 List of the studied authors
List of the studied authors
Yamazi, 1958 (p.149, Rem.); Minoda, 1958 (p.253, Table 1, abundance); M.W. Johnson, 1961 (p.311, Table 2); Northcote & al., 1964 (p.1069, Table II); Motoda, 1966 (p.260); Shih & al., 1971 (p.146, 203); Peterson & Miller, 1975 (p.642, 650, Table 3, interannual abundance); Kasahara & al., 1975 (p.25, eggs, cycles); Peterson & Miller, 1977 (p.717, Table 1, seasonal occurrence); Hirota & Uno, 1977 (p.77, fig. egg, seasonal abundance); Hanaoka, 1977 (p.267, 300, abundance); Gibson & Grice, 1977 (p.85, Table 1, copper pollution); Peterso & al., 1979 (p.467, Table 1); Grice & Marcus, 1981 (p.125, Dormant eggs, Rem.: p.133, 135, 137); Hirota 1981 (p.19, Table 1, length-weight-CHN); Uye S-i., 1982 (p.149, relation length-weight-C-N); Mackas & Sefton, 1982 (p.1173, Table 1); Yamamoto & Nishizawa, 1986 (p.1729, horizontal distribution); Hirakawa, 1986 (p.296,
Rem.); Mackas & Anderson, 1986 (p.115, Table 2); Nagasawa, 1987 (p.101,
Zoothamnium attached); Springer & al., 1989 (p.359, Fig.5), Coyle & al., 1990 (p.763); Hirakawa & al., 1990 (tab.3); Shih & Marhue, 1991 (tab.3); Hattori, 1991 (tab.1, Appendix); Yoo, 1991 (tab.1); Kimmerer, 1993 (tab.2); Hwang & Choi, 1993 (tab.3); Myung & al., 1994 (tab.1); Liang & al., 1994 (p.131, egg production); Uye & Kaname, 1994 (p.43, length v.s. fecal pellet volume); Shih & Young, 1995 (p.69); Madhupratap & al., 1996 (p.77, Table 2: resting eggs); Kotani & al., 1996 (tab.2); Liang & al., 1996 (p.527, population dynamics, production); Marcus, 1996 (p.143); Park & Choi, 1997 (Appendix); Liang & Uye, 1997 (p.415, population dynamic, production); Mauchline, 1998 (p.506, tab.25, 33, 35, 36, 40, 45, 47, 48, 58, 78); Suarez-Morales & Gasca, 1998 a (p.109); Gomez-Gutiérrez & Peterson, 1999 (p.637, Table I, II, III, VI, figs.4, 5, 6, 7, abundance, egg production); Bragina, 1999 (p.196); Pinchuk & Paul, 2000 (p.4, table 1, % occurrence); ; Peterson & al., 2002 (p.353); Peterson & al. 2002 (p.381, Table 2, interannual abundance); Peterson & Keister, 2003 (p.2499, interannual variability); Keister & Peterson, 2003 (p.341, Table 1, 2, abundance, cluster species vs hydrological events); Shimode & Shirayama, 2004 (tab.2); Park, W & al., 2004 (p.464, tab.1); Kasyan, 2004 (p.105); Hooff & Peterson 2006 (p.2610); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño); Deibel & Daly; 2007 (p.271, Table 4, Rem.: Arctic polynyas); Gaudy & Thibault-Botha, 2007 (p.84, Tab.2, Rem.: metabolism); Kang J.-H. & al., 2007 (p.82, fig.3, temporal variation vs. temperature & predation effect); Humphrey, 2008 (p.83: Appendix A); Liu & Hopcroft, 2008 (p.931: Table IV); Ohtsuka & al., 2008 (p.115, Table 5); Coyle & al., 2008 (p.1775, Table 5, abundance): Peterson, 2009 (p.73, Rem.: p. 78, indicator of long-term changes); Hopcroft & al., 2009 (p.9, Table 3, Fig.21, 24); Galbraith, 2009 (pers. comm.); Hopcroft & al., 2010 (p.27, Table 1, 2, fig.9); Bucklin & al., 2010 (p.40, Table 1, molecular biology); Keister & al., 2011 (p.2498, interannual variation); Batten & Walne, 2011 (p.1643, Table I, abundance vs temperature interannual variability); Matsuno & al., 2011 (p.1349, Table 1, abundance vs years); Beltrao & al., 2011 (p.47, Table 1, density vs time, as
C. abdominales); Matsuno & al., 2012 (Table 2); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Volkov, 2012 (p.474, Table 6, fig. 6, 12, abundance, distribution, interannual variation); Lavaniegos & al., 2012 (p. 11, Appendix); Lee J. & al., 2012 (p.95, Table 2: abundance, annual & seasonal distribution); Lee D.B. & al., 2012 (p.88, co-occurrence); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Moon & al., 2012 (p.1, Table 1); Takahashi M. & al., 2012 (p.393, Table 2, water type index); Li C. & al., 2013 (p.101, feeding); Ohashi & al., 2013 (p.44, Table 1, Rem.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Questel & al., 2013 (p.23, Table 3, interannual abundance & biomass, 2008-2010); Suzuki, K.W. & al., 2013 (p.15, Table 2, 3, 4, estuaries, annual occurrence); Oh H-J. & al., 2013 (p.192, Table 1, occurrence); Eisner & al., 2013 (p.87, Table 3: abundance vs water structure); Barton & al., 2013 (p.522, Table 1: metabolism, diapause, population dynamic, feeding mode, biogeo); Coyle & al., 2014 (p.97, table 3, 10, 12); Chiba S. & al., 2015 (p.968, Table 1: length vs climate); Zakaria & al., 2016 (p.1, Table 1, 3, Rem.); Ohtsuka & Nishida, 2017 (p.565, Table 22.1, Fig.22.3: central station in Japanese waters).


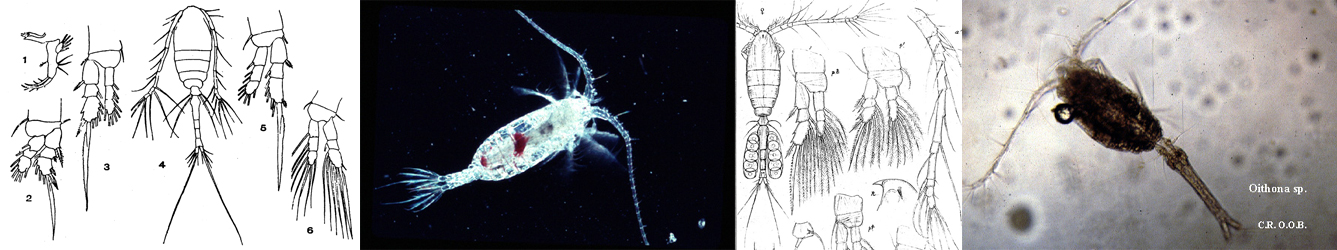

;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}