|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Candaciidae ( Family ) |
|
|
|
Candacia ( Genus ) |
|
|
| |
Candacia bispinosa (Claus, 1863) (F,M) | |
| | | | | | | Syn.: | Candace bispinosa Claus, 1863 (p.191,figs.F,M); Giesbrecht, 1892 (p.424, 440, 771, figs.F,M); Thompson, 1888 d (p.140);
Candacia truncata : Brady, 1883 (p.69, fig.F);
Paracandacia bispinosa : Grice, 1963 (p.173, figs.F,M); Vervoort, 1965 (p.157, Rem.); Owre & Foyo, 1967 (p.95, figs.F,M); Fleminger, 1967 a (tabl.1); Park, 1968 (p.567, Rem.F,M); Corral Estrada, 1970 (p.201, figs.F,M); Kos, 1972 (Vol. I, figs.F, M, Rem.); Lawson, 1977 (p.71, tab.2,3,4,fig.3,5); Björnberg & al., 1981 (p.655, figs.F,M); Chihara & Murano, 1997 (p.755, Pl.79,80: F,M); Mulyadi, 1997 a (p.102, Redescr.F, figs.F); Lapernat, 1999 (p.12, 55, fig.F); Bradford-Grieve & al., 1999 (p.885, 955, figs.F,M); Mulyadi, 2004 (p.100, figs.F, Rem.); Avancini & al., 2006 (p.102, Pl. 71, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.461, figs.F,M, Rem.)
Ref. compl.: Grice & Hulsemann, 1965 (p.225); 1967 (p.19); Park, 1970 (p.478); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 c (p.1037); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Harding, 1974 (p.141, tab. 3, gut contents); Deevey & Brooks, 1977 (p.156, tab.2, Station "S"); Carter, 1977 (1978) (p.36); Haury & Wiebe, 1982 (p.915, horizontal micro-distribution); Vives, 1982 (p.294); Dessier, 1983 (p.89, Tableau 1, 2, Rem., %); Scotto di Carlo & al., 1984 (1044); Cummings, 1984 (p.163, Table 2); Guangshan & Honglin, 1984 (p.118, tab.); Regner, 1985 (p.11, Rem.: p.37); Madhupratap & Haridas, 1986 (p.105, tab.1); Ambler & Miller, 1987 (tab.2, 3); Lozano Soldevilla & al., 1988 (p.60); Ohtsuka & Kubo, 1991 (p.541); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Shih & Young, 1995 (p.69); Webber & Roff, 1995 (tab.1); Webber & al., 1996 (tab.1); Suarez-Morales & Gasca, 1997 (p.1525); Hure & Krsinic, 1998 (p.102); Alvarez-Cadena & al., 1998 (tab.1,4); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p109); Neumann-Leitao & al., 1999 (p.153, tab.2); Siokou-Frangou, 1999 (p.478); Lopez-Salgado & al., 2000 (tab.1); Haury & al., 2000 (p.69, Table 1); Lapernat, 2000 (tabl.3, 4); Lapernat & Razouls, 2001 (p.123, tab.1); Beaugrand & al., 2002 (p.179, figs.5, 6); Conway & al., 2003 (p.112, figs.F,M, Rem.); Hsiao & al., 2004 (p.325, tab.1); Pusch & al., 2004 (251, tab.3); ; CPR, 2004 (p.57, fig.168); Osore & al., 2004 (p.195); Berasategui & al., 2005 (p.313, fig.2); Hwang & al., 2007 (p.23); Dur & al., 2007 (p.197, Table IV); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); C.-Y. Lee & al., 2009 (p.151, Tab.2); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Hsiao S.H. & al., 2011 (p.475, Appendix I); Hsiao & al., 2011 (p.317, Table 2, fig.6, indicator of seasonal change); Hirai & al., 2013 (p.1, Table I, molecular marker)
