|
|
 |
|
Calanoida ( Order ) |
|
|
|
Clausocalanoidea ( Superfamily ) |
|
|
|
Euchaetidae ( Family ) |
|
|
|
Euchaeta ( Genus ) |
|
|
| |
Euchaeta media Giesbrecht, 1888 (F,M) | |
| | | | | | | Syn.: | Euchäta media Giesbrecht, 1892 (p.246, 263, 772, figs.F);
Euchaeta diegensis Esterly, 1911 (p.323, figs.F); Sewell, 1948 (p.556); Brodsky, 1950 (1967) (p.202, figs.F, Rem.); ? De Decker & Mombeck, 1964 (p.13); Hernandez-Trujillo, 1991 (1993) (tab.I);
? E. acuta pacifica Esterly, 1911 (p.324, fig.M); Brodsky, 1950 (1967) (p.199, figs.M); Roe, 1972 b (p.528: Rem.);
Paraeuchaeta media : Bradford & al., 1983 (p.43, figs.F, M, Rem.); Errhif & al., 1997 (p.422); Chihara & Murano, 1997 (p.801, Pl.107,110: F,M); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Hirai & al., 2013 (p.1, Table I, molecular marker);
Pareuchaeta media : Heinrich, 1990 (p.17) | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.39, Rem. F); Esterly, 1905 (p.160, figs.F); A. Scott, 1909 (p.66, figs.F); Bigelow, 1926 (p.230); Farran, 1929 (p.208, 238); Sewell, 1929 (p.133, 149); Candeias, 1926 (1929) (p.32, 49, figs.F); Rose, 1933 a (p.116, figs.F); Farran, 1936 a (p.89); Mori, 1937 (1964) (p.46, figs.F); Sewell, 1947 (p.116); Brodsky, 1950 (1967) (p.201, figs.F); C.B. Wilson, 1950 (p.214, figs.M, Rem.F,M); Vervoort, 1957 (p.77, 84, Rem.); Tanaka, 1958 (p.330, figs.F); Grice, 1962 (p.201, figs.F); Vervoort, 1963 b (p.161, Rem.); Paiva, 1963 (p.43); Gaudy, 1963 (p.23); Owre & Foyo, 1967 (p.54, figs.F); Park, 1968 (p.551, Redescr.M, figs.F,M); Tanaka & Omori, 1968 (p.221); Shih & al., 1971 (p.39); Vives, 1972 (p.212, figs.F); Tanaka, 1973 (p.135, fig.F); Park, 1975 c (p.5, figs.F,M); 1978 (p.207, figs.F,M); Dawson & Knatz, 1980 (p.6, figs.F,M); Björnberg & al., 1981 (p.633, 635, figs.F,M); Gardner & Szabo, 1982 (p.264, figs.F,M); Roe, 1984 (p.357); Park, 1995 (p.24, Rem.F,M, figs.F,M); Mauchline, 1999 (n°182, p.8, figs.F,M); Bradford-Grieve & al., 1999 (p.880, 925, figs.F,M); G. Harding, 2004 (p.53, figs.F,M); Mulyadi, 2004 (p.72, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.651, figs.F,M, Rem.); Blanco-Bercial & al., 2011 (p.103, Table 1, Biol. mol, phylogeny) |  issued from : T. Park in Antarctic Res. Ser. Washington, 1978, 27. [p.209, Fig.67]. Female: A, forehead (lateral); B, C,D,E, distal end of metasome and genital segment (left side, right side, dorsal and ventral); F, outer lobe of Mx1; G, P1 (anterior); H, P2 (anterior). Nota: Proportional lengths of prosome and urosome 65 : 35. Frontal eminence of forhead pronounced, bearing suprafrontal sensilla. Rostrum powerfully developed, pointing obliquely forward at an angle of about 75° with reference to body; its tip curved slightly downward. Posterilateral corner of metasome not prolonged posteriorly, its posterior margin broadly rounded either dorsally and laterally. Genital segment strongly asymmetrical in either dorsal or ventral view, with large protuberance anterior to middle of its left side and low ridge forming characteristic step posterior to middle of its right side. Ventrally, genital field asymmetrical, rotated clockwise anout 30° with reference to body axis. Laterally, genital prominence directed antero ventrally, with genital field facing obliquely forward. Genital flanges