|
|
 |
|
Calanoida ( Order ) |
|
|
|
Calanoidea ( Superfamily ) |
|
|
|
Calanidae ( Family ) |
|
|
|
Neocalanus ( Genus ) |
|
|
| |
Neocalanus robustior (Giesbrecht, 1888) (F,M) | |
| | | | | | | Syn.: | Calanus robustior Giesbrecht,1888, 1892 (p.91, 129, 770, figs.F,M); Giesbrecht & Schmeil, 1898 (p.18, Rem. F,M)); Thompson & Scott, 1903 (p.233, 242); Wolfenden, 1905 (p.996, figs.F); Esterly, 1905 (p.129, figs.F, Rem.); 1906 a (p.77, fig.M); Wolfenden, 1911 (p.193: Rem.); Farran, 1929 (p.207, 217); Rose, 1933 a (p.61, figs.F,M); Mori, 1937 (1964) (p.17, figs.F); Oliveira, 1945 (p.191); Chiba & al., 1957 (p.306); 1957 a (p.11); Marques, 1958 (p.205, fig.F); Furnestin, 1960 (p.167); Giron-Reguer, 1963 (p.25); Gaudy, 1963 (p.19, Rem.); Dowidar & El-Maghraby, 1970 (p.267); Lee & al., 1971 (p.1150); Vives & al., 1975 (p.35, III); Fernandez, 1975 (p.1, fig.17, 18, 19, 20, 21, metabolism/lux), Brodsky, 1972 (1975) (p.9, 71, 88, 121, figs.F,M); Vyshkvartzeva, 1972 (1975) (p.188, figs.); 1976 (p.14); 1977 a (p.97, figs.); Fernandez, 1978 (p.97, metabolism/food, Rem.: Table 19); Svetlichnyi, 1980 (p.28, Table 1, passive submersion); Gaudy & Boucker, 1983 (p.37, Table 1, 4, 5, Rem.: metabolism); Seguin & al., 1993 (p.23, 26: Rem.); Pagano, 2009 (p.115);
Calanus comptus : T. Scott, 1894 b (p.26, figs.M);
Calanus (Neocalanus) robustior : Fleminger, 1967 a (tabl.1);
Megacalanus robustior : A. Scott, 1909 (p.13, Rem.) | | | | Ref.: | | | Sars, 1925 (p.8); Rose, 1929 (p.7); Lysholm, 1945 (p.7); Vervoort, 1946 (p.40, 44, Rem.); Sewell, 1947 (p.17); Tanaka, 1956 (p.261); Fagetti, 1962 (p.7); Grice, 1962 (p.178, fig.F); Brodsky, 1962 c (p.98, figs.F,M); Vervoort, 1963 b (p.83, Rem.); Paiva, 1963 (p.12, figs.F); Chen & Zhang, 1965 (p.27, figs.F); Owre & Foyo, 1967 (p.32, figs.F,M); Vinogradov, 1968 (1970) (p.268, 277); Vidal, 1968 (p.16, figs.F); Park, 1968 (p.533, figs.M, Rem.F,M); Tanaka, 1969 (p.255, figs.F); Grice, 1972 (p.178, fig.F); Kos, 1972 (Vol. I, figsF, M, Rem.); Chen & Zhang, 1974 (p.103, figs.M); Bradford & Jillett, 1974 (p.6, figs.F); Björnberg & al., 1981 (p.616, figs.F,M); Zheng & al., 1982 (p.6, figs.M); Fleminger, 1985 (p.276, 285, Table 1, 4, fig.M, Rem.: A1); Bradford, 1988 (p.74, 76, 79, Rem.); Zheng Zhong & al., 1984 (1989) (p.227, figs.F,M); Nishida, 1989 (p.173, fig. 3, table 1, 2, 3: dorsal hump); Chihara & Murano, 1997 (p.740, Pl.68: F,M); Bradford-Grieve & al., 1999 (p.877, 908, figs.F,M); Miller, 2002 (p.133, 139, Rem.); Bucklin & al., 2003 (p.335, tab.2, fig. 2, Biomol.); Conway & al., 2003 (p.150, figs.F,M, Rem.); Machida & al., 2006 (p.1071, tab.1,3, fig.3,4, molecular phylogeny); Vives & Shmeleva, 2007 (p.904, figs.F,M, Rem.); Fornshell & Ferrari, 2014 (p.111, fig.8: Von Vaupel Klein's organ) | 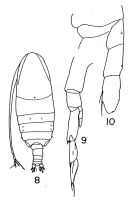 Issued from : T.S. Park in Fishery Bull. Fish Wild. Serv. U.S., 1968, 66 (3). [Pl.531, Pl.1, Figs.8-10]. Male: 8, habitus (dorsal); 9, left P5; 10, endopod of left P5. Nota: The distal segment of the left rudimentary endopod, in some specimens, has 2 or 3 small setae.
