|
|
 |
|
Calanoida ( Order ) |
|
|
|
Clausocalanoidea ( Superfamily ) |
|
|
|
Euchaetidae ( Family ) |
|
|
|
Paraeuchaeta ( Genus ) |
|
|
| |
Paraeuchaeta elongata (Esterly, 1913) (F,M) | |
| | | | | | | Syn.: | Euchaeta elongata Esterly, 1913 (p.182, Descr.F, figs.F); Fleminger, 1967 a (tabl.1); Morris, 1970 (p.2300); Dagg & Littlepage, 1972 (p.162, weight, growth-RNA-DNA-protein); Gardner & Szabo, 1982 (p.262, figs.F,M); Yen J., 1982 (105, attack rates); 1983 (p.69, predation rates); 1985 (p.577, feeding, selective predation); Greene & Landry, 1985 (p.1408, feeding/prey selection); Mackas & Anderson, 1986 (p.115, Table 2); Rudyakov, 1986 (tab.2); Landry & Fagerness, 1988 (p.509, Table 1, 3, 5, 6, fig.1, 6, 8, grazing vs behaviour & morphology); Mauchline, 1998 (tab.33, 63); Park, W & al., 2004 (p.464, tab.1); Ohman, 1990 (p.257, fig.); Lewis & al., 2006 (p.501, figs.F, Table I); Papastephanou & al., 2006 (p.3078, Table 3); Waggett & Buskey, 2008 (p.111, Table 1); Yen J., 1982 (p.105, feeding); 1983 (p.69, feeding); Bailey & Yen, 1983 (p.71, predation); Ohman, 1990 (p.257, predation vs prey); Lewis & al., 2010 (p.695, Table I); Gusmao & al., 2013 (p.279, Table 3, sex-specific predation by fish);
Euchaeta japonica Marukawa, 1921; Campbell, 1929 (p.313, fig.M); 1934 (p.1, life history); Mori, 1937 (1964) (p.47, figs.F,M); Sewell, 1948 (p.391, 556, 560); Nakai, 1955 (p.12, chemical composition); Lewis & Ramnarine, 1969 (p.1347, figs. N, developement); Itoh, 1970 a (p.8: tab.2); Lewis & al., 1971 (p.1, chelators effect); Mackas & Sefton, 1982 (p.1179); Ohman, 1986 (p.673, fig. 11, predation); Lewis & Thomas, 1986 (p.1079, tidal transport); Bollens & Frost, 1991 (p.155); Hirakawa & al., 1992 (p.1, seasonal variability abundance); Ferrari & Dahms, 2007 (p.32, Rem. N, p.62, 66, 68, : copepodites, fig.23, Table III);
Pareuchaeta japonica : Minoda, 1958 (p.253, Table 1, 2, abundance); Morioka, 1972 a (p.314); Brodsky, 1948; Davis, 1949 (p.34, figs.F,M); Hirakawa & al., 1990 (tab.3); Hirakawa, 1991 (p.374, 376: fig.2);
? Pareuchaeta japonica : Brodsky, 1950 (1967) (p.210, figs.F,M, Rem.); Shih & al., 1971 (p.147); Bragina, 1999 (p.195); Pinchuk & Paul, 2000 (p.4, table 1, % occurrence);
Paraeuchaeta japonica : Vaupel-Klein, 1970 (p.4, 17); Peterson & Miller, 1975 (p.642, 650, occurrence); 1976 (p.14, Table 1, 2, 3, abundance vs interannual variations); 1977 (p.717, Table 1, seasonal occurrence); Coyle & al., 2014 (p.97, table 3, 10) Pinchuk & Paul, 2014 (p.4, table 1, % occurrence);
Pareuchaeta elongata : Tanaka, 1958 (p.355, figs.F,M); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio region, Table 6, 10); Tanaka & Omori, 1968 (p.234, figs.F,M); Vinogradov, 1968 (1970) (32, 52, 256); Minoda, 1971 (p.26); Heptner, 1971 (p.110, figs.F,M); Zvereva, 1976 a (p.70, fig.F); Coyle & al., 1990 (p.763); Hattori, 1991 (tab.1, Appendix); Yoo, 1991 (tab.1); Kim & al., 1993 (p.269); Kotani & al., 1996 (tab.2); Ikeda & Hirakawa, 1996 (p.261, life cycle); Park & Choi, 1997 (Appendix); Mauchline, 1998 (tab.20, 30, 42, 47); Mackas & al., 2004 (p.875, Table 2) | | | | Ref.: | | | Bradford & al., 1983 (p.20); Park, 1994 (p.320); 1995 (p.84, Rem.F, M, figs.F,M); Chihara & Murano, 1997 (p.800, Pl.106,112: F,M); Braga & al., 1999 (p.79, 84, tab.1, figs.6, 7, 8, Rem.: Biol. mol.); Jeong & al., 2011 (p.117, 120, figs.F,M, Rem.: genital aperture); Soh & al., 2013 (p.37, figs.F,M); Lewis A., 2014 (p.1200, Table I, II, figs. labrum-paragnaths) | 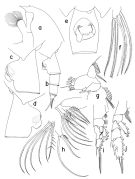 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.190, Fig.80]. Female: a, forehead (left side); b, urosome (left); c, d, e, genital somite (left, right, ventral, respectively); f, left caudal ramus (ventral); g, Mx1 (first inner lobe omitted), posterior; h, Mx2 (fifth lobe and endopod seoarated), posterior; i, P1 (anterior); j, P2 (anterior). Nota: Distal end of prosome produced into a lobular process. Laterally, left genital flange (Fig.80-c) low and elongate; posterior edge of genital field not produced distally into a lobe; right genital flange (Fig.80-d) large, produced distally into a large, rounded lobe exte,ding beyond posterior edge of genital field; ventrally, genital prominence (Fig.80-e) asymmetrical, right genital flange being much larger than left, and the entire prominence is turned counterclockwise. A1, A2, Md and Mxp as in P. malayensis; Mx1 with 1+9+3 setae on 1st inner lobe, and 1 seta each on 2nd and 3rd inner lobes; basis with 4 setae, without small outermost seta as found in P. malayensis; 1st endopodal segment with 3 setae; outer lobe with 9 setae, of which 2 proximal ones are short. In Mx2, 1 endopodal seta armed with long spinules in addition to dense rows of very short spinules. In P1 exopod, 1st segment not separated from 2nd, without an outer spine.
|
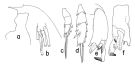 issued from : T. Park in Bull. Scripps Inst. Oceanogr. Univ. California, San Diego, 1995, 29. [p.191, Fig.81]. Male: a, forehead (left side); b, left caudal ramus (ventral); c, exopod of P1 (anterior); d, exopod of P2 (anterior); e, distal exopodal segments of left 5th leg (lateral, tilted clockwise; f, idem (medial). Nota: In P1, 1st exopodal segment fully separated from 2nd, without an outer spine.
|
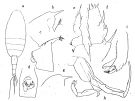 issued from : O. Tanaka in Publs Seto Mar. Biol. Lab., 1958, VI (3). [p.356, Fig.75]. As Pareuchaeta eliongata. Female: a, habitus (dorsal); b, forehead (lateral right side); c, last thoracic segment and genital somite (lateral left side); d, genital somite (ventral); e, exopod of P1; f, exopod of P2. Nota: The proportional lengths of the first three urosome segments in 0.01 mm. are97:52:43. Male: g, forehead (lateral); h, P5; i, terminal segments of left P5.
|
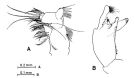 issued from : O. Tanaka & M. Omori in Publs Seto Mar. Biol. Lab., 1968, XVI (4). [p.235, Fig.9]. As Pareuchaeta elongata. Female: A, mx1. Male: B, terminal segments of left P5.
|
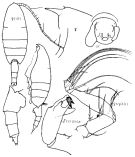 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.211, Fig.123]. As Pareuchaeta japonica. Female (from NW Pacif.): habitus (dorsal and lateral ledt side); forehead (lateral); Mp1 (J), Mxp; corner of the last thoracic segment and urosome (lateral right side); Gf, complex genital (ventral). Male: S5Le, distal segments of left P5.
|
 issued from : M.V. Heptner in Trudy. Inst. Okeanol., 1971, 92. [p.111, Fig.18]; As Pareuchaeta elongata. Female (from Kuril-Kamchatka Trench).
Nota: Formula of the exopod of P1 spines on the last two segments, successively : Aa = 0; Bb < BC Bb ≈ 7/8 BC); Cc > BC (Cc ≈ 1 1/3 BC). P2: Aa > AB (Aa = AB + 1/2 Bb); Bb < BC (Bb ≈ 3/4 BC); Cc < CD (Cc ≈ 8/9 CD). Variant: Aa = AB; Cc = CD. [cf. legend in drawings of Heptner, 1971, for Pareuchaeta rubra]. Male: ld, serrated lamella of left exopod of P5.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.22, Figs.3-11]. As Euchaeta japonica. Female: 3, habitus (lateral); 4, Mx2; 5, genital segment (ventral); 7, A1; 8, P2. Male: 6, habitus (lateral); 9, P5 (with parasite); 10-11, distal segment of left P5
|
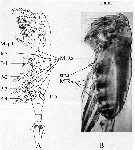 issued from : A.G. Lewis, S. Allen & G. Martens in Crustaceana, 2006, 79 (4). [p.503, Fig.1]; As Euchaeta elongata. Female (from off British Columbia coast): A, ventral view of an idealized calanoid copepod (modified from Owre & Foyo, 1967); B, prosome and portion of the genital somite (lateral view). Mxpd = maxilliped; MIR = median intersomitic ridge; IcS = intercoxal sclerite; P = swimming legs.
