|
|
 |
|
Calanoida ( Order ) |
|
|
|
Calanoidea ( Superfamily ) |
|
|
|
Calanidae ( Family ) |
|
|
|
Neocalanus ( Genus ) |
|
|
| |
Neocalanus gracilis (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Calanus gracilis Dana, 1849; Brady, 1883 (p.35, figs.F,M); T. Scott, 1894 b (p.25); Giesbrecht, 1892 (p.90, 128, 770, figs.F,M); Giesbrecht & Schmeil, 1898 (p.17, Rem. F,M); Thompson & Scott, 1903 (p.232, 241); Esterly, 1905 (p.128, figs.F, Rem.F,M); Pearson, 1906 (p.5, Rem.); Farran, 1908 b (p.20); Wolfenden, 1911 (p.193); Pesta, 1920 (p.495); Rose, 1925 (p.151); Farran, 1926 (p.229); 1929 (p.207, 217); Dakin & Colefax, 1933 (p.203); Rose, 1933 a (p.60, figs.F,M); Farran, 1936 a (p.77); Mori, 1937 (1964) (p.16, figs.F); Chiba & al., 1957 (p.306); 1957 a (p.11); Furnestin, 1960 (p.167); Heinrich, 1960 (p.36); Gaudy, 1962 (p.93, 99, Rem.: p.100, Pl.II: juv., Tableau 2: development); Shmeleva, 1963 (p.141); V.N. Greze, 1963 a (tabl.2); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Gaudy, 1963 (p.19); Shmeleva, 1965 b (p.1350, lengths-volume-weight relation); Pavlova, 1966 (p.43); Ehrhardt, 1967 (p.736, geographic distribution), Rem.); Berdugo & Kimor, 1968 (p.448); Dowidar & El-Maghraby, 1970 (p.267); Carli, 1971 (p.373, tab.2); Lee & al., 1971 (p.1150); Apostolopoulou, 1972 (p.327, 331); Nival & al., 1972 (p.63, respiration); Brodsky, 1972 (1975) (p.9, 71, 87, 116, 121, figs.F,M); Vyshkvartzeva, 1972 (1975) (p.188, figs.M); 1976 (p.14); 1977 a (p.97, figs.); Porumb, 1980 (p.168); Vaissière & Séguin, 1980 (p.23, tab.2); Svetlichnyi, 1980 (p.28, Table 1, passive submersion); Petipa & Borichenko, 1985 (tab.2); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea); Gorsky & al., 1988 (p.133, Table I, C-N composition); Yoo, 1991 (tab.1); Seguin & al., 1993 (p.23, occurrence); Errhif & al., 1997 (p.422); Gilabert & Moreno, 1998 (tab.1, 2); Moraitou-Apostolopoulou & al., 2000 (tab.I); Sautour & al., 2000 (p.531, Table II, abundance); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters); Pagano, 2009 (p.115); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Sobrinho-Gonçalves & al., 2013 (p.713, Table 2, fig.8, seasonal abundance vs environmental conditions); Zaafa & al., 2014 (p.67, Table I, occurrence):
Calanus (Neocalanus) gracilis : Fleminger, 1967 a (tabl.1);
Cetochilus longiremis Claus, 1863 (p.171, fig.F); Gourret, 1889 ;
? Calanus americanus Herrick, 1887;
Megacalanus gracilis : A. Scott, 1909 (p.12) | | | | Ref.: | | | Sars, 1925 (p.7); Sewell, 1929 (p.26); Rose, 1929 (p.7); Wilson, 1932 a (p.28, figs.F,M); Farran, 1936 a (p.77); Dakin & Colefax, 1940 (p.85, figs.F,M); Wilson, 1942 a (p.195, fig.M); Vervoort, 1946 (p.40, Rem.); Oliveira, 1946 (p.454, figs.F); Sewell, 1947 (p.17, Rem.); Farran & Vervoort, 1951 (n°32, p.3, figs.F,M); Marques, 1953 (p.91); Tanaka, 1956 (p.261, Rem.M); Vervoort, 1957 (p.31, Rem.); Fish, 1962 (p.5); Grice, 1962 (p.177, figs.F); Brodsky, 1962 c (p.98, figs.F); Björnberg, 1963 (p.13, figs.F); Vervoort, 1963 b (p.82, Rem.); Chen & Zhang, 1965 (p.27, figs.F); Owre & Foyo, 1967 (p.31, figs.F,M); Mazza, 1967 (p.60, 62: clé juv., F,M); Vilela, 1968 (p.9, fig.M); Vidal, 1968 (p.16, figs.F); Park, 1968 (p.532, figs.M, Rem.M); Ramirez, 1969 (p.41, figs.F, Rem.); Corral Estrada, 1970 (p.66, figs.F, M, Rem.); Bradford, 1972 (p.32, figs.F); Grice, 1972 (p.177, figs.F); Razouls, 1972 (p.94, Annexe: p.8, figs.F,M); Marques, 1973 (p.232); 1974 (p.11); Chen & al., 1974 (p.103, figs.M); Bradford & Jillett, 1974 (p.6); Marques, 1976 (p.985); Björnberg & al., 1981 (p.618, figs.F,M); Schnack, 