|
|
 |
|
Calanoida ( Order ) |
|
|
|
Diaptomoidea ( Superfamily ) |
|
|
|
Centropagidae ( Family ) |
|
|
|
Centropages ( Genus ) |
|
|
| |
Centropages furcatus (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Catopia furcata Dana,1849;
Centropages lenucculari Oliveira,1945;
? no Manaia velificata Oliveira, 1946 (1947) (p.466);
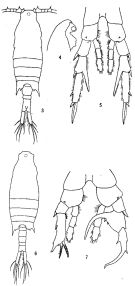
no Centropages furcatus : Björnberg, 1963 (p.42) | | | | Ref.: | | | Brady, 1883 (p.83, figs.F,M); Giesbrecht, 1892 (p.304, 320, 771, figs.F,M); Giesbrecht & Schmeil, 1898 (p.56); T. Scott, 1894 b (p.77); Giesbrecht & Schmeil, 1898 (p.56, Rem. F,M); Thompson & Scott, 1903 (p.234, 246); Wolfenden, 1905 (1906) (p.1016, Rem.F,M); Carl, 1907 (p.8, figs.M); A. Scott, 1909 (p.113, Rem.); Carl, 1907 (p.8, figs.M, Rem.); Wolfenden, 1911 (p.356); Sewell, 1912 (p.353, 360); Pesta, 1912 a (p.46, figs.F,M); 1913 (p.32); Sewell, 1914 a (p.220); Früchtl, 1924 b (p.45); Gurney, 1927 (p.150); Farran, 1929 (p.209, 255); Sewell, 1932 (p.229); 1933 (p.26); Dakin & Colefax, 1933 (p.205); Farran, 1936 a (p.108); Mori, 1937 (1964) (p.63, figs.M, Rem.F,M); Dakin & Colefax, 1940 (p.91, figs.F,M); Pesta, 1943 (p.21, figs.F,M); Marques, 1951 a (p.13); 1955 (p.18); Chiba & al., 1957 (p.308); 1957 a (p.11); Marques, 1958 (p.215); 1966 (p.7); 1976 (p.993); 1982 (p.761); Carvalho, 1952 a (p.145, figs.F, M); Brodsky, 1962 c (p.130, figs.F); Fish, 1962 (p.15); Tanaka, 1963 (p.9, Rem.F,M); Gaudy, 1963 (p.25); Kasturirangan, 1963 (p.31, figs.F,M); Vervoort, 1964 b (p.308); 1965 (p.88, Rem.); Gonzalez & Bowman, 1965 (p.247, figs.F,M, Rem.); Chen & Zhang, 1965 (p.77, figs.F,M); Marques, 1966 (p.7); Ramirez, 1966 (p.14, figs.F,M); Neto & Paiva, 1966 (p.25); Saraswathy, 1966 (1967) (p.77); Owre & Foyo, 1967 (p.74, figs.F,M); Koga, 1968 (p.17, fig.: egg); Björnberg, 1972 (p.32, figs.N, Rem.N); Kos, 1972 (Vol. I, figs.F0, M, Rem.); Lee, 1972 (p.10, figs.F,M, spermatophore and coupling device); Fleminger & Hulsemann, 1973 (p.344, figs.F,M, Rem., carte); Goswami & Goswami, 1974 (p.109, fig. caryotypes); Fleminger, 1975 (p.395, 397); Marques, 1976 (p.993); Greenwood, 1977 (p.61, figs.M); Marques, 1982 (p.761); Zheng Zhong & al., 1984 (1989) (p.244, figs.F,M); Sazhina, 1985 (p.60, figs.N); Chihara & Murano, 1997 (p.766, Pl.82,85: F,M); Mulyadi, 1998 (p.60, figs.F,M); Bradford-Grieve & al., 1999 (p.884, 951, figs.F,M); Bradford-Grieve,1999 b (p.139, figs.F,M, figs.180, 192); Al-Yamani & Prusova, 2003 (p.55, figs.F,M); Conway & al., 2003 (p.117, figs.F,M, Rem.); Mulyadi, 2004 (p.128, figs.F,M, Rem.); Phukham, 2008 (p.69, figs.F,M); Al-Yamani & al., 2011 (p.42, 44, figs.F,M) | 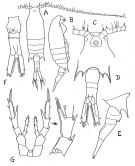 issued from : Brodsky K.A. in Issled. dal'nevost. Morei SSSR, 1962, 8. [Fig.32, p.131]. Female: A, habitus (dorsal view); B, idem (lateral view); C, head (ventral view); D, Th5 and urosome (dorsal view); E, Th5 and urosome (lateral view); F, urosome (ventral view); G, P5.
