|
|
 |
|
Calanoida ( Order ) |
|
|
|
Eucalanoidea ( Superfamily ) |
|
|
|
Eucalanidae ( Family ) |
|
|
|
Eucalanus ( Genus ) |
|
|
| |
Eucalanus elongatus (Dana, 1849) (F,M) | |
| | | | | | | Syn.: | Calanus elongatus Dana,1849; no Sars, 1925 (p.20);
? Calanus erythrochilus Leuckart,1859;
No Eucalanus spinifer T. Scott, 1894 b (p.29, figs.F);
? Eucalanus atlanticus Wolfenden, 1904 (p.113); van Breemen, 1908 a (p.15); Candeias, 1926 (p.27); Rose, 1933 a (p.69, fig. juv.M); Unterüberbacher, 1964 (p.15, Rem.); Sameoto & al., 2002 (p.12);
Non Eucalanus giesbrechti Mori, 1937; Ohtsuka & al., 2015 (p.123, Rem.: p.125);
no Eucalanus elongatus : Esterly, 1905 (p.131); Campbell, 1929 (p.308);
? Eucalanus elongatus hyalinus : Morris, 1970 (p.2300);
No Eucalanus elongatus Park, 1968 (p.538, figs.F,M); No synonym of Eucalanus giesbrechti in Ohtsuka & al., 2015 (p.123, Table 2);
? Eucalanus elongatus : Isari & al., 2011 (p.51, Table 2, abundance vs distribution, in Thracian Sea); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1).
? Eucalanus elongatus elongatus : Acha & al., 2020 (p.p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions). | | | | Ref.: | | | Giesbrecht, 1892 (part., p.131,150, figs.F,M); Giesbrecht & Schmeil, 1898 (part., p.20); Cleve, 1904 a (p.189); Sewell, 1929 (p.48); Rose, 1933 a (p.67, figs.F,M, non localisations); Vervoort, 1946 (p.84, Rev., F,M, juv.); no atlantic forms: [Farran, 1908 b (p.21); With, 1915 (p.48, figs.F,M); Pesta, 1920 (p.498); Lysholm & Nordgaard, 1921 (p.8); [Rose, 1925 (p.151)]; Farran, 1926 (p.230); 1929 (part. p.207, 218); Rose, 1929 (p.14)]; [Rose, 1933, p.67, figs.F,M)]; ? Sewell, 1929 (p.48, figs.M: anormality); ? Sciacchitano, 1930 (p.8, Rem.); Dakin & Colefax, 1933 (p.204); [Jespersen, 1934 (p.46)]; Tanaka, 1935 (p.143, figs.F); [Hardy & Gunther, 1935 (1936) (p.139)]; Farran, 1936 a (p.78); Gibbons, 1936 (p.385, figs. Nauplii); Mori, 1937 (1964) (p.21, figs.F,M); Dakin & Colefax, 1940 (p.95, figs.M); Wilson, 1942 a (part., p.184); [Massuti Alzamora, 1942 (p.86)]; [Lysholm & al., 1945 (p.8)]; Sewell, 1947 (p.44, Rem.); 1948 (p.555, Rem.); Davis, 1949 (p.16, figs.F, Rem.); [Marques, 1947 (p.35)]; C.B. Wilson, 1950 (part., p.208); [Marques, 1953 (p.92); 1955 (p.33)]; Krishnaswamy, 1953 (p.111); Rose, 1956 (p.459); Tanaka, 1956 (p.267); Vervoort, 1957 (p.32, Rem.); ? Chiba & al., 1957 (p.306); ? 1957 a (p.11); Marques, 1959 (p.206); ? Fagetti, 1962 (p.10); Brodsky, 1962 c (p.105, figs.F); Ganapati & Shanthakumari, 1962 (p.7, 15); [Gaudy, 1962 (p.93, 99, Rem.: p.101)]; V.N. Greze, 1963 a (tabl.2)]; [Vervoort, 1963 b (p.88-89, Rem.]; [Shmeleva, 1965 b (p.1350, lengths-volume -weight relation)]; [Shmeleva, 1963 (p.141)]; [Duran, 1963 (p.12)]; [Paiva, 1963 (p.17, Rem.)]; [Giron-Reguer, 1963 (p.25)]; [Gaudy, 1963 (p.20, Rem.)]; [Unterüberbacher, 1964 (p.15, Rem.)]; De Decker, 1964 (p.15, 22, 28); De Decker & Mombeck, 1964 (p.12); [Ehrhardt, 1967 (p.737, geographic distribution), Rem.)]; ? De