1982 (p.145, fig.Md); Roe, 1984 (p.356); Fleminger, 1985 (p.276, 285, Table 4, Rem.: A1); Regner, 1985 (p.20); Sazhina, 1985 (p.28, figs.N); Mauchline, 1987 (p.719); Bradford, 1988 (p.74, 76, 79, Rem.); Nishida, 1989 (p.173, fig. 3, table 1, 2, 3: dorsal hump); Schnack, 1989 (p.137, tab.1, fig.6: Md); He & al., 1992 (p.250); Bradford-Grieve, 1994 (p.41, figs.F,M, fig.98); Chihara & Murano, 1997 (p.740, Pl.68: F,M); Barthélémy, 1999 a (p.10, Fig.19, H); Bradford-Grieve & al., 1999 (p.877, 908, figs.F,M); Miller, 2002 (p.135, 139B, Rem.); Bucklin & al., 2003 (p.335, tab.2, fig. 2, Biomol.); Conway & al., 2003 (p.149, figs.F,M, Rem.); Mulyadi, 2004 (p.23, figs.F,M, Rem.); Conway, 2006 (p.8: copepodite stages 1-6, Rem); Machida & al., 2006 (p.1071, tab.1,3, fig.3,4, molecular phylogeny); Avancini & al., 2006 (p.59, Pl. 28, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.902, figs.F,M, Rem.) | 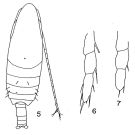 Issued from : T.S. Park in Fishery Bull. Fish Wild. Serv. U.S., 1968, 66 (3). [p.531, Pl.1, Figs.5-7]. Male: 5, habitus (dorsal); 6, endopod of left P5; 7, endopod of left P5 ( In some specimens the setae of the endopod are almost completely reduced).
|
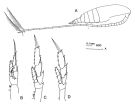 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). New Zealand Oceanographic Institute Memoir, 102, 1994. [p.42, Fig.16]. Female: A, habitus (lateral right side); B, P1; C, P2; D, P5.
|
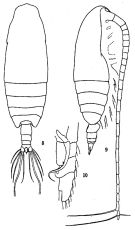 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.1, 8-10]. Female (from E China Sea): 8, habitus (dorsal); 9, idem (lateral right side); 10, endopod of P1 (anterior).
|
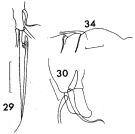 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.42, Lam. VI, figs.29, 30, 34 ]. Female (from off Mar del Plata): P1 (details); 30, basipodal segment 2 and endopodal segment 1 of P1; 34, outer margin of basipodal segment 1 of Mx2. Scale bars in mm: 0.1 (29); 0.2 (30); 0.1 (34).
|
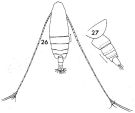 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.40, Lam. V, figs. 26, 27]. Female (from off Mar del Plata): 26, habitus (dorsal); 27, urosome (lateral left side). Scale bars in mm: 1 (26); 0.2 (27).
|
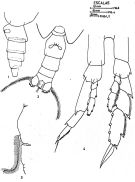 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.5]. Female (from Canarias Is.): 1, urosome (lateral left side); 2, idem (ventral); 3, outer margin of basipodal segment 1 of Mx2. Nota: Right A1 longer than the left (for an individual: right length = 5.20 mm and left length = 4.40 mm). Male: P5. The right and left A1 of the same length.
|
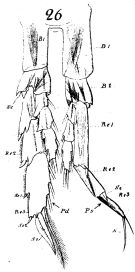 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.26]. As Calanus gracilis. Male; 26, P5 (anterior surface). Pd = right leg; Ps = left leg; B1 = basis 1 (= Coxa); B2 = basis 2 (= Basis); Re = exopodite; Ri = endopodite; Se = outer seta; St = terminal seta.
|
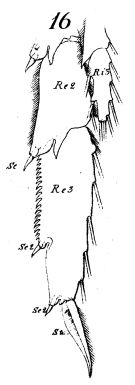 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.16]. As Calanus gracilis. Male; 16, P3 (partial, anterior surface)
|
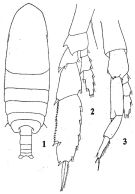 issued from : Q.-c. Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.1, Figs.1-3]. Male (from China Seas): 1, habitus (dorsal); 2, P4; 3, P5.
|
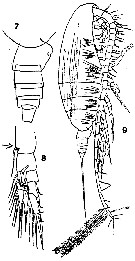 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.4, Figs.7-9]. As Calanus gracilis. Female: 7, urosome (lateral); 8, P1 (anterior); 9, habitus (lateral). Nota: There is a process (tooth-like) at the base of the marginal seta of the basipodal segment 2 of P1.