Canadacia bispinosa : Go al., 1997 (tab.1, lapsus calami) | | | | Ref.: | | | Candacia bispinosa : Giesbrecht & Schmeil, 1898 (p.129, Rem. F,M); Thompson & Scott, 1903 (p.235, 250); A. Scott, 1909 (p.151, Rem.); Wolfenden, 1911 (p.357); Pesta, 1920 (p.538); Sars, 1925 (p.352); 1925 (p.152); 1929 (p.43); Farran, 1929 (p.210, 272); Sewell, 1932 (p.334); Rose, 1933 a (p.255, figs.F,M); Tanaka, 1935 a (p.214, figs.F,M); Farran, 1936 a (p.114); Mori, 1937 (1964) (p.84, figs.F); Wilson, 1942 a (p.174; fig.juv.F); Lysholm & al., 1945 (p.41); Sewell, 1947 (p.246); 1948 (p.391, 346, 396, 408, 412, 414, 433, 443, 450, 457, 460, 470); Moore, 1949 (p.60); C.B. Wilson, 1950 (p.181); Chiba & al., 1957 a (p.11); Fagetti, 1962 (p.33); Grice, 1962 (p.235, figs.F,M, Rem.); Brodsky, 1962 c (p.140, figs.F); Duran, 1963 (p.24); V.N. Greze, 1963 a (tabl.2); Crisafi, 1963 (p.101, figs.F,M, juv.); De Decker & Mombeck, 1964 (p.11); Tanaka, 1964 c (p.245) and authors before 1963: cf. in Vervoort, 1965 (p.157); El-Maghraby, 1965 (p.56, Appendix); Pavlova, 1966 (p.44); Mazza, 1966 (p.72); 1967 (p.324, 367); Séguin, 1968 (p.488); Vinogradov, 1968 (1970) (p.268, 269); Dowidar & El-Maghraby, 1970 (p.267); Deevey, 1971 (p.224); Apostolopoulou, 1972 (p.328, 365); Tranter, 1977 (p.596); Frontier, 1977 a (p.16); Kovalev & Shmeleva, 1982 (p.85); Zheng & al., 1982 (p.70, figs.F); De Decker, 1984 (p.317) ; Padmavati & al., 1998 (p.347); Barthélémy & al., 1998 (p.721, genital area); Barthélémy, 1999 a (p.9, Fig.12, C); Moraitou- Apostolopoulou & al., 2000 (tab.I); Gallienne & al., 2004 (p.5, tab.3); Boxshall & Halsey, 2004 (Rem. p.84); | 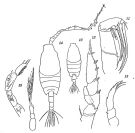 issued from : O. Tanaka in Suisan Gakkai Ho, Tokyo, 1935, 6 (4). [Pl.VI, p.227]. Female: 10, habitus (dorsal view); 11, Mx2; 12, P3; 13, P5. Male: 14, habitus (dorsal view); 15, P5 (left and right)
|
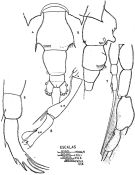 issued from : J. Corral Estrada in Tesis Doct., Univ. Madrid, A-129, Sec. Biologicas, 1970. [Lam.53, figs.4-8]. As Paracandacia bispinosa. Female (from Canarias Is.): 4, urosome (dorsal); 5, idem (lateral right side); 6, P5. Male: P5; 8, right A1 (grasping segments).
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.70, Fig.40]. Female: a, habitus (dorsal); b, urosome (ventral); c, exopodite segment 3 of P3; d, P5. Scale bar in mm.
|
 issued from : T. Mori in The Pelagic copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl. 56, Figs.9-14]. Female: 9, P3; 10, habitus (dorsal); 11, A1; 12, Mx2; 13, P5; 14, P1.