|
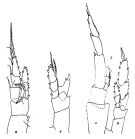 issued from : J.M. Bradford & J.B. Jillett in Crustaceana, 1974, 27 (1). [p.14, Fig.2,B,F,L]. Female: B, P1; F, P5. Male: L, P5 (L = left, R := right)
|
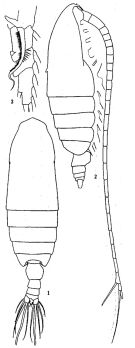 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.2, 1-3]. Female (from E China Sea): 1, habitus (dorsal); 2, idem (lateral right side); 3, endopod of P1 (anterior).
|
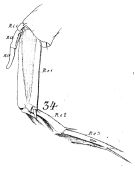 issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf.8, Fig.34]. As Calanus robustior. Male: left P5 (anterior surface).
|
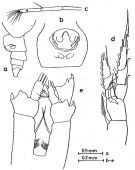 issued from : O. Tanaka in Publ. Seto Mar. Lab., 1969, XVII (4). [Fig.1]. Female (from off Peru): a, urosome (lateral left side); b, genital segment (ventral); c, distal part of A1; d, P1; e, P4 (incomplete). Nota Female: - A1 is abnormal in that the 18th segment is regenerated, carrying 9 setae at the distal margin.
- P4 has an abnormal endopod, 1-segmented instead 3 segments.
- Genital segment strongly swollen; there is a ridge-like structure behind the genital opening.
- P1 characteristic in structure.
|
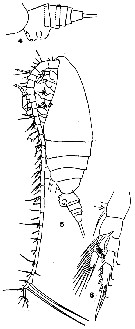
 issued from : Q.-c. Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.1, Figs.4-5]. Male (from China Seas): 4, habitus (dorsal); 5, P5.
|
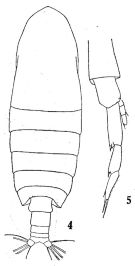
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.4, Figs.4-6]. As Calanus robustior. Female: 4, urosome (lateral); 5, habitus (lateral); 6, P1 (anterior).
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.3, Fig.11]. As Calanus robustior. Female: 11, P5 (anterior).
|
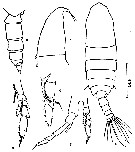 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.7, Fig.1]. Male: a, habitus (dorsal); b, head (lateral); c, urosome (lateral, right side); d, endopod of P1; e, P4; f, P5. Scale bar in mm.
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.129, Fig.5]. As Calanus robustior. Female (from San Diego Region): a, habitus (lateral); b, outer margin of 1st basipodal segment of Mx2; c, genital segment (lateral); d, endopod of P2. b2 = 2nd basipodite segment; R1.1 = 1st endopodite segment; Si = inner marginal bristle.
|
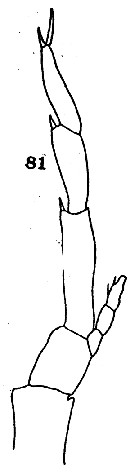 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1906, 3 (5). [Pl.14, Fig.81]. Male (from San Diego, California): 81, left P5.
|
 issued from : R.J. Machida, M.U. Miya, M. Nishida & S. Nishida in Mar. Biol., 2006, 148. [p.1077, Table 5]. Comparisons of selected biological characteristics of 6 species olus 1 subspecies of Neocalanus.
|
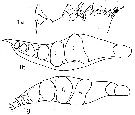 issued from : N.V. Vyshkvartzeva in Issled. Fauny Moreï, 1972, 12 (20). [p.168, Fig.7, a, b, g]. As .