|
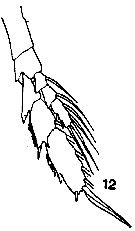 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.3, Fig.12]. As Calanus gracilis. Female: P5 (anterior).
|
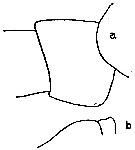 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.128, Fig.4]. As Calanus gracilis. Female (from San Diego Region): a, genital segment (lateral); b, outer margin of 1st basipodal segment of Mx2.
|
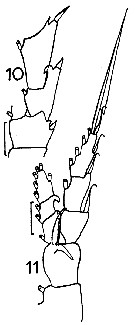 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.33, Fig.4 (10-11)]. Female (from Kaikoura, New Zealand): 10, P2 (basipod2 and exopod segments 1 and 2); 11, P1. Scale bars: 0.1 mm (10, 11). Nota: Recognised by P1, the incurved hook on the 1st exopod segment of P2 and the long A1 which are almost twice the body length.
|
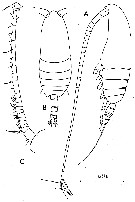 issued from : C. Razouls in Th. Doc. Etat Fac. Sc. Paris VI, 1972, Annexe. [Fig.23]. Female (from Banyuls, Gulf of Lion)): A, habitus (lateral). Male: B, habitus (dorsal); C, forehead and right A1 (dorsal view).
|
 issued from : R.J. Machida, M.U. Miya, M. Nishida & S. Nishida in Mar. Biol., 2006, 148. [p.1077, Table 5]. Comparisons of selected biological characteristics of 6 species olus 1 subspecies of Neocalanus.
|
 issued from : N.V. Vyshkvartzeva in Issled. Fauny Moreï, 1972, 12 (20). [p.168, Fig.7, 2a, 2b, 2g). As calanus gracilisFemale Md (masticatory edge): 2a, lateral aspect; 2b, left Md (above view); 2g, right Md (above view).
|
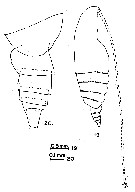 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.176, Pl.1, Figs.19-20]. Female (from equatorial Pacific): 19, habitus (lateral); 20, posterior part of thorax and urosome (lateral, left side).
|
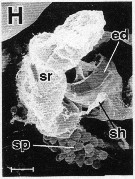 issued from : R.-M. Barthélémy in These Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.19, H]. Female (from Gulf of Marseille, NW Mediterranean Sea): H, detail of the genital area (left side. ed = egg-laying duct; sh = shell duct; sr = seminal receptacle. Note the spherical spermatozoides (sp) outside the seminal receptacle reared.
|
 Issued from : G.S. Brady in Rep. Scient. Results Voy. Challenger, Zool., 1883, 8 (23). [Pl. V]. As Calanus gracilis. Female: 1, habitus (dorsal); 2, Mx1; 3, terminal seta of P1; 4, P4. Male: 5, P4; 6, right P5.
|
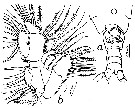 issued from : T.K.S. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.13, Fig.3]. Female (from Brazilian waters): a, last thoracic segment and urosome (ventral); b, Mx1.
|
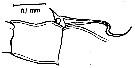 issued from : T.K.S. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.14, Fig.4]. Female: Double hook on the 2nd basal segment of P1. Nota: The distinct double hook on P1 is already present in the young copepodites.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.6, Fig.1]. As Calanus gracilis. Male: 1, habitus (dorsal).
|
 issued from : P.E. Lapernat & C. Razouls in Vie Milieu, 2002, 52 (1). [p.28, Pl. VI, fig.1]. Masticatory edge of Md gnathobase female (from off Malta, Mediterranean Sea). Nota: Itoh's index value: 383.4 (number of teeth : 10). For Schnack (1989) = 444.
|
 issued from : P.E. Lapernat & C. Razouls in Vie Milieu, 2002, 52 (1). [p.20, Pl. I, fig.2]. Masticatory edge of Md gnathobase female (from off Malta, Mediterranean Sea).
|
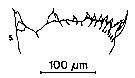 issued from : S.B. Schnack in Crustacean Issue, 1989, 6. [p.143, Fig.6: 5]. 5, Neocalanus gracilis (from off NW Africa, upwelling region): Cutting edge of Md.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Fig.1]. As Calanus gracilis. Female: 1, A2 (anterior view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Fig.4]. As Calanus gracilis. Female: 4, gnathobase of Md (anterior view). Si = inner seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Fig.7]. As Calanus gracilis. Male: 7, gnathobase of Md (posterior view). Nota: Same scale than the Md drawing female
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Fig.3]. As Calanus gracilis. Male: 3, A2 (anterior view). Same scale than the A2 drawing female.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Fig.9]. As Calanus gracilis. Female: Md (mandibular palp).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Figs. 17, 18]. As Calanus gracilis. Female: 17, Mx2 (posterior view). Male: 18, Mx2 (posterior view). Drawings 17 and 18 at the same scale.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Figs. 20, 21]. As Calanus gracilis. Female: 20, Mxp (anterior view). Male: 21, Mxp (posterior view); Drawings 20 and 21 at same scale.