|
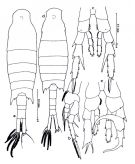 issued from : Mulyadi in The Raffles Bull. Zool., 1998, 46 (1). [Fig.4, p.61]. Female: a, habitus (dorsal view); b, P1; c, P5. Nota: A1 24-segmented, reaching distal end of anal somite; anterior margin of segments 1, 2 and 5 produced into 1 sharp teeth. Genital complex produced into triangular lobes on either side and into a rounded ventral lobe on the right side; urosomal segment 2 very short; urosomal segment 3 asymmetrical, twice as long as urosomal segment 2. Caudal rami relatively slender, asymmetrical, right ramus slightly longer than the left one. Male: d, habitus (dorsal view); e, P5. Nota: Right A1 geniculate, anterior margin of segments 15-16 with very small row of teethAnal somite well developed, as long as urosomal segment 3. Caudal rami 3 times as long as anal somite
|
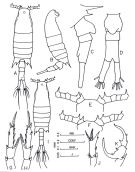 issued from : Al-Yamani F.Y. & Prusova I. in Common Copepods of the Northwestern Arabian Gulf: Identification Guide. Kuwait Institute for Scientific Research, 2003. [Fig.19, p.56]. Female: A, habitus (dorsal view); B, idem (right lateral view); C, urosome (right lateral view); D, urosome (ventral view); E, anterior part of head and A1 (dorsal view); G, left exopodite of P5; H, right exopodite of P5. Male: I, habitus (dorsal view); F, anterior part of head and A1 (dorsal view); K, right exopodite of P5; J, left exopodite of P5.
|
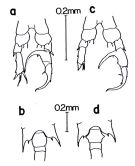 issued from : Fleminger A. & Hulsemann K. In: The Biology of the Indian Ocean, B. Zeitzchel Ed., 1973. [Fig. 6, p.345]. Comparison between C. furcatus and C. velificatusC. furcatus : a, Male (P5, posterior view); b, Female (fifth thoracic segment and genital segment, dorsal view); specimens from Gulf Panama (off Panama City). C. velificatus : Male (P5, posterior view); d, Female (fifth thoracic segment and genital segment, dorsal view); specimens from Gulf of Mexico (off Galveston).
|
 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.28, 3-7]. Female (from E China Sea): 3, habitus (dorsal); 4, forehead (lateral); 5, P5 (posterior). Male: 6, habitus (dorsal); 7, P5 (posterior).
|
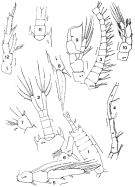 issued from : F.C. Ramirez in Bol. Inst. Biol. Mar., Mar del Plata, 1966, 11. [Lam.III, Figs.1-12]; Male (from off Mar del Plata): 1, habitus (dorsal); 2, urosome (dorsal); 3, proximal portion of right A1; 4, right A1 (segments 17 to 19); 5, P3 (exopodite); 6, right P4; 7, left P4; 8, terminal spine of exopodal segment of P4; 9, right P5; 10, left P5; 11, posterior part cephalothorax and urosome (dorsal); 12, P5.
|
 issued from : Mori T. in The pelagic Copepoda from the neifhbouring waters of Japan. S. Shirai ed., Tokyo, 1937; 2nd edit, 1964. [Pl.32, Figs.1-2]. Male: 1, P5; 2, habitus (dorsal).
|
 issued from : J.G. Greenwood in Proc. R. Soc. Qd, 1977, 88. [p.62, Fig.5, a-b]. Male (from 27°20'S, 153°15'E): a, P5 (posterior view with endopods omitted); b, P5 (anteerior view).
|
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.20, Fig.71]. Male: 71, right P5.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.18, Fig.13]. Female: 13, A1 (proximal segments (dorsal view).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.17, Fig.50]. Female: 50, exopod segments 2 and 3 of P5.
|
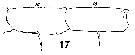 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.18, Fig.17]. Male: 17, segments 15 and 16 of right A1.
|
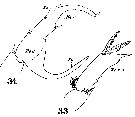 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.17, Figs.33, 34]. Male: 33, exopod of left P5 (posterior view); 34, exopod of right P5 (posterior view).