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.236, Pl.32, Figs.7-14]. Female (from equatorial Pacific): 7, habitus (dorsal); 8, posterior part of thorax and urosome (right side); 9, Md (biting edge); 10, Mx2; 11, Mxp; 12, P5. Two spine-like protrusions arising from the genital segment; these protrusions, particularly the one on the left side, are quite variable in shape and size. In one specimen the left protrusion reaches nearly to the anal segment; there is a small rounded knob arising from the ventral side of the genital segment near the posterior end (fig.8. The preanal segment is asymmetrical. The left side is dilated near the anterior end, but there is some variation in the size of the protrusion. The ventral surface of this segment may or may not have a row of hairs arising from near the posterior end. The basal tooth of Md is undivided, but one or more spine-like processes are present on this tooth The large terminal finger-like protrusion of the distal segment of P5 is finely serrate on both sides. Male: 13, segments 16 to 21 of right A1; 14, P5. Nota: Segment 20 of right A1 swollen distally. There are 3 outer edge spines on the distal segment of right P5; the proximal spine is shorter than the distal two spines; the penultimate segment of left P5 iis enlarged and the outer edge seta of the distal segment is shorter than the terminal ones.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.101, Fig.56]. As Paracandacia bispinosa. Female (from 07°29'S, 121°05'E): A, habitus (dorsal); B, posterior part of last thoracic segment and urosome (lateral right side); C, urosome (ventral); D, Mx2; E, P3; F, P5.
|
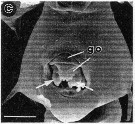 issued from : R.-M. Barthélémy in Thèse Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.12, C]. Female (from Tripoli coast, E Mediterranean Sea): B, external ventral view genital double-somite. go = genital operculum; Note the complex operculum with two lateral lobes (arrowed), and the multilobed medial part and ornamented of mico-folds. Scale bar: 0.050 mm.
|
 issued from : R.-M. Barthélémy in Thèse Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.12, F, G]. Female: F, G, external ventral view genital double-somite (details genital area). go = genital operculum; Note the complex operculum, with multilobed operculum medial part (arrowed) and ornamented of micro-folds (thin arrows); lateral expansions (thick arrows) in continuated with medial part and the posterior pad (asterisk). Scale bars: 0.010 mm.
|
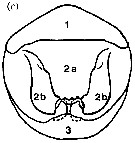 issued from : R.-M. Barthélémy, C. Cuoc, D. Defaye, M. Brunet & J. Mazza in Phil. Trans. R. Soc. Lond., B, 1998, 353. [p.729, Fig.54 c]. c, Schematic external genital area Candacia bispinosa. Nota: Compare with C. simplex, C. armata and C. ethiopica. Homologous structures have the same number: 1, anterior thickening of genital operculum; 2a, median part of genital operculum; 2b, lateral expansions of genital operculum; 3, posterior pad.
|
 issued from : R.-M. Barthélémy, C. Cuoc, D. Defaye, M. Brunet & J. Mazza in Phil. Trans. R. Soc. Lond., B, 1998, 353. [p.733, Fig.63]. Schematic representation of external genital area in the species studied; Dashed line, limit of the anterior pad and lateral thickenings; shading, posterior pad; dots, genital operculum.
|
 Issued from : J.M. Bradford-Grieve, E.L. Markhaseva, C.E.F. Rocha & B. Abiahy in South Atlantic Zooplankton, edit. D. Boltovskoy. 1999, Vol. 2, Copepoda; [p.1066, Fig. 7.375: Candacia Paracandacia bispinosa (= Candacia bispinosa)]. Gns = genital segment; r = right antennula. Female characters (from key, p.955): - P5 terminal segments end in processes at least one of which is finger-like process and may be finely serrated on one or both margins. - In dorsal view, genital segment with spine-like protrusions arising from each side and directed posteriorly. Male (from key, p.925): - P5 not chelate on right but ends in long plumose seta; right A1 has no teeth in geniculate region. - One or both of right A1 segments 16 and 19-20 with rounded or elongate protrusions. - A1 segment 16 with rounded protrusion distally, segment 19-20 produced distally.