|
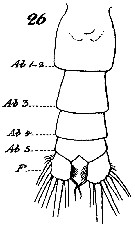 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 7, Fig.26]. As Calanus gracilis. Female: 26, abdomen (ventral). Ab 1-2 = genital dpuble-somite; F = Furca (= caudal rami).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Figs.6, 7, 8]. As Calanus gracilis. Female: 6, basis 1, 2 and exopod of P2 (posterior view); 7, exopoite 1 of P2; 8, P1 (anterior view). Basis 1 = coxa; basis 2 = basis.
|
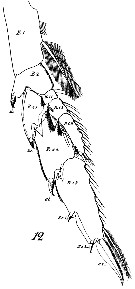 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.12]. As Calanus gracilis. Female: 12, P3 (anterior view).
|
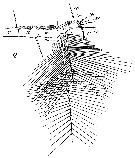 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.2]. As Calanus gracilis. Female: 2, distal portion of A1. Ae = aesthetasc. St = terminal seta. Sdi = distal seta (ventral view)
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.4]. As Calanus gracilis. Female: 4, A1, segments 13 to 25.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 8, Fig.4]. As Calanus gracilis. Female: 4, A1, segments 1 to 12.
|
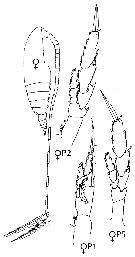
 issued from : R. Gaudy in Rev. Trav. St. Mar. End., Bull. 27 (42). [p.125, Tableau 2]. Identification of copepodids stages Calanus gracilis ( = Neocalanus gracilis). A: Stage; B: number of swimming legs; C: number of thoracic segments; D: number of abdominal segments; E, Size.
|
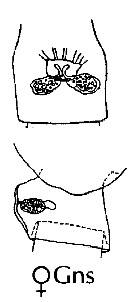
 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.981, Fig. 7.20]. Female: habitus (lateral); P1; P2; P5. Nota: Basis of P1 with visible spine at base of inner seta. Basis of P2-P5 naked
|
 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.981, Fig. 7.20]. Female: genital segment (ventral and lateral, respectively). Nota: genital segment slightly swollen ventrally. Spermathecae of typical structure, more simple than in N. robustior.
|
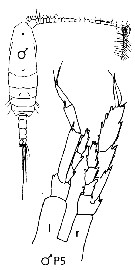 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.981, Fig. 7.20]. Male: habitus (dorsal); P5 (I = left leg; r = right leg). Nota: P5 significantly asymmetrical, with left leg modified. Setation of left P5 endopodite: endopodal segments 1 and 2 with 1 inner seta each; endopodal segment 3 with 6 setae.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.23, Fig.10]. Female (from 07°29'S, 121°05'E): a, habitus (dorsal); b, last thoracic segment and urosome (lateral right side); c, P1; d, P5 (exopod missing). Male: e, habitus (dorsal); f, last thoracic segment with P5 and urosome (lateral); g, P5 (endopod missing).
|
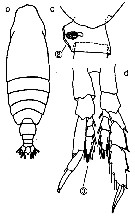 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.747, Pl. 68, fig.86 b-d]. Female: b, habitus (dorsal); c, posterior corner of last thoracic segment and genital segment (lateral). Male: d, P5. Nota: Numbers show species characteristics. Fig.b, after Owre & Foyo, 1967; Figs.c, d, after Brodsky, 1972.
|
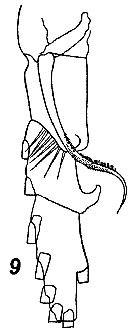 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.41, Fig.11, 9]. Neocalanus gracilis after Brodsky, 1972. 9, distal part of basis and endopod of P1.
|
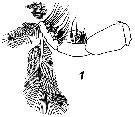 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.89, Fig.23: 1]. Neocalanus gracilis after Vyshkvartzeva, 1977 b. Mxp male.
|
 Issued from M. Rose in Résult. Camp. scient. Prince Albert I, 1929, 78, p.7. Remarques concernant Neocalanus gracilis identifié aux cours des campagnes de 1904, 1910, 1905,1911, 1912, 1913 et 1914.
|
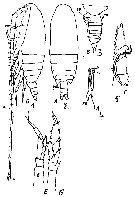 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. As Calanus (Neocalanus) gracilis. From Gisebrecht, 1892, after Brodsky, 1962. Female: 1, habitus (lateral); 2, same (dorsal); 3, abdomen (latrral); 4, basis of P5; 5, P5. Male: 6, P5.