|
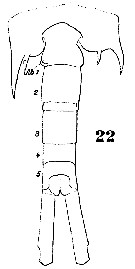 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.38, Fig.22]. Male: 22, last thoracic segment and urosome (dorsal).
|
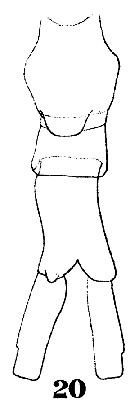 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.38, Fig.20]. Female: 20, urosome (ventral).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.38, Fig.15]. Female: 15, forehead (lateral).
|
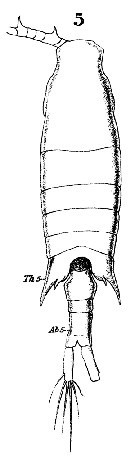 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.38, Fig.5]. Female: 5, habitus (dorsal).
|
 issued from : N. Phukham in Species diversity of calanoid copepods in Thai waters, Andaman Sea (Master of Science, Univ. Bangkok). 2008. [p.151, Fig.25]. Female (from W Malay Peninsula): a, habitus (dorsal); b, P5. Male: c, habitus (dorsal); d, P5. Body length after the drawings: F = 1.617 mm; M = 1.565 mm.
|
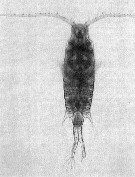 Issued from : I. Siokou & al. in Medit. Mar. Sci., 2013, 14/1. [p.246, Fig.11] Centropages furcatus female in the Aegean Sea (39°26'N, 25°33'E). (photo by C. Frangoulis).
|
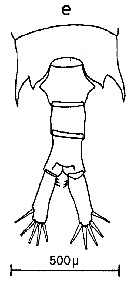 Issued from : C.M. Lee in Bull. mar. Ecol., 1972, 8 (1). [p.4, Text-fig.1,e]. Dorsal view of the fifth metasome segment and urosome of female of Centropages furcatus (from Tema and Takoradi, West Africa).
|
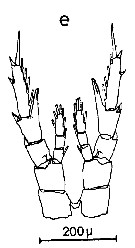 Issued from : C.M. Lee in Bull. mar. Ecol., 1972, 8 (1). [p.5, Text-fig.2 e]. P5 female of Centropages furcatrus.
|
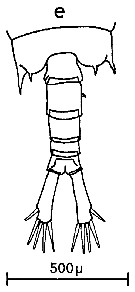 Issued from : C.M. Lee in Bull. mar. Ecol., 1972, 8 (1). [p.4, Text-fig.1 e]. Dorsal view of the fifth metasome segment and urosome of male of Centropages furcatus (from Tema and Takoradi, West Africa).
|
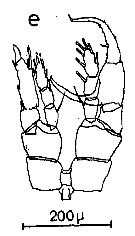 Issued from : C.M. Lee in Bull. mar. Ecol., 1972, 8 (1). [p.5, Text-fig.2 e]. P5 male of Centropages furcatrus.
|
 Issued from : J. Carl in Rev. Suisse Zool., 1907, 15. [Pl. I, figs.6, 7]. Centropages furcatus Male (from Baie d'Amboine = Ambon Bay, Indonesia): 6, Right P5, Re2 and Re3 (posterior surface; Re: exopod); 7, Left P5 (ending of exopodite).