asymmetrical; right flange small, produced at middle into pointed process; left flange much larger and triangular with its top posterior to pointed process of right flange. Appendicular caudal, setae straight, much thicker and longer than terminal setae. A1 (when pressed against body) extending beyond distal end of metasome by its last 2 segments. Outer coxal lobe of Mx1 with 7 setae, 2 most proximal ones much shorter than others. In Mx2, 1 of 6 apical setae bearing long, widely separated spines in addition to short spinules found densely along its entire length. Long endopodal setae of Mxp terminating in spinule (as in E. marina). First 2 exopodal segments of P1 fused, leaving no trace of articulation; with single external spine reaching distal end of following segment. In P2, external spine of 2nd exopodal segment reaching base of 1st external spine of 3rd; 2nd external spine of 3rd exopodal segment arising from lobe separated from segment by deep incision, reaching close to base of 3rd external spine; 1st and 3rd external spines of 3rd exopodal segment of about equal size.
|
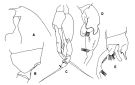 issued from : T. Park in Antarctic Res. Ser. Washington, 1978, 27. [p.210, Fig.68]. Male: A, forehead (lateral); B, last metasomal and genital segments (lateral); C, P5 (anterior); D, middle part of exopod of left P5 (anterior); E, middle part of exopod of left P5 (medial).
|
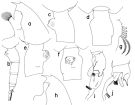 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.123, Fig.13]. Female: a, forehead (left side); b, urosome (left); c, d, e, f, genital somite (left, dorsal, right, ventral, respectively); g, outer lobe of Mx1. Male: h, forehead (left); i, j, exopod of left 5th leg (anterior, medial, respectively).
|
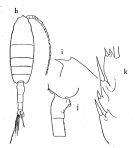 Issued from : O. Tanaka in Publs Seto mar. Biol. Lab., 1958 [p.329, Fig.62]. Female: h, habitus (dorsal); i, forehead (lateral); j, last thoracic segment and urosomal segments 1 and 2 (lateral right side); k, outer margin of exopod of P2.
|
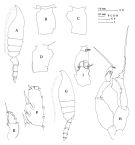 issued from : J.M. Bradford, L. Haakonssen & J.B. Jillett in Mem. N.Z. Oceanogr. Inst., 1983, 90. [p.45, Fig.23]. As Paraeuchaeta media. Female: A, habitus (lateral left side); B-D, genital segment (dorsal, left lateral, right lateral, respectively); E, exopod of P1; F, exopod segment 3 of P2. Male: G, habitus (lateral left side); H, P5; I, terminal part of left P5 exopod. Nota: - P1 exopod female: Bb = BC; Cc > BC. - P2 exopod female: Aa = AB; Bb = 1/3 BC; Cc = 5/8 CD; Dd = 1/4 CD.
|
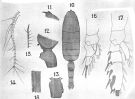 issued from : A. Scott in Siboga-Expedition, 1909, XIX a. [Plate XX, Figs.10-18]. Female (from Banda Sea): 10, habitus (dorsal); 11, forehead (lateral); 12, last thoracic and genital segments (left side); 13, genital segment (dorsal); 14, A1; 15, Mxp (end hair); 16, P1; 17, P2; 18, part of terminal spine of exopodite of P3.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.19, Figs.9-13]. Female: 9, habitus (dorsal; with eggs); 10, P2; 11, last thoracic and genital segmens (lateral); 12, forehead (lateral); 13, genital area.
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1911, 6 (14). [Pl.28, Fig.37]. Female (from San Diego Region): 37, forehead (lateral).