| | | | | Compl. Ref.: | | | Jespersen, 1939 (p.38, Rem.); Massuti Alzamora, 1942 (p.110); Lysholm & al., 1945 (p.7); Oliveira, 1945 (p.191); Sewell, 1948 (p.329, 400, 508, 513, 516, 531, 544, 555); C.B. Wilson, 1950 (p.269); Krishnaswamy, 1953 (p.108); King & Hida, 1955 (p.11); Duran, 1955 (p.52); Yamazi, 1958 (p.147, Rem.); Fagetti, 1962 (p.6); Cervigon, 1962 (p.181, tables: abundance distribution); Duran, 1963 (p.11); Grice, 1963 a (p.495); Unterüberbacher, 1964 (p.14, Rem.); De Decker & Mombeck, 1964 (p.13); Anraku & Azeta, 1965 (p.13, Table 2, fish predator) Grice & Hulsemann, 1965 (p.223); Neto & Paiva, 1966 (p.18, Table III); Mazza, 1966 (p.69); Mullin, 1966 (p.546, Table I, III, diet); Grice & Hulsemann, 1967 (p.13); De Decker, 1968 (p.45); Vinogradov, 1968 (1970) (p.78, 79, 80, 81, 83, 91, 268, 269, 277); Evans, 1968 (p.13); Park, 1970 (p.474); Morris, 1970 (p.2300); Timonin, 1971 (p.281, trophic group); Deevey, 1971 (p.224); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 a (p.322); Bainbridge, 1972 (p.61, Appendix Table II, vertical distribution %); Rudyakov, 1972 (p.886, Table 1: sinking rate); Boucher & Thiriot, 1972 (p.47, Tableau 4); Razouls S., 1972 b (p.2, respiration); Heinrich, 1973 (p.95); Björnberg, 1973 (p. 283, 388); S. Razouls, 1972 (p.1, respiration); 1974 (147, oxygen rate); Ikeda, 1974 (p.1, respiration, ammonia excretion rate); Corral Estrada & Pereiro Muños, 1974 (tab.I); Vives & al., 1975 (p.35, tab.II, IV); Arashkevich, 1975 (p.351, digestion time); Mullin & Evans, 1976 (p.119, distribution chart, length, sex ratio, vertical distribution); Tranter, 1977 (p.596); Carter, 1977 (1978) (p.35); Deevey & Brooks, 1977 (p.256, Table 2, Station "S"); Timonin & Voronina, 1977 (p.283)); Dessier, 1979 (p.201); Brenning, 1981 (p.1, Rem.: p.9); Vives, 1982 (p.290); Kovalev & Shmeleva, 1982 (p.82); Dessier, 1983 (p.89, Tableau 1, Rem., %); Tremblay & Anderson, 1984 (p.4, Rem.); De Decker, 1984 (p.315); Cummings, 1984 (p.163, Table 2); Scotto di Carlo & al., 1984 (p.1041); Alvarez-Marques, 1984 (p.113, annual abundance); Guangshan & Honglin, 1984 (p.118, tab.); Regner, 1985 (p.11, Rem.: p.23); Brenning, 1985 a (p.28, Table 2); Brinton & al., 1986 (p.228, Table 1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Wishner & Allison, 1986 (tab.2); Madhupratap & Haridas, 1986 (p.105, tab.1); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Boucher & al., 1987 (p.133, spatial distribution/physical structure); Lozano Soldevilla & al., 1988 (p.57); Dessier, 1988 (tabl.1); Madhupratap & Haridas, 1990 (p.305, fig.6: vertical distribution night/day; fig.7: cluster); Heinrich, 1990 (p.16); Timonin, 1990 (p.479); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Scotto di Carlo & al., 1991 (p.270); Ashjian & Wishner, 1993 (p.483, abundance, species group distributions); Hays & al., 1994 (tab.1); Kouwenberg, 1994 (tab.1); Heinrich, 1995 (tab.1); Shih & Young, 1995 (p.68); Siokou-Frangou, 1997 (tab.1); Go & al., 1997 (tab.1); Suarez-Morales & Gasca, 1997 (p.1525); Hwang & al., 1998 (tab.II); Alvarez-Cadena & al., 1998 (tab.1, 2, 3, 4); Noda & al., 1998 (p.55, Table 3, occurrence); Stephen, 1998 (p.341, Rem., chart); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p108); Reid & Hunt, 1998 (p.310, figs.2, 3, Rem.); Lapernat, 1999 (p.10, 55); Lapernat, 2000 (tabl.3, 4); Razouls & al., 2000 (p.343, Appendix); Escribano & Hidalgo, 2000 (p.283, tab.2); Lopez-Salgado & al., 2000 (tab.1); Haury & al., 2000 (p.69, Table 1); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); d'Elbée, 2001 b (tabl. 1); Lapernat & Razouls, 2001 (p.123, tab.1); Rebstock, 2001 (tab.2); Holmes, 2001 (p.38); Rebstock, 2002 (p.71, Table 3, 5, 6, Fig.2, climatic variability); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, figs.5, 6); Labat & al., 2002 (p.741, tab.3); Hwang & al., 2003 (p.193, tab.2); Vukanic, 2003 (p.139, tab.1); Hsiao & al., 2004 (p.325, tab.1); Lan & al., 2004 (p.332, tab.1); Lo & al., 2004 (p.218, fig.6); Fernandez de Puelles & al., 2004 (p.654, fig.7); Pusch & al., 2004 (251, tab.3); Gallienne & al., 2004 (p.5, tab.3); ; CPR, 2004 (p.57, fig.167); Lo & al., 2004 (p.89, tab.1); Berasategui & al., 2005 (p.313, fig.2); Prusova & Smith, 2005 (p.76); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: stations group); Isari & al., 2006 (p.241, tab.II); Lavaniegos & Jiménez-Pérez, 2006 (p.149, tab.2, 3, Rem.); Hwang & al., 2006 (p.943, tab.I); Mackas & al., 2006 (L22S07, Table 2); Hwang & al., 2007 (p.23); Fernandez de Puelles & al., 2007 (p.348); Albaina & Irigoien, 2007 (p.435: Tab.1); Valdés & al., 2007 (p.103: tab.1); Khelifi-Touhami & al., 2007 (p.327, Table 1); Cabal & al., 2008 (289, Table 1); McKinnon & al., 2008 (p.843: Table.I); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.518); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Raybaud & al., 2008 (p.1765, Table A1); Waggett & Buskey, 2008 (p.111, Table 1); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); C.