| | | | | Compl. Ref.: | | | Cleve, 1904 a (p.187); Carl, 1907 (p.16, figs.M); Gurney, 1927 (p.150); Wilson, 1942 a (p.177); Sewell, 1948 (p.322, 325); C.B. Wilson, 1950 (p.186); Krishnaswamy, 1953 (p.117); Deevey, 1956 (tab. IV); Grice, 1956 (p.52); Yamazi, 1958 (p.149, Rem.); Kott, 1957 (p.5, 8); Deevey, 1960 (p.5, Table II, annual abundance, Rem.: fig.18, 19); Fagetti, 1962 (p.29); Cervigon, 1962 (p.181, tables: abundance distribution); Ganapati & Shanthakumari, 1962 (p.8, 15); Wickstead, 1962 (p.546, food & feeding); Gaudy, 1963 (p.25, Rem.); De Decker, 1964 (p.15, 20, 28); De Decker & Mombeck, 1964 (p.11); Neto & Paiva, 1966 (p.25, Table III); Fleminger, 1967 a (tabl.1); Delalo, 1968 (p.138); Itoh, 1970 (tab.1); Timonin, 1971 (p.281, trophic group); Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night, Table II: %, Table IV); Binet & al., 1972 (p.71); Brodsky, 1972 (p.256); Heinrich, 1973 (p.95); Björnberg, 1973 (p. 385); Patel, 1975 (p.659); Tranter, 1977 (p.598); Carter, 1977 (1978) (p.36); Binet, 1979 (p.400); Dessier, 1979 (p.89, 201, 205); Stephen & Iyer, 1979 (p.228, tab. 1, 3, 4, figs.3, 4); Paffenhöfer & Knowles, 1979 (p.35, fecal pellets); Chen Q-c, 1980 (p.794); Paffenhöfer & Knowles, 1980 (p.355, feeding); Sreekumaran Nair & al., 1981 (p.493); Madhupratap & al., 1981 (p.266, fig.1f: abundance vs. geographic transect); Vives, 1982 (p.293); Dessier, 1983 (p.89, Tableau 1, 2, Rem., %); Turner & Dagg, 1983 (p.16); Dagg, 1983 (p.167, egg production vs physical event); Guangshan & Honglin, 1984 (p.118, tab.); De Decker, 1984 (p.316, 319: Rem., 341: chart); 1984 a (p.155); Binet, 1984 (tab.3); 1985 (p.85, tab.3); Brenning, 1985 a (p.16, 23, Table 2); Paffenhöfer, 1985 (p.104, distribution); Brinton & al., 1986 (p.228, Table 1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); M. Lefèvre, 1986 (p.33, 92); Sarkar & al., 1986 (p.178); Madhupratap & Haridas, 1986 (p.105, tab.1); Valentin & al., 1986 (p.117, Table V); Diouf & Diallo, 1987 (p.260); Jimenez-Perez & Lara-Lara, 1988; Williams D. & al., 1988 (p.580); Daan & al., 1988 (p.45, Table 4, carnivorous behaviour); Dessier, 1988 (tabl.1); Wiebe & al., 1988 (tab.7); Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Madhupratap & Haridas, 1990 (p.305, fig.4: vertical distribution night/day; fig.7: cluster); Othman & al., 1990 (p.561, 563, Table 1); Lindo, 1991 (tab.3); Hattori, 1991 (tab.1, Appendix); Yoo, 1991 (tab.1); Dai & al., 1991 (tab.1); Hernandez-Trujillo, 1991 (1993) (tab.I); Palomares Garcia & Vera, 1995 (tab.1); Shih & Young, 1995 (p.69); Kotani & al., 1996 (tab.2); Webber & al., 1996 (tab.1); Madhupratap & al., 1996 (p.866); Ramaiah & al., 1996 (p.3); Ramaiah & Nair, 1997 (tab.1); Park & Choi, 1997 (Appendix); Sharaf & Al-Ghais, 1997 (tab.1); Lakkis, 1998 (p.236, Rem.); Padmavati & al., 1998 (p.347); Achuthankutty & al., 1998 (p.1, Table 2: seasonal abundance vs monsoon); Hwang & al., 1998 (tab.II); Hsieh & Chiu, 1998 (tab.2); Wong & al, 1998 (tab.2); Mauchline, 1998 (tab.8, 16, 25, 40, 47, 48); Hopcroft & al., 1998 (tab.2); Suarez-Morales & Gasca, 1998 a (p109); Hernandez-Trujillo, 1999 (p.284, tab.1); El-Serehy, 1999 (p.172, Table 1, occurrence); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Dolganova & al., 1999 (p.13, tab.1); Razouls & al., 2000 (p.343, Appendix); Fernandez-Alamo & al., 2000 (p.1139, Appendix); Suarez-Morales & al., 2000 (p.751, tab.1); Dalal & Goswami, 2001 (p.22, fig.2); Lo & al., 2001 (1139, tab.I); El-Serehy & al., 2001 (p.116, Table 1: abundance vs transect in Suez Canal); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters, p.1353); Hernandez-Trujillo & Suarez-Morales, 2002 (p.748, tab.1); Dunbar & Webber, 2003 (tab.1); Osore & al., 2003 (p.69); Hwang & al., 2003 (p.193, tab.2); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.325, tab.1); Rezai & al., 2004 (p.489, tab.2, p.495, tab.8); Chang & Fang, 2004 (p.456, tab.1); Krumme & Liang, 2004 (p.407, tab.1); Lan & al., 2004 (p.332, tab.1); Lo & al.