|
 Issued from : W. Giesbrecht in Fauna Flora Golf. Neapel, 1892, 19. [Taf. 39, figs. 15, 16, 17, 20]. As candace bispinosa. Female: 15, urosome (ventral); 16, thoracic segments and urosome (lateral); 17, urosome (ventralà; 20, habitus (dorsal).
|
 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. As Paracandacia bispinosa. From Giesbrecht, 1892 and others, after Brodsky, 1962. Female: 1, habitus (dorsal); 2, ggenital segment and abdominal segment (dorsal); 3, abdomen (ventral); 4-5, abdomen lateral (left and right view, respectively); 6, P5. Male: 7, P5.
| | | | | Compl. Ref.: | | | Yamazi, 1958 (p.151, Rem.); Heinrich, 1960 (p.36); 1961 (p.90); Gaudy, 1962 (p.93, 99, Rem.: p.113) ; Björnberg, 1963 (p.55, Rem.); Timonin, 1971 (p.281, trophic group); Guglielmo, 1973 (p.399); El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); McKinnon & al., 2008 (p.843: Tab.1); Dias & al., 2010 (p.230, Table 1); Williamson & McGowan, 2010 (p.273, Table 3, Pacific central gyres: N and S); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Mazzocchi & Di Capua, 2010 (p.424); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Uysal & Shmeleva, 2012 (p.909, Table I); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, pers. comm.); Lidvanov & al., 2013 (p.290, Table 2, % composition); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Mazzocchi & al., 2014 (p.64, Table 4, abundance) ; Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 16 | | |
|
Distribution map of Candacia bispinosa by geographical zones
|
| | | | | | | | | | | |  issued from : T.J. Lawson in Marine Biology, 1977, 43. [Fig.5, p.78]. As Paracandacia bispinosa. issued from : T.J. Lawson in Marine Biology, 1977, 43. [Fig.5, p.78]. As Paracandacia bispinosa.
Distribution map for the Indian Ocean. |
 issued from : Mulyadi in Treubia, 1997, 31 (2). [p.109, Fig.16]. issued from : Mulyadi in Treubia, 1997, 31 (2). [p.109, Fig.16].
Distribution of Candaciidae in Indonesian waters. 12: C. bispinosa. |
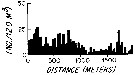 issued from : L.R. Haury & P.H. Wiebe in Deep-Sea Res., 1982, 29 (7A). [p.918, Fig.1, A]. issued from : L.R. Haury & P.H. Wiebe in Deep-Sea Res., 1982, 29 (7A). [p.918, Fig.1, A].
Abundance horizontal distribution of Paracandacia (= Candacia) bispinosa: non-random spatial structure.
Samples collected with a Longhurst-Hardy Plankton Recorder (LHPR) in the northern Sargasso Sea (34°10'N; 71°35'W) at three depth stata.
Nota: The pattern reveal that dominant variations occurred on scales of tens to hundreds of meters. The fluctuations in abundance are evident on different scales simultaneously.
After the authors, the abundance of species was not significantly nor consistently correlated to either temperature or alinity on the scales resolved by the tows, thus the LHPR data indicate that the physical environment or depth changes along the horizontal tows, was not, in general, a direct causative factor for the patterns observed. |
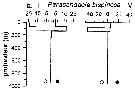 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.10 b]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.10 b].
Verical distribution of Paracandacia bispinosa (= Candacia bispinosa) at an oligotrophe site (off NW Cape Verde Islands: 21° N, 31° W) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
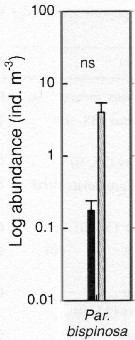 Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6]. Issued from : S.-H. Hsiao, S. Kâ, T.-H. Fang & J.-S. Hwang inHydrobiologia, 2011, 666. [p.326, Fig.6].
Variations in the most abundant copepod species (mean ± SE) along the transect in the boundary waters between the northern part Taiwan Strait and the East China Sea i March (black bar) and October (grey bar) 2005 (Mann-Whitney U test, sig. ns: P >0.05.