-Y. Lee & al., 2009 (p.151, Tab.2); W Zhang & al., 2009 (p.266: table 2); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Licandro & Icardi, 2009 (p.17, Table 4); C.E. Morales & al., 2010 (p.158, Table 1); Brugnano & al., 2010 (p.312, Table 2, 3); Williamson & McGowan, 2010 (p.273, Table 3, Pacific central gyres: N and S); Cornils & al., 2010 (p.2076, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Mazzocchi & Di Capua, 2010 (p.424); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Nowaczyk & al;, 2011 (p.2159, Table 2); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Hsiao S.H. & al., 2011 (p.475, Appendix I); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Mazzocchi & al., 2011 (p.1163, fig.6, long-term time-series 1984-2006); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Mazzocchi & al., 2012 (p.135, annual abundance 1984-2006); Delpy & al., 2012 (p.1921, Table 2); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Salah S. & al., 2012 (p.155, Tableau 1); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Brugnano & al., 2012 (p.207, Table 2, 3); Bode & al., 2012 (p.108, spatial distribution vs time-series, % biomass); Lavaniegos & al., 2012 (p. 11, Appendix); Hidalgo & al., 2012 (p.134, Table 2, 3, figs.6, 8, occurrence vs hydrology); Teuber & al., 2013 (p.28, Table 1, respiration rates); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Teuber & al., 2013 (p.1, Table 2, fig.5, respiration rates, enzyme activity); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Hirai & al., 2013 (p.1, Table I, molecular marker); Tseng & al., 2013 a (p.1, Table 3, abundance); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & al., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Bode & al., 2015 (p.268, Table 1, 2, figs.3, 4, chemical components, trophic level, geographic zone); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Trottet & al., 2017 (p. 7, Table 3: resting stage); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Bode & al., 2018 (p.840, Table 1, respiration & ingestion rates, depth); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution). | | | | NZ: | 22 | | |
|
Distribution map of Neocalanus gracilis by geographical zones
|
| | | | | | | | | | | | | | | | | |  issued from : R.J. Machida, M.U. Miya, M. Nishida & S. Nishida in Mar. Biol., 2006, 148. [p.1077, Fig.4]. issued from : R.J. Machida, M.U. Miya, M. Nishida & S. Nishida in Mar. Biol., 2006, 148. [p.1077, Fig.4].
Relationship between the phylogeny and distribution pattern of the Neocalanus species.
Shaded portions of the maps represent distributions based on the litterature. Clades A-E correspond to those in Fig.3, p.1075. |
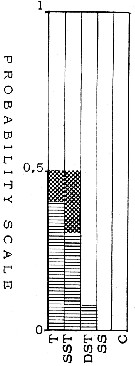 issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.14, Fig.5]. issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.14, Fig.5].
Probability of occurrence of Neocalanus gracilis in different environments.
T: Tropical waters (above 36.00 p.1000 salinity and above 20°C temperature); SST: Surface subtropical waters (salinity around 36 p.1000 and temperature 18°C or less); DST: Deeper shelf waters (salinity between 34 and 36, temperature under 20°C p.1000); SS: Surface shelf waters (same salinity and temperature above 20°C);
C: coastal waters with low salinity and variable temperature.
In each column no shading means no probability of finding the species in the samples from this environment; horizontal shading indicates the probability of finding the sepecies in percentages less than one in samples from this environment; cross shading indicates the probability of finding it in percentages higher than one and black shading represents the probability of finding it in the largest percentages of the total number of copepods.
Nota: Björnberg finds quite frequently in catches from Brazilian waters in very saline water layers (35.90 p.1000 or more) in the surface tropical and subtropical waters. It usually was caught in the upper layers during the night or in the early morning. Catches made during the day were poor in number of specimens, strongly indicating migration of the species to deep waters during the day. |
 issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998. issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998.