*, 2004 (p.218, fig.6); Lo & al., 2004 (p.89, tab.1); Wang & Zuo, 2004 (p.1, Table 2, dominance, origin); Gallienne & al., 2004 (p.5, tab.3); Satapoomin & al., 2004 (p.102, tab.2, 4, 6); Kazmi, 2004 (p.228, Rem.: p.233); Lakkis & al., 2005 (p.152); Alvarez-Silva & al., 2005 (p.39); Rezai & al., 2005 (p.157, Table 5: spatial & temporal variations); Obuid Allah & al., 2005 (p.123, occurrence % vs metal contamination); Waggett, 2005 (p.17, Table 3.3, behaviour); Hwang & al., 2006 (p.943, tab.I); Dias & Araujo, 2006 (p.36, Rem., chart); Lavaniegos & Jiménez-Pérez, 2006 (p.138, tab.2, 3, Rem.); Rakhesh & al., 2006 (p.93, Table 2, spatial distribution); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño, Rem.: p.70, 71, 72); Hwang & al., 2007 (p.24); Dur & al., 2007 (p.197, Table IV); Kovalev, 2006 (p.67: Lessepsian migration); Gaudy & Thibault-Botha, 2007 (p.84, Tab.2, Rem.: metabolism) ; Jitlang & al., 2008 (p.65, Table 1); McKinnon & al., 2008 (p.843: Tab.1); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Humphrey, 2008 (p.83: Appendix A); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.519); Fernandes, 2008 (p.465, Tabl.2); Selifonova & al., 2008 (p.305, Tabl. 2); Rakhesh & al., 2008 (p.154, abundance vs stations); Waggett & Buskey, 2008 (p.111, fig.3, Table 1); Muelbert & al., 2008 (p.1662, Table 1, 3); Lopez Ibarra, 2008 (p.1, Table 1, figs.11, 16: abundance, Table 3: N, C isotopes, figs.17, 18, 19, 22, 24, 25, 26, Table 4: index trophic); Hernandez-Trujillo & al., 2013 (p.61, egg production); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); Tseng L.-C. & al., 2008 (p.46, table 2, abundance vs moonsons); C.-Y. Lee & al., 2009 (p.151, Tab.2); Tseng & al., 2009 (p.327, fig.5, feeding); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Magalhaes & al., 2009 (p.187, Table 1, %); Cornils & al., 2010 (p.2076, Table 3); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Zenetos & al., 2010 (p.397); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Fazeli & al., 2010 (p.153, Table 1); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Shanthi & Ramanibai, 2011 (p.132, Table 1); Hsiao S.H. & al., 2011 (p.475, Appendix I); Maiphae & Sa-ardrit, 2011 (p.641, Table 2, 3, Rem.); Saitoh & al., 2011 (p.85, Table 5); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Hsiao & al., 2011 (p.317, Table 2, indicator of seasonal change); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Magris & al., 2011 (p.260, abundance, interannual variability); Beltrao & al., 2011 (p.47, Table 1, density vs time); El-Sherbiny & al., 2011 (p.837, seasonal abundance), Johan & al., 2012 (2013) (p.1, Table 1); Uysal & Shmeleva, 2012 (p.909, Table I, Rem.: p.927); Tseng & al., 2012 (p.621, Table 3: abundance, indicator species); Jose & al., 2012 (p.20, fig.3 a,b,c: % vs monsoon); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Lavaniegos & al., 2012 (p. 11, Table 1, fig.3, seasonal abundance, Appendix); Naz & al., 2012 (p.61, Table 4, relative abundance); Siokou & al., 2013 (p.238, Rem.: p.246); Gubanova & al., 2013 (in press, p.4, Table 2); Palomares-Garcia & al., 2013 (p.1009, Table I, fig.7, abundance vs environmental factors); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tachibana & al., 2013 (p.545, Table 1, seasonal change 2006-2008); Tseng & al., 2013 (p.507, seasonal abundance); Jagadeesan & al., 2013 (p.27, Table 3, 4, 5, 6, fig.11, seasonal abundance); Rakhesh & al., 2013 (p.7, Table 1, abundance vs stations); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); Alvarez-Silva & Torres-Alvarado, 2013 (p.241, Table1: seasonal abundance); Mendoza Portillo, 2013 (p.37: Fig.7, seasonal dominance); Lidvanov & al., 2013 (p.290, Table 2, % composition); Barton & al., 2013 (p.522, Table 1: metabolism, diapause, feeding mode, biogeo); El-Serehy & al., 2013 (p.2099, Rem.: p.2101); Palomares-Garcia & al., 2013 (p.187, vertical distribution, egg production); Terbiyik Kurt & Polat, 2013 (p.1163, Table 2, fig.9, seasonal distribution); Oh H.