Femele Md (masticatory edge): 1a, lateral aspect); 1b, left Md (above); 1g, right Md (above aspect).
|
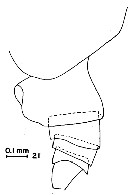 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.176, Pl.1, Fig.21]. Female (from equatorial Pacific): 21, posterior part of thorax and urosome (lateral, left side).
|
 issued from : A. Fleminger in Mar. Biol., 1985, 88. [p.284, Fig.6, F]. Male (from SE Indian): Left A1 proximal segments (ventral view); Nota: see remarks in Calanus s.l. pacificus californicus (Fleminger, 1985, p.275) concerning the dimorphism in the female A1.
|
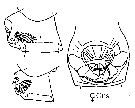 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.981, Fig. 7.21]. Female: genital segment (ventral and lateral, respetively). Nota: Genital segment very swollen ventrally. Left and right spermatheca enclosed under common cover.
|
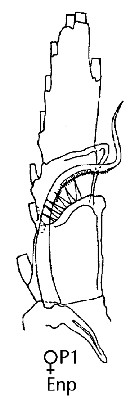 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.981, Fig. 7.21]. Female: Endopodite of P1. Nota: - Basis of P1 with large spine at base of inner seta. - Basis of P2-P5 naked.
|
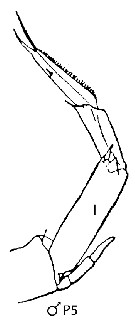 issued from : J.M. Bradford-Grieve, E.L. Markhaseva & C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, Edit. D. Boltovskoy. 1999. Vol.2. Copepoda. [p.981, Fig. 7.21]. Male: Left P5. Nota: Setation of endopodite of left (I) P5 highly reduced.
|
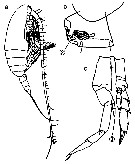 Issued from : M. Chihara & M. Murano in An Illustrated Guide to Marine Plankton in Japan, 1997. [p.747, Pl. 68, fig.86 a, c-d]. Female: a, habitus (lateral); c, posterior corner of last thoracic segment and genital segment. Male: d, P5. Nota: Numbers show species characteristics. Figs. c, d, after Brodsky, 1972.
|
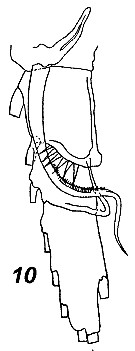 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.41, Fig.11, 10]. Neocalanus robustior after Brodsky, 1972. 10, distal part of basis and endopod.
|
 Issued from M. Rose in Résult. Camp. scient. Prince Albert I, 1929, 78, p.8. Remarques concernant Neocalanus robustior identifié aux cours des campagnes de 1904, 1911, et 1912.
|
 Issued from : J.A. Fornshell & F.D. Ferrari in Crustaceana, 2014, 87 (1). [p.111, Fig.1, A-C]. Von Vaupel Klein's organ on right endopodal segment 1 and right basal seta of P1. A, anterior, distal up, central lumen of attenuation indicated with dotted lines; B, right basal seta; C, proximal endopodal segment Scale line; 80 µm The Von Vaupel Klein organ is an association (srtucture) of the distal seta from basis and the proximal endopodal segment of P1. This structure shows significant variability among many gymnoplean copepods, in the shape of the distodorsal corner of the proximal endopodal segment, presence and location of denticles on the anterior face of the segment, presence and size of denticles along the distal margin of the segment , number of pores on the segment, shape of the seta that originates on the basis, and the morphology of the basis at the origin of the seta. The function of this complex structure is not known.
|
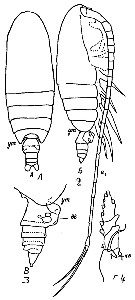 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. As Calanus (Neocalanus) robustior. After Brodsky, 1962. Female: 1-2, habitus (dorsal and lateral, respectively); 3, abdomen (lateral); 4, basis and endopodite of P1.