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1911, 6 (14). [Pl.29, Figs.49, 55]. Female: 49, part of last thoracic segment and genital segment (lateral, left side); 55, genital segment (ventral). Nota: For Esterly, the urosome and genital segment resembles Euchaeta media Giesbrecht, but the genital convexity and the protuberances about the orifice are different; the right side of the segment as seen in outline is more irregular, with a smaller hnob-like protrusion near the posterior margin of the segment.
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1911, 6 (14). [Pl.31, Fig.92]. Female: 92, part of exopod of P2. Nota: In exopod of P2, the middle spine of the outer margin reaches two-thirds of the distance from its base to the base of the third outer marginal spine (in Euchaeta media) it is about one-third as long).
|
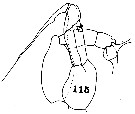 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1911, 6 (14). [Pl.32, Fig.115]. As Euchaeta acuta var. pacifica. With doubt. Male (from San Diego Region): 115, P5 (the left leg is at left). Nota: P5 resemble those of Euchaeta acuta (see in Giesbrecht, 1892, Pl.16, fig.18, 21 (not 19)), but the process on the 2nd segment of exopodite of the left leg is not pyramidal and pointed, and in addition the endopodite of the right leg is broadened at the distal end and carries a flap-like appendage.
|
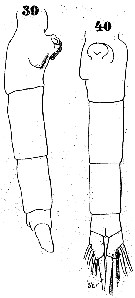 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.37, Figs.39, 40]. As Euchäta media. Female: 39-40, urosome (lateral and ventral, respectively).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf.16, Fig.13]. As Euchäta media. Female: 13, exopodal segments 2 and 3 of P2 (posterior view).
|
 issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1905, 2 (4). [p.161, Fig.25]; Female (from San Diego Region): a, habitus (lateral), with egg sac; b, forehead (lateral); c-d, genital segment (dorsal and right lateral side, respectively); e, P2; f, Mx1 (Le.1 = 1st lobe of outer margin; Li.1 = 1st lobe of inner margin; Ri.2 and 3 = fused 2nd and 3rd segments of endopod, bristles not shown); g, caudal rami (Si = inner marginal bristle). Nota: A1 extend a little beyond the posterior border of the genital segment.1st outer marginal lobe of Mx1 with 8 bristles, 2nd basal with 3, fused 2nd and 3rd segments of the endopodite with 4. Outer border of 1st segment of exopodite of P1 concave; outer border of 3rd segment of P2 and its outer bristles different than in the following pair.
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.200, Pl.13, Figs.10-14]. Female (from 01°10'S, 134°57'W): 10, genital segment (dorsal); 11, same (ventral); 12, same (left side); 13, same (right side); 14, P2. Characteristic swelling on the left side and the knob-like protrusion on the right side of the genital segment. The middle spine on the outer edge of the 3rd exopodal segment of P2 is elongate and the spine on the 2nd exopodal segment reaches beyond the base of the proximal spine on the 3rd segment.
|
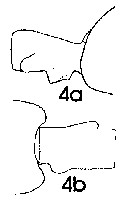 issued from : J. Mauchline in ICES Ident. Leafl. Plankton, 1999, N°182. [p.3, Fig.2: 4a-4b]. Female (Northeast Atlantic): 4a, genital double-somite (right side); 4b, same (dorsal).
|
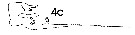 issued from : J. Mauchline in ICES Ident. Leafl. Plankton, 1999, N°182. [p.4, Fig.3: 4c]. Male (Northeast Atlantic): 4c, terminal two segments, exopodal segments of left P5.
|
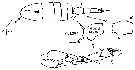 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.53]. Female & Male.
|
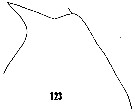 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. [p.26, Fig.123]. Female: 123, forehead (lateral).