-J. & al., 2013 (p.192, Table 1, occurrence); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Lopez-Ibarra & al., 2014 (p.453, fig.6, biogeographical affinity); Nakajima & al., 2015 (p.19, Table 3: abundance); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production, Table 4: % vs. season); Rojas-Herrera & al., 2016 (p.40, Table 2: temporal abundance) ; Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Hernandez-Trujillo & Esqueda Escarcega, 2016 (p.1, egg production); Trottet & al., 2017 (p. 7, Table 3: resting stage); Jerez-Guerrero & al., 2017 (p.1046, Table 1: temporal occurrence); Palomares-Garcia & al., 2018 (p.178, fig.3: relative frequency, Table 1); Acha & al., 2020 (p.p.1, Table 3: occurrence % vs ecoregions). | | | | NZ: | 17 + 1 doubtful | | |
|
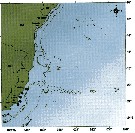
Distribution map of Centropages furcatus by geographical zones
|
| | | | | | | | | | | | | | | 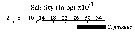 issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.8, Fig.6]. issued from : C.T. Achuthankutty, N. Ramaiah & G. Padmavati in Pelagic biogeography ICoPB II. Proc. 2nd Intern. Conf. Final report of SCOR/IOC working group 93, 9-14 July 1995. Workshop Report No. 142, Unesco, 1998. [p.8, Fig.6].
Salinity ranges for C. furcatus in coastal and estuarine waters of Goa (India).
Shaded area indicates the range of higher abundance. |
 issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.36]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.36].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S) |
 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 34. Jahrgang 1985. Mat.-nat. wiss. Reihe, 6. [p.5, Fig.1]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 34. Jahrgang 1985. Mat.-nat. wiss. Reihe, 6. [p.5, Fig.1].
T-S Diagram for Centropages furcatus from 8° S - 26° N; 16°- 20° W.
SO: Southern Surface Water (S °/oo: 34,50; T°C: 29,0); ND: Northern Water of the Surface Layer (S °/oo: 37,5; T°C: 21,0); SD: Southern Deep Water of the surface layer (S °/oo: 35,33; T°C: 13,4). See commentary in Temora stylifera and Brenning (1985 a, p.6). |
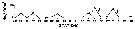 Issued from : M. Madhupratap, S.R. Sreekumaran Nair, C.T. Achuthankutty & V.R. Nair in Indian J. Mar. Sci., 1981, 10. [p.267, Fig.1f, g]. Issued from : M. Madhupratap, S.R. Sreekumaran Nair, C.T. Achuthankutty & V.R. Nair in Indian J. Mar. Sci., 1981, 10. [p.267, Fig.1f, g].
Distribution of copepod species in the Andaman Sea (around Andaman-Nicobar Islands). f: Centropages furcatus; g: Centropages calaninus.
Samples collected in vertical hauls (200-0 m) using a net (mesh aperture 500 µm). Cf. Madhupratap & al., 1981: Indian J. Mar. Sci., 10, p.258.
No. = number of individuals. |
 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 29- Centropages furcatus.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : S. Hernandez-Trujillo & G.Ma. Esqueda Escarcega in CICIMAR Oceanides, 2016, 31 (1). [p.3, Table 1 & 2]. Issued from : S. Hernandez-Trujillo & G.Ma. Esqueda Escarcega in CICIMAR Oceanides, 2016, 31 (1). [p.3, Table 1 & 2].
Table 1: Egg production rate (TPH: eggs female-1 day -1 mean during Marca Roja-VII cruise in Abril 2015. n = number of experiments with 5 replicates. Superficial water temperature, salinity and chlorophyll a at 10 m depth. Min: minimum number of eggs; Max: number maximum of eggs.