| | | | | Compl. Ref.: | | | Wilson, 1942 a (p.195); Sewell, 1948 (p.329, 400, 516, 520, 531, 545, 555, 566); C.B. Wilson, 1950 (p.269); Yamazi, 1958 (p.147, Rem.); Björnberg, 1963 (p.15, Rem.); De Decker & Mombeck, 1964 (p.13); Grice & Hulsemann, 1965 (p.223); Mazza, 1966 (p.69); 1967 (p.326); Grice & Hulsemann, 1968 (tab.2); Morris, 1970 (p.2300); Timonin, 1971 (p.281, trophic group); Roe, 1972 (p.277, tabl.1, tabl.2); Rudyakov, 1972 (p.886, Table 1: sinking rate); Björnberg, 1973 (p.283, 388); Vives & al., 1975 (p.35, tab.II, IV); Mullin & Evans, 1976 (p.119, chart distribution, length, sex ratio, vertical distribution); Deevey & Brooks, 1977 (p.256, Table 2, Station "S"); Tranter, 1977 (p.596); Carter, 1977 (1978) (p.35); Brenning, 1981 (p.1, Rem.: p.9);Vives, 1982 (p.190); Kovalev & Shmeleva, 1982 (p.82); Dessier, 1983 (p.89, Tableau 1, Rem., %); De Decker, 1984 (p.315); Guangshan & Honglin, 1984 (p.118, tab.); Brenning, 1985 a (p.28, Table 2); Madhupratap & Haridas, 1986 (p.105, tab.1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Lozano Soldevilla & al., 1988 (p.57); Heinrich, 1990 (p.16); Timonin, 1990 (p.479); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Shih & Young, 1995 (p.68); Suarez-Morales & Gasca, 1997 (p.1525); Alvarez-Cadena & al., 1998 (tab.3,4); Mauchline, 1998 (tab.8, 30, 64); Hure & Krsinic, 1998 (p.99); Suarez-Morales & Gasca, 1998 a (p108); Stephen, 1998 (p.341, 342, Rem., charts); Neumann-Leitao & al., 1999 (p.153, tab.2); Lapernat, 2000 (tabl.3, 4); Rebstock, 2001 (tab.2); 2002 (p.71, Table 3, 4, climatic variability); Uysal & al., 2002 (p.17, tab.1); Beaugrand & al., 2002 (p.179, figs.5, 6); Shimode & Shirayama, 2004 (tab.2: ?); Hsiao & al., 2004 (p.325, tab.1); Pusch & al., 2004 (251, tab.3); Gallienne & al., 2004 (p.5, tab.3); Lo & al., 2004 (p.89, tab.1); Prusova & Smith, 2005 (p.76); Mackas & al., 2006 (L22S07, Table 2); Hwang & al., 2007 (p.23); Albaina & Irigoien, 2007 (p.435: Tab.1); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.518); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Muelbert & al., 2008 (p.1662, Table 1); Licandro & Icardi, 2009 (p.17, Table 4); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Williamson & McGowan, 2010 (p.273, Table 3, Pacific central gyres: N and S, as robustior); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Mazzocchi & Di Capua, 2010 (p.424); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Teuber & al., 2013 (p.28, Table 1, respiration rates); Teuber & al., 2013 (p.1, Table 2, respiration rates); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Hirai & al., 2013 (p.1, Table I, molecular marker); Schukat & al., 2013 (p.1, Table 1, 2, fig.2, respiration, ingestion); Lidvanov & al., 2013 (p.290, Table 2, % composition); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2); Belmonte, 2018 (p.273, Table I: Italian zones); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Bode & al., 2018 (p.840, Table 1, respiration & ingestion rates, depth); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 17 | | |
|
Distribution map of Neocalanus robustior by geographical zones
|
| | | | | | | | | | | |  issued from : R.J. Machida, M.U. Miya, M. Nishida & S. Nishida in Mar. Biol., 2006, 148. [p.1077, Fig.4]. issued from : R.J. Machida, M.U. Miya, M. Nishida & S. Nishida in Mar. Biol., 2006, 148. [p.1077, Fig.4].
Relationship between the phylogeny and distribution pattern of the Neocalanus species.