|
 issued from : T. Park in Smiths. Contr. Zool., 1975, 196. [p.6, Fig.2]. Female (G. of Mexico): a, forehead (lateral); b, genital segment (ventral). Male: c, exopod of left P5 (anterior).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.16, Fig.36]. As Euchäta spinosa. Female: 36, outer margin of Mx2.
|
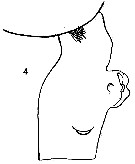 issued from : O. Tanaka in I O B C Handbook, 1973, IV. [p.148, Fig.9: 4]. Female (from Indian Ocean): 4, genital segment (lateral). Nota: Genital segment asymmetrical, shorter than the following three segments together. The right side with a low rounded process near the distal margin.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.73, Fig.41]. Female (from 07°29'S, 121°05'E): a, habitus (dorsal); b, posterior part of last thoracic segment and genital complex (dorsal); c, same (lateral left side); d, forehead (lateral); e, P1; f, P2; g, left caudal ramus (dorsal). Male: h, habitus (dorsal); i, forehead (lateral); j, P5; k, exopod of left P5.
|
 Issued from : T. Park in Smithsonian Contr. Zool., 1975, 196. [p.6, Fig.2]. Female (from G. of Mexico & Caribbean Sea): a, forehead (lateral); b, genital segment (ventral). Male: c, exopod of left P5 (anterior).
|
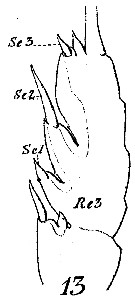
 Euchaeta media Euchaeta media Female: 1 - See Key to acuta species Group. 2 - Dorsally, genital somite asymmetrical. 3 - Dorsally, genital somite with steplike notch on right side (Fig.13-d).
| | | | | Compl. Ref.: | | | Cleve, 1904 a (p.190); Sewell, 1948 (p.391); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Grice, 1963 a (p.495); Gaudy, 1963 (p.23, Rem.); Unterüberbacher, 1964 (p.23); De Decker & Mombeck, 1964 (p.12); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio/Kuroshio transitional area, Table 7, 9, 10); Mazza, 1966 (p.70); 1967 (p.331); Fleminger, 1967 a (tabl.1); Morris, 1970 (p.2300); Park, 1970 (p.476); Gueredrat, 1971 (p.300, fig.2, 10, Table 1, 2); Deevey, 1971 (p.224); Timonin, 1971 (p.281, trophic group); Bainbridge, 1972 (p.61, Appendix Table III: occurrence); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 b (p.528); Harding, 1974 (p.141, tab.2, gut contents); Vives & al., 1975 (p.42, tab.II); Tranter, 1977 (p.596); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Carter, 1977 (1978) (p.35); Dessier, 1979 (p.205); Hayward, 1980 (p.295, Table 2, vertical distribution, feeding); Vives, 1982 (p.291); Dessier, 1983 (p.89, Tableau 1, Rem., %); Tremblay & Anderson, 1984 (p.5); De Decker, 1984 (p.316); Guangshan & Honglin, 1984 (p.118, tab.); Cummings, 1984 (p.163, Table 2); Longhurst, 1985 (tab.2); Brinton & al., 1986 (p.228, Table 1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Madhupratap & Haridas, 1986 (p.105, tab.1); Rudyakov, 1986 (tab.1); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Lozano Soldevilla & al., 1988 (p.58); Jimenez-Perez & Lara-Lara, 1988; Hernandez-Trujillo, 1989 a (tab.1); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Hernandez-Trujillo, 1991 (1993) (tab.I); Suarez, 1992 (App.1); Hattori, 1991 (tab.1, Appendix); Mauchline, 1992 a (p.2); Shih & Young, 1995 (p.70); Go & al., 1997 (tab.1); Alvarez-Cadena & al., 1998 (tab.1,2,3,4); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p.109); Mauchline, 1998 (tab.42); Lavaniegos & Gonzalez-Navarro, 1999 (p.239, Appx.1); Lapernat, 2000 (tabl.3, 4); Haury & al., 2000 (p.69, Table 1, Fig.7); Lopez-Salgado & al., 2000 (tab.1); Rebstock, 2001 (tab.2); Holmes, 2001 (p.52); Beaugrand & al., 2002 (p.179, figs.5, 6); Rebstock, 2002 (p.71, Table 3, 5, 6, Figs.2, 3: climatic variability); Hsiao & al., 2004 (p.326, tab.1); Pusch & al., 2004 (251, tab.3); Lo & al., 2004 (p.89, tab.1); Berasategui & al., 2005 (p.313, fig.2); Lavaniegos & Jiménez-Pérez, 2006 (p.143, tab.2, 3, Rem.); Mackas & al., 2006 (L22S07, Table 2); Lopez-Ibarra & Palomares-Garcia, 2006 (p.63, Tabl. 1, seasonal abundance vs El-Niño, Rem.: p.72); Dur & al., 2007 (p.197, Table IV); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.520); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Fernandes, 2008 (p.465, Tabl.2); C.-Y. Lee & al., 2009 (p.151, Tab.2); Galbraith, 2009 (pers. comm.); Williamson & McGowan, 2010 (p.273, Table 3, Pacific central gyres: N and S); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2): Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Palomares-Garcia & al., 2013 (p.1009, Table I, abundance vs environmental factors); Sano & al., 2013 (p.11, Table 2, fig.5, feeding habits); Lidvanov & al., 2013 (p.290, Table 2, % composition); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions) ; Zakaria & al., 2016 (p.1, Table 1, Rem.); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence). | | | | NZ: | 18 | | |
|
Distribution map of Euchaeta media by geographical zones
|
| | | | | | | | | | | | 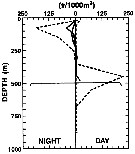 issued from : L. Haury, C. Fey, C. Newland & A. Genin in Progress in Oceanography, 2000, 45 [p.83, Fig.7]. issued from : L. Haury, C. Fey, C. Newland & A. Genin in Progress in Oceanography, 2000, 45 [p.83, Fig.7].
Vertical distribution of Euchaeta media female from day and night tows taken over (solid lines) and away from (dashed lines) the seamount Fieberling Guyot (32°25'N, 127°47'W).
The horizontal line at 500 m marks the approximate depth of the summit plain. Note the reduction in abundance at night over the seamount. |
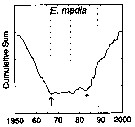 issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 a]. issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 a].
Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California (31°-35°N, 117°-122°W, from years 1950 to 2000.
Cumulative sums of nonseasonal anomalies from the long-term means of copepod abundance .
A negative slope indicates a period of below-average anomalies; a positive slope indicates a period of above-average anomalies. Abrupt changes in slope indicate step changes. Step changes are marked with arrows (downward -pointing for decreases).
The October 1966 cruise (prior to the increase in sampling depth), March 1976 cruise (prior to the 1976-77 climatic regime shift), and October 1988 cruise (prior to the hypothesized 1989 climatic regime shift) are marked with vertical lines. |
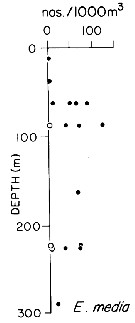 issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.299, Fig.2]. issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.299, Fig.2].
Plots of day (open circle) and night (filled circle) depth distributions of Euchaeta media in the North Pacific central gyre (main sampling location: 25°N, 155°W with drogue), September 1968. |
 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.10 g]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.10 g].