Table 2: Mean length of prosome (mm), quantity of carbone (µgC), K: enviromental and physiological factor according to Froese (2006), Rennie & Verdon (2008), Blackwell & al. (2000), Trudel & al;, (2005), Hernandez-Trujillo & al. (2008, 2013). Calculation of the weight (µg C) obtained by the relation mass-length (Lavaniegos & Ohman, 2007; Hernadez-Trujillo & al?, 2013): M = aLPb, where M = mass (µg), LP prosome length (mm), a intercept and b allometric coefficient (Durbin & al., 1978). The individuals variations in the relation between the mass and the length assumes an idea body, where the b valor is equal to 3, the 'condition factor' K = a. (M/L3, wghere M mass in µg C, L = prosome length (mm) and a = 0.1 (Hernadez-Trujillo & al;, 2013). |
 Issued from : S. Hernandez-Trujillo & G.Ma. Esqueda Escarcega in CICIMAR Oceanides, 2016, 31 (1). [p.4, Table 3]. Issued from : S. Hernandez-Trujillo & G.Ma. Esqueda Escarcega in CICIMAR Oceanides, 2016, 31 (1). [p.4, Table 3].
Egg production rate (TPH: eggs female-1 day -1). TSM: temperature (°C) of egg incubation. After authors cited. |
 Issued from : J.R. Palomares-Garcia, S. Hernandez-Trujillo, G.M. Esqueda-Escarcega & A. pérez-Morales, 2018 (p.L178, Fug.3]. Relationships frequency vs abundance of pelagic copepods in the La Paz Bay and the 22 main species (particularly the 3 dominant) over 96 species in the zone (about 21% of the total from the tropical Pacific species.); 59 species (13%) in the La Paz Bay show an annual cycle. Issued from : J.R. Palomares-Garcia, S. Hernandez-Trujillo, G.M. Esqueda-Escarcega & A. pérez-Morales, 2018 (p.L178, Fug.3]. Relationships frequency vs abundance of pelagic copepods in the La Paz Bay and the 22 main species (particularly the 3 dominant) over 96 species in the zone (about 21% of the total from the tropical Pacific species.); 59 species (13%) in the La Paz Bay show an annual cycle. |
 Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.37, Table 3]. Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.37, Table 3].
Summary of abundance and steady isotopics 15N and 13C in Subeucalanus subcrassus.
O = main feeding: omnivore; T = tropical biogeographical affinity; zones (a-f) = Cf. fig.1 in the same author.
Compare with Subeucalanus subcrassus and other species (Tabla 3).
Considered sometimes as carnivore. |
| | | | Loc: | | | Cosmopolite (equatorial-subtropical): Antarct. (very rare) (in T. Scott, 1913), sub-Antarct. (SW Pacif.), South Africa (E & W), Angola, Baia Farta, Congo, G. of Guinea, off Lagos, Ivorian shelf, Casamance, Dakar, Morocco-Mauritania, off Madeira, Argentina, Uruguay (continental shelf), Brazil (Campos Basin, Vitoria-Cabo de Sao Tomé, Mucuri estuary, Curuça estuary, off Amazon), Barbada Is., Porto Rico, Caribbean Colombia, Belize, Caribbean Sea, Jamaica, G. of Mexico, off Timbalier Island (Louisiana), Florida, Delaware Bay, Long Island Sound, Medit. ( W Basin: very rare, Turkey: Iskenderun Bay, Marmara Sea, W Black Sea), Canal of Suez ( Ras-el-Ech, Bitter Lakes, Lake Timsah), Suez Bay, Red Sea, Gulf of Oman, Arabian Gulf, UAE coast, G. of Mannar, Palk Bay, Natal, Mascarene Basin, Madagascar (Nosy Bé), Rodrigues Is. - Seychelles, Arabian Sea, Indian, Karachi coast, India (Saurashtra coast, Mangalore coast, Bombay, Goa, Cochin, Lawson's Bay, Madras, Burhabalanga estuary, Hooghly estuary Godavari region, Kakinada Bay), E India (coastal), Bay of Bengal, Andaman & Nicobar Is., Barren Island, off Phuket, Straits of Malacca, W Malay Peninsula (Andaman Sea), Cilacap Bay, off Labuan, Ambon Bay, Indonesian Seas, Australia (W, North West Cape), Thailand, Indonesia-Malaysia, Bintulu