Shaded portions of the maps represent distributions based on the litterature. Clades A-E correspond to those in Fig.3, p.1075. |
 issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998. issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998.
P.341, Fig.5: Distribution of Neocalanus robustior and Neocalanus gracilis in the Indian Ocean. |
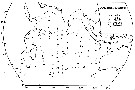 issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998. issued from : R. Stephen in Pelagic Biogeography ICoPB II. Proc. 2nd Int. Conf. Final report of SCOR/IOC working group 93. 9-14 July 1995. Workshop Rep. No.142. UNESCO, 1998.
P.342, Fig.6: Distribution of Neocalanus robustior in the Indian Ocean. |
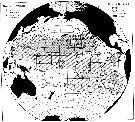 issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.120, Fig.1]. issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.120, Fig.1].
Distribution of Neocalanus robustior in the Pacific Ocean
Tows were taken from the surface to the depth indicared by open or filled circle. Contour intervals were chosen arbitrarily, based on breaks in the data abundances.
The copepodite stages III, IV, V, and adults were counts in aliquots (subsampled with a plankton splitter.The samples were taken during expeditions in May, Yune and August 1952; December-November 1953; September-November 1953; July-September 1955; August-September 1956; October-February 1957-1958; November 1960; June-August 1960; March-April 1960-1961; September 1964; January 1966 and March-May 1967. Most of the samples were taken by oblique tows to various depths (net: 500-550 µ mesh aperture, 1 m mouth diameter, with a flowmeter).
Nota: N. robustior is most abundant in the eastern Central North Pacific; it occurs only occasionally north of 35° N, and is rare or absent in the eastern tropical Pacific, the Peru-Chile Current, and the western South Pacific.
Compare with the distribution of N. gracilis for the same expeditions.. |
 issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.124, Table 1]. issued from : M.M. Mullin & P.M. Evans, 1976, 30 (2). [p.124, Table 1].
Cephalothorax length and mandibular width : Cephalothorax length, by area, of Neocalanus robustior and Neocalanus gracilis in the Pacific Ocean.
2-4 stations were pooled in each zoogeographic area. * No occurrences in samples examined.
Nota: After Mullin & Evans (1976, p.127), almost by definition, sympatric species, as suggested N. rosustior and N. gracilis, occupy different ecological niches ; of less certainty is the proposition that they must do so to avoid competitive exclusion and extinction of either. The latter statement may not be proven simply by the observation that ecological differences can usually be found between sympatric species, since the conclusion that these differences are sufficient to reduce competition to an acceptable level is invalid unless the extent and nature of the presumed competition have been demonstrated.
The pelagic environment seems to be relatively monotonous with respect to conventionally measured properties, although it may be rather more diverse from the zooplankters point of view. In any case, the plants are very small and the planktonic herbivores generally have no persistent spatial association with a particular species of food but must feed by filtration on a variety of particles.
In the two species of Neocalanus, as in Calanus (= Mesocalanus lighti and C. (Mesocalanus) tenuicornis (Mullin, 1969), the sizes of the mandibular blades of any developmental stage are more similar than the sizes of the bodies. If there had been strong evolutionary pressure for divergence in size of feeding appendages between sympatric competitors, one would expect the mandibular blades to be at least as dissimilar as the bodies, or more so. In fact, failure to diverge could mean that the congeners do not compete for a common, limiting source of food ; that divergence in selection of food is based of some property of food particles other than their size or on a morphological feature other than mandibular morphological feature other than mandibular width ; or it could simply be a consequence of continuous breeding and the considrable increase in bodily size which accompagnies juvenils development. Continuous breeding means that all developmental stages of both congeners are present in the water at any time.Hence, a slight increase in mandibular size of the larger congener may make it more distinct from the smaller at the same developmental stage, but at each stage, except that of the adult female the mandibular size of the larger congener would still be very similar to the next older stage of the smaller congener. Similarly, little advantage may be gained by a reduction in mandibular size of the smaller of two congeners as long as the juvenile stages of the larger are always present.