P. 341, Fig.5: Distribution of Neocalanus gracilis and Neocalanus tonsus in the Indian Ocean. |
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.32, Table 5]. issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.32, Table 5].
Vertical distribution of Neocalanus gracilis at the ''40-Mile station'' in the Florida Current (± 25°35'N, 79°27'W).
SL 53: 18 V 1958; SL 55: 21 VII 1958. A: during midday; B: during midnight |
 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 2]. As Calanus gracilis (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 2]. As Calanus gracilis (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
 issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.121, Fig.2]. issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.121, Fig.2].
Distribution of Neocalanus gracilis in the Pacific Ocean
Tows were taken from the surface to the depth indicared by open or filled circle. Contour intervals were chosen arbitrarily, based on breaks in the data abundances.
The copepodite stages III, IV, V, and adults were counts in aliquots (subsampled with a plankton splitter.The samples were taken during expeditions in May, Yune and August 1952; December-November 1953; September-November 1953; July-September 1955; August-September 1956; October-February 1957-1958; November 1960; June-August 1960; March-April 1960-1961; September 1964; January 1966 and March-May 1967. Most of the samples were taken by oblique tows to various depths (net: 500-550 µ mesh aperture, 1 m mouth diameter, with a flowmeter).
Nota: The distribution of N. gracilis is similar to N. robustior, both species being warm-water cosmopolites in the terminology of McGowan (1971), but the range of N. gracilis extends farther to the north and south, and this species is relatively more common in the western South Pacific than is N. robustior. |
 issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.124, Table 1]. issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.124, Table 1].
Cephalothorax length and mandibular width : Cephalothorax length, by area, of Neocalanus robustior and Neocalanus gracilis in the Pacific Ocean.
2-4 stations were pooled in each zoogeographic area. * No occurrences in samples examined.
Nota: See discussion concerning the supposed sympatric species N. gracilis and N. robustiorfrom Mullin & Evans (1976, p.127-128) in Neocalanus robustior |
 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.13]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.13].
Verical distribution of Neocalanus gracilis at eutrophic (Mauritania coast: 20°32' N, 18°36' W) (1 from night and 2 from day) and oligotrophic sites (off NW Cape Verde Islands: 21° N, 31° W) (3 from night and 4 from day) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
 Issued from S. Razouls in XXIII rd Congress of Athens, 3-11 November 1972. [p.2]. Oxygen consumed by individual (adult) in the Banyuls Bay. Issued from S. Razouls in XXIII rd Congress of Athens, 3-11 November 1972. [p.2]. Oxygen consumed by individual (adult) in the Banyuls Bay.
(1) Hydrological season in the stability period: Eté = Summer: 18-20 °C; Hiver = Winter: 13-10°C.
Espèces = species; Saison = Season; Lg céph.= cephalothoracic length; an = individual. |
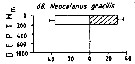 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 68- Neocalanus gracilis.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : Ya-Qu. Chen in CatCOFI Rep., 1986, Vol. XXVII. [p.220]. Issued from : Ya-Qu. Chen in CatCOFI Rep., 1986, Vol. XXVII. [p.220].
For the author, during the Krill Expedition from 23°N to 3°S in the Pacific East tropical (May-June 1974), this species shows abundances less than 0.5% of the total copepods. This species was found mainly in mid-depth to surface samples taken south of 10°N (stations 18-22). The greatest depth of occurrence was 500 m, where the number (142 individuals/1000 m3) was less than that at 50 m In night samples the species was found, in greater numbers than in day samples, between 0 and 350 m. The maximum number was 1020 individuals/m3 in 0-50 m at station 21. The species was not present in the oxygen-deficient area. (see in Subeucalanus subtenuis Chart Fig.1, hydographic conditions Figs.12, 13). |
 Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1). Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1).
Cf. explanations of these measures in Calanoides natalis from the same authors.