coast, SW Celebes, Philippines, Gulf of Tonkin, Hong Kong, China Seas (Yellow Sea, East China Sea, South China Sea, Xiamen Harbour), Taiwan Strait, Taiwan (S, E, SW, Kaohsiung Harbor, NW, N, Mienhua Canyon, NE), S Korea, Geumo Is., Korea Strait, Japan Sea, Japan (Tokyo Bay, Tanabe Bay), off Sanriku, W Pacif. (equatorial), Australia (G. of Carpentaria, Great Barrier, Moreton Bay, Shark Bay), New Caledonia, Moorea Is., California, Baja California (Bahia Magdalena, W), G. of California, Coyuca lagoon (Guerrero, W Mexico), La Paz, Acapulco Bay (rare), Guaymas Basin, G. of Tehuantepec, W Mexico, Zihuatanejo Bay, W Costa-Rica, Bahia Cupica (Cilombiaz), off Ecuador, Peru, off Chile (N & S) | | | | N: | 271 (Antarct.: 1; S Atlant.: 28; N Atlant.: 35; Medit.: 9; Suez Canal: 3; Red Sea: 5; Indian: 72; Indonesia-Malaysia: 16; Pacif.: 98) | | | | Lg.: | | | (16) F: 1,8-1,39; M: 1,71-1,49; (26) F: 1,82; M: 1,72-1,66; (34) F: 1,83-1,68; M: 1,6; (47) F: 1,7-1,6; M: 1,65-1,55; (78) F: 1,6; M: 1,55; (91) F: 1,7-1,6; M: 1,7-1,55; (104) F: 1,9; M: 1,7; (150) F: 1,85; (290) F: 1,75-1,6; M: 1,7-1,45; (322) F: 1,8; M: 1,5; (332) F: 1,82-1,76; M: 1,61-1,58; (333) F: 1,78-1,66; M: 1,61-1,59; (354) F: 1,73; (530) F: 1,9; M: 1,8; (776) F: 1,7-1,6; M: 1,7-1,55; (785) F: 1,84; 1,7; M: 1,55; (786) F: 1,86-1,61; M: 1,69-1,61; (795) F: 1,9; M: 1,4; (866) F: 1,38-1,82; (937) F: 1,68-1,78; M: 1,54-1,66; (991) F: 1,6-1,78; M: 1,55-1,65; (1047) F: 1,5-1,7; (1308) F: 1,65-1,92; M: 1,58-1,92; (1122) F: 1,65; M: 1,6; 0) F: (1230) F: 1,6-1,3; M: 1,5-1,7; {F: 1,38-1,92; M: 1,40-1,92}
Prosome length: (1254) F: 1.13 ± 0.12. | | | | Rem.: | epipelagic.
Sampling depth (Antarct., sub-Antarct.) : 0-1200 m.
For Fleminger & Hulsemann (1973, p.444-445) two forms exist: Atlantic (C. velificatus) and Indian-Pacific (C. furcatus s.s).
The form observed in the eastern Mediterranean would have a lessepsian origin, the form observed in the western part needs confirmation. After Siokou (2013, p.246) the occurrence of the species in the Northeast Aegean Sea suggests its spreading by the Levantine Intermediate Water pathway from the NW Levantine Sea through the easter straits of the Cretan Arc and along the east Aegean Sea. During the sampling period the Levantine Iitermediate Water (T = 16.2°C; S = 38.8) covered the entire water column.
Centropages furcatus carli Früchtl,1923 (M)
Syn.: C. furcatus : Carl, 1907 (p.8, figs.M)
Ref.: Früchtl, 1923 (p.452); 1924 b (p.45, figs.M); Sewell, 1932 (p.229, Rem.M); Vervoort, 1964 b (p.308)
Loc.: Indonesia-Malaysia (Ambon Bay).
After Itoh (1970 a, fig.2, from co-ordinates) the Itoh's index value of the mandibular gnathobase = 540.
After Mulyadi (2004, p.129) a variety of the male was described by Carl (1907) from Ambon Bay (SW Ceram), and has subsequently been recorded from the Aru Islands by Früchtl (1923) and the Indian Ocean by Sewell (1932). This species is the most widely distributed species of the genus and often occurring in large numbers.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is omnivorous.
After Barton & al. (2013, Table 1) this species is a mix of 'ambush' feeding mode and 'cruising' feeding mode (see definition in Centropages typicus).
Note the frequency between this species and Temora discaudata and, probably, their competition as did Centropages typicus and Temora stylifera in the bay of Banyuls (NW Mediterranean Sea, France).
See in DVP Conway & al., 2003 (version 1)
| | | Last update : 19/06/2023 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed February 20, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