Vertical partitioning of the pelagic habitat has often been cited as a means by which related species can reduce competition (see in Baker, 1970 concerning euphausids : Pugh, 1974, siphonophores ; Clarke, 1973, myctophid fishes). Zalkhina (1970) found that the centers of vertical distribution in the upper 300 m of tropical, cyclopoid copepods were different and partial vertical separation of calanoids congeneric species of Metridia and Pleuromamma. The depths of maximal abundance of Calanus (= Mesocalanus) lighti andC. (= Mesocalanus) tenuicornis in the eastern Central North Pacific are distinct during at least part of the year. As in the presnt case, the species of smaller bodily size and greater geographical range tends to occur deeper in the water comumn. In both Calanus and Neocalanus, however, the overlap between congeners is considerable. |
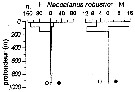 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.10 h]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.10 h].
Verical distribution of Neocalanus robustior at an oligotrophe site (off NW Cape Verde Islands: 21° N, 31° W) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
 Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1). Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1).
Cf. explanations of these measures in Calanoides natalis from the same authors.
Compare with Neocalanus gracilis. |
| | | | Loc: | | | South Africa (E), Namibia, off Angola, Congo, G. of Guinea, Dakar, Cape Verde Is., off NW Cape Verde Is., Great Meteor Seamount, Azores, Canary Is., Morocco-Mauritania, off Madeira, off Rio de La Plata (shelfbreak), Uruguay (continental shelf), Brazil, off Rio de Janeiro, Campos Basin, off Natal, off Amazon, Caribbean Colombia, Caribbean Sea, Jamaica, Yucatan, G. of Mexico, Florida, Sargasso Sea, Station "S" (32°10'N, 64°30'W), Bermuda, Gulf Stream (41°N, 58°W), Bay of Biscay, Medit. (Alboran Sea, Suidi Fredj coast, W basin (rare), off Barcelona, Ligurian Sea, S Adriatic Sea, Lebanon Basin, Alexandria, W Egyptian coast), Arabian Sea, Laccadive Is., Maldive Is., Rodrigues Is., Seychelles, Mascarene Basin, Minicoy Is., Indian, W Australia, Philippines, Pacif. (W equatorial), China Seas (East China Sea, South China Sea), Taiwan (S, E, N: Mienhua Canyon), Japan, off SE Japan, Okhotsk Sea, Aleutian Is., G. of Alaska, Bikini Is., Pacif. (central N), Pacific (central gyres: N and S), Hawaii, off Hawaii (E, NE), California, W Costa Rica, Clipperton Is., Galapagos, off Peru, Pacif. (SE tropicical), off SE Australes Is., off Tuamotu Is., off Juan Fernandez Is., Chile, Tasmania (in Taw, 1978) | | | | N: | 146 | | | | Lg.: | | | (11) F: 3,93-3,67; (14) F: 3,9-3,42; M: 2,84; (17) M: 3,3; (28) F: 3,8-3,55; M: 2,93; (35) F: 4,32-4,05; (47) F: 4-3,65; M: 3; (105) F: 4,65; (72) F: 4,41-4,03; M: 3,55-3,16; (73) F: 4,42-3,92; (101) F: 4,08-3,4; (125) M: 3; (142) F: 3,17; (150) F: 3,87-3,62; M: 3,07; (187) F: 4,12-4,09; (199) F: 4,1-3,42; M: 3,12; (237) M: 3,1; (290) F: 4,1-3,65; (321) F: 3,59; (338) M: 3,5-3,1; (866) F: 3,8-4,5; M: 3,1-3,6; (991) F: 3,17-4,41; M: 2,84-3,55; (1023) M: 3,53; (1230) F: 3,2-4,1; M: 2,8-3,0; {F: 3,17-4,65; M: 2,80-3,60} | | | | Rem.: | epi-mesopelagic. Sargasso Sea: 0-1500 m.
Numerous references under the designation of Calanus robustior. May be confused with N. gracilis, but of a larger size.
According to Björnberg (1963, p.15) this species is recorded for the first time in Brazilian waters in very small numbers.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a coarse-filter feeder.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 25/10/2022 | |
| | | | On the second figure, where it says "Female: B, P2" it is actually the Female P1.
| |
|
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 28, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
