Fierro Gonzalvez, 2014 (p.1, Tab. 3, 5, occurrence, abundance); Chiba S. & al., 2015 (p.968, Table 1: length vs climate); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence) ; Ohtsuka & Nishida, 2017 (p.565, Table 22.1);El Arraj & al., 2017 (p.272, Table 2); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Hure M. & al., 2018 (p.1, Table 1: abundance, % composition); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution); Bode-Dalby & al., 2026 (p.6-14, figs and tables) | | | | NZ: | 22 | | |
|
Distribution map of Mesocalanus tenuicornis by geographical zones
|
| | | | | | | | | | | | | | | | | |  issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.11, Fig.1]. issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.11, Fig.1].
Probability of occurrence of Calanus ( = Mesocalanus) tenuicornis in different environments.
T: Tropical waters (above 36.00 p.1000 salinity and above 20°C temperature); SST: Surface subtropical waters (salinity around 36 p.1000 and temperature 18°C or less); DST: Deeper shelf waters (salinity between 34 and 36, temperature under 20°C p.1000); SS: Surface shelf waters (same salinity and temperature above 20°C);
C: coastal waters with low salinity and variable temperature.
In each column no shading means no probability of finding the species in the samples from this environment; horizontal shading indicates the probability of finding the sepecies in percentages less than one in samples from this environment; cross shading indicates the probability of finding it in percentages higher than one and black shading represents the probability of finding it in the largest percentages of the total number of copepods.
Nota: For Björnberg, this species appeared only sparingly in the samples belonging to southern waters off Brazil with larger frequency in surface subtropical and in greater numbers in deeper shelf waters. It can be classified as an eurythermic cryophile prefering the lower end of the temperature range for warm waters. |
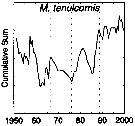 issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 u]. issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 u].
Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California (31°-35°N, 117°-122°W.
Cumulative sums of nonseasonal anomalies from the long-term means of copepod abundance from years 1950 to 2000.
A negative slope indicates a period of below-average anomalies; a positive slope indicates a period of above-average anomalies. Abrupt changes in slope indicate step changes.
The October 1966 cruise (prior to the increase in sampling depth), March 1976 cruise (prior to the 1976-77 climatic regime shift), and October 1988 cruise (prior to the hypothesized 1989 climatic regime shift) are marked with vertical lines. |
 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 1]. As Calanus tenuicornis (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 1]. As Calanus tenuicornis (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
| | | | Loc: | | | Cosmopolite: (sub-Antarct.: SW & SE Pacif., sub-Arct.: rare), mainly tropical and temperate): South Africa (E & W), Atlant., Brazil, Caribbean Colombia, Caribbean Sea, Belize, Yucatan, G. of Mexico, off Cape Hatteras, Sargasso Sea, off E Cape Cod, off E Nova Scotia, Flemish Cape, North Sea, G. of Guinea, Canary Is., Morocco-Mauritania, Ibero-moroccan Bay, Portugal, off Coruña, off Gijon, Bay of Biscay, off Ireland (N & SW), Medit. (Alboran Sea, Sidi Fredj coast, Baleares, Banyuls, Ligurian Sea, Tyrrhenian Sea, G. of Napoli, Strait of Messina, Taranto, Adriatic Sea, Ionian Sea, Aegean Sea, Thracian Sea, Lebanon Basin, W Egyptian coast, Alexandria, Black Sea), G. of Aqaba, off Sharm El-Sheikh, Red Sea, Arabian Sea, Natal, Madagascar, Rodrigues Is., Indian, Bay of Bengal, Indonesia-Malaysia, SW Celebes, Pacif. (W equatorial), China Seas (East Yellow Sea, China Sea, South China Sea), Taiwan, Kuroshio Current, Okinawa, Yellow Sea, Korea, Korea Strait, Tsushima Straits, Japon (Ishikari Bay, Kuchinoerabu Is., E, NE), Honshu (Toyama Bay) off S Shikoku Is., off N Hawaii, G. of Alaska (Columbia Bay), British Columbia, Hecate Strait, Vancouver Is., off Washington coast, Oregon (Yaquina, off Newport), California, Santa Monica Basin, W Baja California, G. of California, Pacific (central: N and S gyres), Clipperton Is., W Costa Rica, Bahia Cupica (Colombia), Peru, Chile (S, off Valparaiso, off Santiago), S Pacif. (NPFZ), New Zealand (Kaikoura, SE North Island) | | | | N: | 281 | | | | Lg.: | | | (22) F: 1,8; M: 1,8-1,5; (34) F: 1,84-1,78; (35) [N-Z] F: 2,1-1,8; (38) F: 2,18-2,12; (47) F: 2,5-1,9; M: 1,95-1,85; (72) F: 2,22-1,68; M: 1,88; (101) F: 1,87-1,68; M: 1,8; 1,56; (104) F: 2; (116) F: 2,32; M: 2,2; (125) F: 2,1-1,99; M: 1,77; (131) F: 2,05-1,54; M: 1,87-1,51; (142) F: 2-1,8; M: 1,8-1,5; (180) F: 2,06-1,65; M: 1,8-1,72; (187) F: 2,2-2,06; (196) F: 3,4-2; M: 1,9-1,7; (199) F: 2,13-1,52; M: 1,75-1,44; (237) F: 2,0; M: 1,85-1,6; (290) F: 2,05-2; M: 1,9-1,8; (327) M: 1,84; (330) F: 2,39-1,64; M: 2,02-1,71; (432) F: 2,5-2; (449) F: 2,5-1,9; M: 1,95-1,85; (786) F: 2,19-2,13; M: 2,12-1,81; (866) F: 1,5-2,2; M: 1,5-2; (920) F: 2,02 ± 0,017; (927) F: 2,05-2,08 [winter]; 1,95-1,99 [Fall]; (991) F: 1,8-2,5; M: 1,5-2,2; (1023) F: 1,76-1,89; M: 1,63-1,76; (1137) F: 2,26-1,85; M: 2,09-1,95; {F: 1,50-3,40; M: 1,50-2,20}
Chiba S. & al., 2015 (p.971, Table 1: Total length female (June-July) = 2.5 mm [optimal SST (°C) = 16.7]
| | | | Rem.: | epi- to bathypelagic.
Sampling depth (sub-Antarct.) : 0-50 m. In vertical tow 4000-3000 m (Harding, 1974: Rem.: possible contaminant species from higher up the water column).
For Marshall & Orr (1972, p.8) the size is small (1.5-1.8 mm). A1 is one and half times the length of the whole body. There are no teeth on the inner margin of the coxa in P5. In the female the genital segment is much produced ventrally.
After Ehrhardt (1967, p.888, 893) the species is found in the southwest Tyrrhenian Sea in water salinity between 37.09-38.04 p.1000 (mainly between 37.84- 38.04) and for the Oxygen between 4.45-6.40 cm3/l (mainly between 5.29-5.66).
Plentiful references, in general under the denomination of Calanus tenuicornis, more seldom Neocalanus tenuicornis.
See remarks in M. lighti.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 3 corresponding to large filter feeding herbivorous.
See in DVP Conway & al., 2003 (version 1)
R. Stephen, 2007 : Data sheets of NIO, Kochi, India (on line). | | | Last update : 11/03/2026 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 27, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