Decker, 1968 (p.45); Chen & Zhang, 1965 (p.34, figs.F); [Grice & Hulsemann, 1965 (p.223)]; [Mazza, 1966 (p.69)]; [Pavlova, 1966 (p.43)]; [Owre & Foyo, 1967 (p.36, figs.F,M)]; [Mazza, 1967 (p.65, 367, 384, figs.F,M, juv.)]; [Evans, 1968 (p.13)]; [Berdugo & Kimor, 1968 (p.448)]; [Ramirez, 1969 (p.45, figs.F, Rem.)]; Grice & Hulsemann, 1968 (tab.2); Koga, 1968 (p.16, fig.: egg); Vinogradov, 1968 (1970) (p.277); [Séguin, 1968 (p.488)]; [Corral Estrada, 1970 (p.72, figs.F,M, juv., Rem.)]; [Corral Estrada & Pereiro Muños, 1974 (tab.I)]; [Gamulin, 1971 (p.382, tab.3)]; Shih & al., 1971 (p.39, 147); [Deevey, 1971 (p.224)]; Bradford, 1972 (p.32, figs.F,M); [Binet & al., 1972 (p.71)]; [Roe, 1972 (p.277, tabl.1, tabl.2)]; [Nival & al., 1972 (p.63, respiration)]; [Razouls, 1972 (p.94, Annexe: p.11, figs.F)]; [Boucher & Thiriot, 1972 (p.47, Tableau 4)]; [Champalbert & al., 1973 (p.529, CHN composition)]; Fleminger, 1973 (p.967, 969, 978, 979, 999); Fleminger & Hulsemann, 1973 (p.343); [Harding, 1974 (p.141, tab.2)]; [de Bovée, 1974 (p.109, 124)]; Fleminger, 1975 (p.395); [Gaudy, 1975 (p.109, fig.1, Table I, 4, respiration)]; [Vives & al., 1975 (p.35,tab.II, III, IV XII)]; [Vives, 1976 (p.104)]; Tranter, 1977 (p.596); Goswami & Goswami, 1979 (p.103, figs.); [Dessier, 1979 (p.204)]; [Vaissière Séguin, 1980 (p.23, tab.2)]; Schnack, 1982 (p.145, fig.Md); Sazhina, 1982 (p.1156, Rem., fig.N); Zheng & al., 1982 (p.15, figs.F,M); [Kovalev & Shmeleva, 1982 (p.82)]; Brodsky & al., 1983 (p.204, figs.F, Rem.F,M); Van der Spoel & Heyman, 1983 (p.46, 147, fig.58); Tremblay & Anderson, 1984 (p.5: Rem.); [Scotto di Carlo & Ianora, 1983 (p.150)]; [Scotto di Carlo & al., 1984 (1042)]; [Sameoto, 1984 (p.767, Table I]); [Sazhina, 1985 (p.33, figs.N)]; [Price & Paffenhöfer, 985 (p.115, fig.2, Table 2, feeding)]; Longhurst, 1985 (tab.2); [Regner, 1985 (p.23)]; Madhupratap & Haridas, 1986 (p.105, tab.1); Sarkar & al., 1986 (p.178); [Lozano Soldevilla & al., 1988 (p.57)]; [Schnack, 1989 (p.137, tab.1, fig.6: Md)]; [Pancucci-Papadopoulou & al., 1990 (p.199)]; [Timonin, 1990 (p.479)]; Mitra & al., 1990 (fig.3); Hirakawa & al., 1990 (tab.3); Flint & al., 1991 (p.199); Hattori, 1991 (tab.1, Appendix); [Suarez & Gasca, 1991 (tab.2)]; [Scotto di Carlo & al., 1991 (p.270)]; [Suarez, 1992 (App.1); ]; [Seguin & al., 1993 (p.23)]; [Hays & al., 1994 (tab.1)]; [Kouwenberg, 1994 (tab.1)]; [Webber & Roff, 1995 (tab.2)]; Shih & Young, 1995 (p.69); Kotani & al., 1996 (tab.2); [Siokou-Frangou, 1997 (tab.1)]; Go & al., 1997 (tab.1); Park & Choi, 1997 (Appendix); [Suarez-Morales, 1998 (p.345, Table 1); ]; [Lopez-Salgado & Suarez-Morales, 1998, (p.318)]; Suarez-Morales & Gasca, 1998 a (p.109); Mauchline, 1998 (tab.8); [Lopez-Salgado & al., 2000 (tab.1)]; [Sautour & al., 2000 (p.531, Table II, abundance)]; Madhupratap & al., 2001 (figs.4, 5); [d'Elbée, 2001 (tabl.1)]; [Holmes, 2001 (p.40)]; Lo & al., 2001 (1139, tab.I); [Weikert & al., 2001 (p.229, fig.7,8,9, Rem.)]; [Zerouali & Melhaoui, 2002 (p.91, Tableau I)]; [Beaugrand & al., 2002 (p.179, figs.5, 6)]; [Sameoto & al., 2002 (p.12)]; [Vukanic, 2003 (139, tab.1)]; Conway & al., 2003 (p.163, figs.F,M, Rem.); Goetze, 2003 (p.2322 & suiv.); Hsiao & al., 2004 (p.325, tab.1); [Daly Yahia & al., 2004 (p.366, fig.1)]; [Koppelmann & Weikert, 2007 (p.266: tab.3)]; Boxshall & Halsey, 2004 (p.117: figs.F,M); Lo & al., 2004 (p.89, tab.1); [G. Harding, 2004 (p.40, figs.F,M)]; [Isari & al., 2006 (p.241, tab.II)]; Fernandes, 2008 (p.465, Tabl.2); [Cabal & al., 2008 (289, Table 1)]; [Moscatello & al., 2011 (p.80, Table 4)]; [Brugnano & al., 2012 (p.207, Table 2, 3)] | 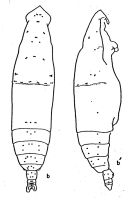 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.989, Fig.13]. Female: b, habitus (dorsal), b', idem (lateral right side).
Dorsal and lateral pattern of integumental organs (black point = sites occuring at 100% frequency, o = 80-99% frequency, x = 10-79% frequency, triangle are sites which are also visible in lateral view but which are assigned to dorsal sets);
|
 issued from : A. Fleminger in Fishery Bull. natn. Ocean. Atmos. Adm., 1973, 71 (4). [p.968, Fig. l (p.969)]. Female: l, genital segment (lateral right side).
|
 issued from : K.A. Brodsky, N.V. Vyshkvartzeva, M.S. Kos & E.L. Markhaseva in Opred Fauna SSSR, 1983, 135. [p.205, Fig.92]. Female.: habitus (after Fleminger, 1973, redrawn); genital somite; P1.
|
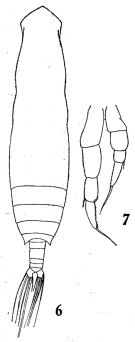 issued from : Q.-c. Chen & S.-z. Zhang & C.-s. Zhu in Studia Marina Sinica, 1974, 9. [Pl.1, Figs.6-7]. Male (from China Seas): 6, habitus (dorsal); 7, P5.
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.116, Fig.6-1]. Female: a, habitus (dorsal); b, forehead (lateral); c-d, urosome (lateral and ventral, respectively); e, A2; f, Md; g, Mx1; h, P1; i, basal portion of P1. Scale bar in mm.
|
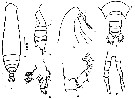 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.117, Fig.6-2]. Male: j, habitus (dorsal); k, forehead (lateral); l-m, urosome (dorsal and lateral, respectively); n, Mx1; o, basal portion of P1; p, P5. Scale bar in mm.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.7, Figs.1-5]. Female: 1, habitus (dorsal); 3, Md; 5, Mx1. Nota: Head triangular. Lateral angles of the last thoracic segment pointed. 2 segments between the anal and genital segments. 1st and 2nd segments of exopodite of A2 not fused; 1st segment of endopodite slightly longer than the 2nd, and over 3 times as long as its broad. Inner margin of the 2nd segment of basipodite of Md with 3 setae, divided into 2 nearly equal portions by the insertion of the endopodite; endopodite with 2 setae on the 1st segment, 5 setae on the 2nd. Male: 2, habitus (dorsal); 4, P5.
|
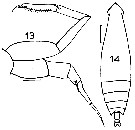 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.33, Fig.4 (13-14)]. Female (from Kaikoura, New Zealand): 14, habitus (dorsal). Male: 13, P5. Scale bars: 1 mm (14); 0.1 mm (13).