Compare with Neocalanus robustior. |
| | | | Loc: | | | Cosmopolite (tropical and temperate); sub-Antarct. (Indian, SW Pacif. SW), South Africa (E & W), Namibia, Angola (Baia Farta), Congo, G. of Guinea, Lagos, Dakar, Cape Verde Is., off Mauritania-NW Cape Verde Is., Great Meteor Seamount, Morocco-Mauritania, Cap Ghir, Canary Is., Portugal, Vigo, off W Cabo Finisterre, off Gijon, Bay of Biscay, off Amazon, off Rio de Janeiro, Brazil-Argentina, Rio de La Plata (offshore), Campos Basin, Barbada Is., Caribbean Colombia, Cariaco Basin, Bahia de Mochima (Venezuela), Caribbean Sea, Yucatan, Jamaica, G. of Mexico, Florida, off Bermuda, Station "S" (32°10'N, 64°30'W), Sargasso Sea, off Cape Hatteras, off Woods Hole, off E Nova Scotia, off S Newfoundland, S Greenland (rare), S Iceland, Ireland, off SW Ireland, North Sea, Cap Ghir, Ibero-moroccan Bay, off W Tangier, Medit. (Alboran Sea, Habibas Is., Sidi Fredj coast, Gulf of Annaba, Castellon, Banyuls, Berre Lagoon, Marseille, Villefrance-s-Mer, Ligurian Sea, Tyrrhenian Sea, Napoli, Messine, Gulf of Taranto, NW Tunisia, Malta, Adriatic Sea, Ionian Sea, Aegean Sea, Thracian Sea, Lebanon Basin, Alexandria, W Egyptian coast, Black Sea), Sharm El-Sheihh, Red Sea, Arabian Sea, Natal, equatorial Indian Ocean, Madras, Nicobar Is. (Nankauri Harbour), off Madagascar S, Nosy Bé, Rodrigues Is. - Seychelles, Mascarene Basin, S Indian (subtropical Convergence), W Australia, Singapore, Indonesia-Malaysia, Flores Sea, SW Celebes, Philippines, China Seas (East China Sea, South China Sea), Taiwan Strait, Taiwan (E, S, SW, NW, N, Mienhua Canyon), Yellow Sea, Okinawa, S Korea, Korea Strait, Japan, Kuchinoerabu Is., Nagazaki, Bering Sea, Alaska, British Columbia, California, W Baja California, Gulf of Caliornia, W Costa Rica, Bahia Cupica (Colombia), Hawaii, off Hawaii (N, S, E), Pacif. (W equatorial), Pacific (central gyres: N and S), Australia (Great Barrier, New South Wales), New Caledonia, New Zealand, S Pacif. (NPFZ), Clipperton Is., Pacif. (SE tropical), Chile (N & S, off Santiago, Concepcion) | | | | N: | 319 | | | | Lg.: | | | (11) F: 3,309-3,036; (14) F: 3,26-2,7; (28) F: 3-2,43; M: 2,55-2,3; (34) F: 2,95-2,48; M: 2,65; (35) [Atlant. N] F: 3,48-3,36; [N-Z] F: 3,75-3,35; (38) F: 3,9-3,5; (45) F: 3,25-3; M: 2,85-2,5; (47) F: 3,25-3; M: 2,8-2,5; (72) F: 3,74-3; M: 3,06-2,73; (75) F: 2,08-1,89; M: 1,86-1,67; (101) F: 3,33-2,11; (104) F: 3; M: 2,9; (125) F: 3,25-2,95; (135) F: 3,6; (142) F: 2,4; (150) F: 3,25-2,92; (180) F: 3,54-3; M: 2,98; (187) F: 3,53; (196) F: 4-2,8; M: 3,4-2,88; (199) F: 3,42-2,89; M: 2,96-2,66; (207) F: 3,63-3,4; M: 3,11-3; (237) F: 3,5-2,5; (254) F: ±3,75; (290) F: 3,2-2,95; (327) F: 3,41-3,15; M: 2,92-2,82; (335) F: 3,38; (336) F: 2,09-1,8; M: 1,7-1,6; (338) M: 2,85-2,7; (340) F: 3,1; 3; (432) F: 4,4-3,7; (449) F: 3,25-2,4; M: 2,8-2,5; (785) F: 4,08-3,57; (795) F: 3; (865) F: 3,1; (866) F: 2,7-3,4; M: 2,1-3; (920) F: 3,19; (991) F: 2,43-4; M: 2,3-3,1; (1122) F: 3,15; M: 2,75; (1137) F: 2,38-2,14; (1230) F: 2,4-4,0; M: 2; {F: 1,80-4,40; M: 1,60-3,40} | | | | Rem.: | Oceanic. Epi- to bathypelagic.
Sampling depth (sub-Antarct.) : 100-750 m. Sargasso Sea: 0-1000 m.
For Björnberg (1963, p.14) usually large specimens were at first mistaken for Megacalanus copepodites (Björnberg, 1959, p.138). The size and the pronounced genital swelling of the genital segment of the females might also lead to a confusion with N. robustior. The A1 reaches far beyond the caudal rami in both sexes, by 6 segments in the male. The hook on the 2nd basal segment of P1 differs from that of N. robustior (A1 reaches only up to the caudal rami).
Can be confused with Neocalanus robustior, but of inferior size and less ventrally pronounced genital segment.
Numerous references under the denomination Calanus gracilis.
For Sewell (1947, p.17) the external marginal spine divides the outer margin of the 3rd segment of exopodite of P2 to P4 in the proportions: P1 (20:15); P2 (34:35); P3 (53:35); P4 (65:33, P5 (45:25).
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a coarse-filter feeder.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 3 corresponding to large filter feeding herbivorous.
See in DVP Conway & al., 2003 (version 1)
R. Stephen, 2007 : Data sheets of NIO, Kochi, India (on line). | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 23, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