See drawings of Hydrological conditions and superficial marine currents in Calanus sinicus. |
| | | | Loc: | | | South Africa (E), off S St. Helena Is., off S Ascension Is., Congo, off St. Paul Is. (E & NE), off NW Cape Verde Is., off Morocco-Mauritania, Great Meteor Seamount, Canary Is., off Madeira, SW Azores, Argentina, Brazil, Bahia, off N Trindade Is., off Natal, off Amazon, Caribbean Colombia, Caribbean Sea, Jamaica, Yucatan, G. of Mexico, Cuba, Florida, Sargasso Sea, off Bermuda: Station S (32°10N, 64°30W), off E Cape Cod, Medit. (Alboran Sea, NW Basin, Marseille (very rare), Ligurian Sea, Naples, Milazzo, Messine, NW Tunisia, off Malta, Adriatic Sea, Ionian Sea, Aegean Sea, Lebanon Basin, Egyptian coast, Alexandria, Port Said), Sharm El-Sheikh, Red Sea, Arabian Sea, off Kenya, off SE Madagascar, Nosy Bé, Mascarene Basin, Maldives Is., Natal, Indian, Bay of Bengal, Australia (W, North West Cape), Indonesia-Malaysia, Flores Sea, Philippines, China Seas (East China Sea, South China Sea), Taiwan Strait, Taiwan, Kuroshio Current, Okinawa, Japan, Izu region, Tanabe Bay, Aleutian Is., Bering Sea, Bikini Is., off California, Pacif. (equatorial), Pacif. (W equatorial), Pacific (central gyres: N and S), Australia (Great Barrier), New Zealand, Hawaii, off NE Hawaii, Pacif. (N central), W Panama, W Colombia, Galapagos, off Chile | | | | N: | 148 (S Atlant.: 12; N Atlant.: 42; Medit.: 34; Red Sea: 4; Indian; 20; Indonesia-Malaysia: 4; Pacif.: 30) | | | | Lg.: | | | (16) F: 1,9-1,8; M: 1,7; (34) F: 1,72-1,56; M: 1,76-1,68; (46) F: 1,95-1,7; M: 2-1,75; (72) F: 2,01-1,74; M: 2,11-1,88; (101) F: 1,8-1,56; M: 1,73-1,56; (120) F: 1,79; M: 1,36; (150) F: 1,87; (180) F: 1,9; M: 2,17-2,13; (199) F: 1,9-1,67; M: 2,05-1,75; (237) F: 1,75-1,95; M: 1,75-1,90; (340) F: 1,9-1,85; (342) F: 2,163-1,957; M: 2,196-1,924; (449) F: 1,95-1,7; M: 2-1,75; (778) F: 1,8-1,72; (1023) F: 1,93; (1122) F: 1,75; (1230) F & M: 1,7-2,0; {F: 1,560-2,163; M: 1,360-2,190}
The mean female size is 1.826 mm (n = 27; SD = 0.1370), and the mean male size is 1.858 mm (n = 20; SD = 0.2175). The size ratio (male : female) is approximately 1.015. | | | | Rem.: | epi-mesopelagic; Sampling depth (off Malta): 2000-3000 m. Overall Depth Range in Sargasso Sea: 0-500 m (Deevey & Brooks, 1977, Station "S"). In vertical tow 4000-3000 m (Harding, 1974, Rem.: possible contaminant species from higher up the water column).
For Grice (1962, p.235) this species, C. simplex Giesbrecht and C. truncata Dana, differ in a few details. It has been suggested by Sars (1903) that these three species may have to be removed from the genus Candacia.
For Acha & al. (2020) this species is observed on Northern shelfbreak in front of Rio de La Plata (Argentina).
After Lapernat & Razouls (2002, p.19) the Itoh's index value = 4347.8 (number of teeth: 2).
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a piercing and sucking carnivorous.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 2 corresponding to small ambush feeding carnivorous.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 03/12/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 17, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