|
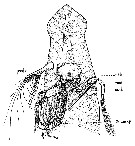 Issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1916, 16 (14). [p.173, Fig.1]; head (from below view) of Eucalanus. The appendages and bristles are shown diagrammatic. On the left ot the figure are shown, from behind forward, Mx2 (ant. mxp), the outer lobe of Mx1, A2 (post. ant.); on the right appear the inner lobe of Mx1 (mxl) with stiff, heavy bristles and the rami of Md (mnd.), the ''jaw'' being omitted. The shield-shaped prominence in the center of the figure bears the scoop-like lip, within which is the opening of the mouth (mth.). The side of the funnel formed by the bristles of Mx2 is shown on the left of the figure, the other side being omitted for the sake of clearness.
| | | | | Compl. Ref.: | | | [Pearson, 1906 (p.4, Rem.)]; ? Esterly, 1916 (p.171, nutrition); 1919 (p.1, 41, light-dark reactions); [Jespersen, 1939 (p.38); Østvedt, 1955 Table 3)]; Yamazi, 1958 (p.147, Rem.); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio/Kuroshio transitional area, Table 7, 10); Timonin, 1971 (p.281, trophic group); Gueredrat, 1971 (p.300, Table 1, 2); Subbaraju & Krishnamurphy, 1972 (p.25, 26 ); [Bainbridge, 1972 (p.61, Appendix Table I: vertical distribution vs day/night)]; [P. Nival & S. Nival, 1973 (p.135, mouth parts, grazing)]; [Harding, 1974 (p.141, Table 2, gut contents); Patel, 1975 (p.659); [Pipe & Coombs, 1980 (p.223, Table II, vertical occurrence)]; Madhupratap & al., 1981 (p.266, fig.1i: abundance vs. geographic transect); Kukina, 1983 (p.355, feeding); Guangshan & Honglin, 1984 (p.118, tab.); Schulz, 1990 (p.188); Madhupratap & Haridas, 1990 (p.305, fig.6: vertical distribution night/day; fig.7: cluster); [Seguin & al., (1993 (p.23, Table 2: abundance, %)]; [Seguin & al., 1994 (p.369, Rem.: p.376: Rem.]; Achuthankutty & al., 1998 (p.1, Table 2, seasonal abundance vs monsoon); ? El-Sherif & Aboul Ezz, 2000 (p.61, Table 3: occurrence); Pakhomov & al., 2000 (p.1663, Table 2, transect Cape Town-SANAE antarctic base); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters); Prusova & Smith, 2005 (p.76); Zuo & al., 2006 (p.162: tab.1); [? Khelifi-Touhami & al., 2007 (p.327, Table 1)]; Hwang & al., 2007 (p.24); Dur & al., 2007 (p.197, Table IV); Morales-Ramirez & Suarez-Morales, 2008 (p.519); Wishner & al., 2008 (p.163, Table 2, fig.8, oxycline); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Rakhesh & al., 2008 (p.154, Table 5: abundance vs stations); [Licandro & Icardi, 2009 (p.17, Table 4)]; [Hafferssas & Seridji, 2010 (p.353, Table 2: as elongates)]; Goetze & Ohman, 2010 (p.2110, Table 1, biogeography); [Mazzocchi & Di Capua, 2010 (p.425)]; [ Medellin-Mora & Navas S., 2010 (p.265, Tab. 2)]; Shanthi & Ramanibai, 2011 (p.132, Table 1); [Andersen N.G. & al., 2011 (p.71, Fig.3: abundance)]; Hsiao S.H. & al., 2011 (p.475, Appendix I); Guo & al., 2011 (p.567, table 2, indicator); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Jose & al., 2012 (p.20, fig.3 a,b,c: % vs monsoon); [Bode & al., 2012 (p.108, spatial distribution vs time-series, % biomass)]; [? Aubry & al., 2012 (p.125, fig.8 a, b, interannual variation)]; [Belmonte & al., 2013 (p.222, Table 2, abundance vs stations)]; in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tseng & al., 2013 (p.507, seasonal abundance); Jagadeesan & al., 2013 (p.27, Table 3, seasonal variation); Anjusha & al., 2013 (p.40, Table 3, abundance & feeding behavior); [ Lidvanov & al., 2013 (p.290, Table 2, % composition) ]; Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Lopez-Ibarra & al., 2014 (p.453, Rem.: p. 460); Fierro Gonzalvez, 2014 (p.1); Belmonte, 2018 (p.273, Table I: Italian zones) (p.1, Tab. 3, 5, occurrence, abundance); Chiba S. & al., 2015 (p.968, Table 1: length vs climate); [ ? Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); ? El Arraj & al., 2017 (p.272, table 2, spatial distribution); ] ; [ ? Belmonte, 2018 (p.273, Table I: Italian zones); ? Chaouadi & Hafferssas, 2018 (p.913, Table II, III, fig.4: occurrence, abundance, seasonal variation)]. | | | | NZ: | 10 + 2 doubtful | | |