Verical distribution of Euchaeta media at an oligotrophe site (off NW Cape Verde Islands: 21° N, 31° W) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
| | | | Loc: | | | South Africa (E), Namibia, Congo, G. of Guinea, off Lagos, Cape Verde Is., off Mauritania-NW Cape Verde Is., Morocco-Mauritania, Great meteor Seamount, Canary Is., off Madeira, Argentina, Brazil, off Rio de Janeiro, off Amazon, Caribbean, Caribbean Sea, Caribbean Colombia, Yucatan, G. of Mexico, Florida, off Bermuda: Station "S" (32°10'N, 64°30'W), off E Cape Cod, G. of Maine (rare), off Rockall Is., Portugal, off W Cape Finisterre, Ibero-moroccan Bay, W Medit. (Algiers Bay, W Egyptian coast), Arabian Sea, Natal, Indian, S Indian (subtropical convergence), Nicobar Is. (Nankauri Harbour), Bay of Bengal, W Australia, Indonesia-Malaysia, Flores Sea, Philippines, China Seas (East China Sea, South China Sea), Taiwan, Kuroshio Current, Okinawa, Japan, Sagami Bay, off Sanriku, California, W Baja California, Bahia Magdalena, La Paz, Gulf of California, off W Guatemala, W Costa Rica, Pacif. (W equatorial), Pacific (central gyres: N and S), Australia (Great Barrier), New Zealand, Bikini Is., Hawaii, off NE Hawaii, Pacif. (central subtropical N), Costa Rica, Pacif. (SE tropical), Peru, off Chile, Chile (N-S) | | | | N: | 128 | | | | Lg.: | | | (3) F: 4,58-3,64; M: 3,88-3,56; (5) F: 3,75; (9) F: 4,1-3,3; M: 3,7-3,3; (14) F: 4-3,3; M: 3,55-3,05; (19) F: 4,24-3,6; M: 3,74-3,42; (20) F: 4,52-3,72; M: 4,2-3,36; (22) F: 4; 3,75-3,3; 4,22; M: 4-3,5; (34) F: 3,8; (47) F: 3,5; (63) F: 4,64-4,12; M: 3,86-3,61; (72) F: 4,51-4,03; M: 4,03-3,79; (73) F: 4,21; (91) F: ± 3,5; (98) F: 4,82-3,7; M: 3,94-3,39; (99) F: 4,32-4; M: 3,63; (101) F: 3,52; (142) F: 3,3; (143) F: 4,22; (199) F: 4,26-3,8; M: 3,88-3,57; (244) F: 3,6; (432) F: 3,77-3,68; (1122) F: 4,5; M: 4,02; {F: 3,30-4,82; M: 3,05-4,20}
The mean female size is 3.939 mm (n = 36; SD = 0.4185), and the mean male size is 3.681 mm (n = 22; SD = 0.2833). The size ratio (male : female) is 0.911 (n = 10; SD = 0.373), or ± 91 %. In samples, the sex ratio (female : male) is temporary 1.833. | | | | Rem.: | epi- bathypelagic. Considered as vetical migrator. Sargasso Sea: 0-1000 m (Deevey & Brooks, 1977, station "S"); 2000-1000 m (Harding, 1974). 369-616 at Station T-1 (E Tori Is., E Japan).
For Vervoort (1963 b, p.161) this species is closely related E. acuta Giesbrecht, 1892. For Park (1995, p.22) this species belongs to acuta Group. The two species can be separated by the following characters:
E. media : 1- Posterolateral thoracic border slightly produced and triangular in outline. 2- Tubercle on the left side of the genital somite rounded. On the right side a slight widening of the outline, terminating into a rounded protuberance just behind the middle of the genital somite. 3- A1 reach slightly beyond the end of the thorax. 4- Appendicular seta of the caudal rami lengthened and thickened at its base; remaining setae straight, as long as the urosome. (Cf the same characters in E. acuta. For the author, specimens from a number of different locations in all three great oceans have been examined for purposes of comparison and no significant geographical variations were found in their morphology. In 1978 this species was found between 26°30'S and 36°45'S in the southwestern Atlantic and between 33°01'S and 34°11'S in the southeastern Pacific (first records of this species from these areas)
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a seizing and masticating carnivorous.
After Sano & al. (2013, p.14, Table 2) atomic C:N ratio = 5.1 ±0.2, n = 8. | | | Last update : 15/11/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 17, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