|
Distribution map of Eucalanus elongatus by geographical zones
|
| | | | | | | | | | | | 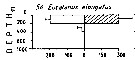 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 56- Eucalanus elongatus.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
| | | | Loc: | | | South Africa (E & ?W), SE Atlant. (in Pakhamov & al., 2000), ? Red Sea (Sharm El-Sheikh), G. of Aden, G. of Oman, Arabian Sea, Laccadive Is., Maldive Is., Madagascar, Indian, India (Saurashtra coast, Porto Novo, Mangalore coast, Goa, Madras, Gulf of Mannar, Palk Bay, Lawson's Bay, Madras, Mandarmani creek, Hooghly estuary, Kakinada Bay), Bay of Bengal, Nicobar Is., Barren Island, W Australia, Indonesia-Malaysia, Viet-Nam (Cauda Bay), China Seas (Yellow Sea, East China Sea, Taiwan Strait, South China Sea), Taiwan, off SW & NE Taiwan, Mienhua Canyon, Kuroshio Current, Okinawa, Korea, Japan, off Hokkaido, Kuril Is., Pacif. (central N), California, W Baja California, off W Guatemala, W Costa Rica, Pacif. (W equatorial), Australia (Great Barrier), New Zealand (Kaikoura), off Juan Fernandez Is., Galapagos-Ecuador, Peru, Chile | | | | N: | 87 ?? (some forms being confused) | | | | Lg.: | | | (28) F: 5,73-4,53; M: 4,46-3,60; (91) F: ± 4,5; M: ± 3,2; (104) M: 4,5; (116) F: 7,1; M: 4,2; (125) F: 5,76-5,62 ; (131) F: 4,53-5,73; M: 3,6-4,46; (150) F: 7-6,5; (290) F: 5,2-5,55; (338) M: 3,5-4,1; (795) F: 5,5; M: 3,8; (991) F: 5,75-8,5; M: 3,75-5; (1023) F: 5,22; M: 4,19; {F: 4,55-8,50; M: 3,20-5,00}
Chiba S. & al., 2015 (p.971, Table 1: Total length female (June-July) = 6.5 mm [optimal SST (°C) = 3.3]. | | | | Rem.: | Depth: 298-526 m at Station N-1 (SW Bösö, E middle Japan) from Furuhashi (1966 a).
Vervoort, 1963 b, p.88-89) rejects Johnson's (1938) subdivision of the Eucalanus elongatus complex into a number of separate species and subspecies, basing the conclusion largely on material from the seas of the East Indies. Part of that material possessed characters intermediate between those used by Johnson to discriminate between his various species and subspecies. This made strongly doubt the value of the taxons institued by Johnson. Though he believes that the various characteristics used by Johnson to established the taxons in his 1938 paper are in need of a critical revision, the occurrence of two separate species, Eucalanus bungii Giesbrecht, 1892 and Eucalanus inermis Giesbrecht, 1892, besides Eucalanus elongatus (Dana, 1849), seems to be beyond doubt. Nomenclatural difficulties might arise from the fact that Dana's figure and description of Eucalanus elongatus are inadequate to decide to which of the various taxons of the E. elongatus complex it really refers.
The geographical distribution is limited here to the Indo-Pacific form, sensu stricto.
The varieties are established according to the rank of the species.
The forms reported by the authors under this denomination, between brackets, should presumably correspond to Eucalanus hyalinus.
Khelifi-Touhami & al., 2007 (p.327, Table 1) note the two species E. elongatus and E. hyalinus from the algerian coast, the first needs confirmation.
After us, this species is noted for the first time in the Red Sea by El-Sherif & Aboul Ezz (2000, p.72) and need confirmation.
For Itoh (1970 a, fig.2, from co-ordonates) the Itoh's index value of the mandibular gnathobase = 200.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a coarse-filter feeder.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 28/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed March 23, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