|
 issued from : A.G. Lewis, S. Allen & G. Martens in Crustaceana, 2006, 79 (4). [p.504, Fig.2, B]; Female: B, dissected 3rd pereiopod showing Ics and strut. IcS = intercoxal sclerite.
|
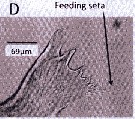
 issued from : J. Yen in Limnol. Oceanogr., 1985, 30 (3). [p.Fig.2]. As Euchaeta elongata. Scanning electron micrographs of the setae on the Mxp of an adult female where denticles are about 0.012 mm long. Scale bar: 0.010 mm.
|
 issued from : A.G. Lewis & A. Ramnarine inJ. Fish. Res. Bd. Canada, 1969, 26. [p.1349, Fig.1]. As Euchaeta japonica. Naupliar stages (from 49°17'06\\\"N, 123°50'W): a, 1st nauplius (dorsal view); b, 2nd nauplius (dorsal view); c-f, 3rd through 6th nauplii (ventral views). Nota: The egg + N1 stage is the most bcritical part of early developement . This is followed by rge N2, N3, N4, N5 and N6 stages with the last exhibiting the highest average survival. The egg to N2 stages are pre-feeding stages and appear to be dependent upon dissolved materials in the environment to supplement those not available or in low concentration in the deutoplasm or other stored nutrients. The N3 to N6 stages are feeding stages and the higher survival is felt to be due, at least in part, to the nutrient supply in the sea water. The N6, however, moults into the 1st copepodite, a change which is abrupt enough that Campbell (1934) termed it a metamorphosis. Whether or not the term is applicable is open to question but the preparation for the change may impose requirements on the stage that are not faced by the other feeding stages.
|
 issued from : M.-K. Jeong, H.-L. Suh & H.Y. Soh in Ocean Sci., 2011, 46 (2). [p.121, Fig.3]. Female (E Korea): A-B, habitus (dorsal and lateral, respectively); C, genital double-somite (dorsal); D, A1. Male; E-F, habitus (dorsal and lateral, respectively); G, P5; H, 2nd exopodal segment of left P5; I, A1.
Scale bars in miccrometers.
|
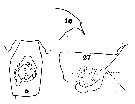 Issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1913, 11 (10). [Pl. 10, Figs. 5, 16, 27]. As Euchaeta elongata. Female (from S California): 5, genital segment and tip of last thoracic segment (ventral); 27, same (lateral); 16, forehead (lateral). genital segment length (ventral view) : 0.857 mm. Rostrum (from basal part to tip : 200 <µm. Nota:
|
 Issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1913, 11 (10). [Pl. 11, Fig. 37]. As Euchaeta elongata. Female: 37, 2nd and 3rd exopodal segments of P2. Nota: P2 with the outer marginal spine on the end of the 2nd exopodal segment reaches nearly to the tip of the 1st spine of the last segment; the middle spine of the the 3rd segment is long and extends to the distal margin of the segment, but the proximal and distal spines are short.
|
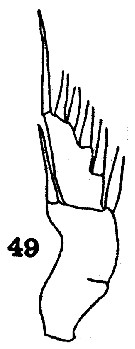 Issued from : C.O. Esterly in Univ. Calif. Publs Zool., 1913, 11 (10). [Pl. 12, Fig. 49]. As Euchaeta elongata. Female: 49, exopod of P1. Nota: Exopod with 2 segments, though there is some indication of the suture between the 1st and 2nd segments; the outer margin of the proximal division is deeply concave and bears 1 spine, which reaches to the end of the distal division.
|
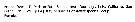 Paraeuchaeta elongata Female: 1 - See key to species Groups and independent species of Paraeuchaeta (p.30). 2 - Right genital flange large with extremely large posterior lobe (Fig.80-c).
|
 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.38, Fig.20]. Female (from Korean waters): A-B, habitus (dorsal and lateral, respectively); C, A1. Scale bars: A-B = 1000 µm; C = 500 µm.
|
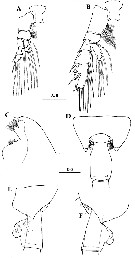 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.39, Fig.21]. Female: A, P1; B, P2; C, forehead (lateral); D, last thoracic segment and genital segment (dorsal); E-F, same (left and right sides, respectively). Scale bars: 500 µm.
|
 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.40, Fig.22]. Male: A-B, habitus (dorsal and lateral); C, A1. Scale bars: A-B = 1000 µm; C = 200 µm.
|
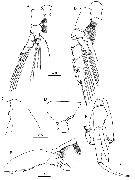 Issued from : H.Y. Soh, S.Y. Moon & J.H. Wi in Invertebrate Fauna of Korea (eds) Incheon: NIBR, 2013, 21 (28). [p.41, Fig.23]. Male: A, P1; B, P2; C, forehead (lateral); D, last thoracic segment and 1st urosomal segment (lateral); E, P5; F, distal segment of left P5. Scale bars: A, B = 200 µm; C-E = 500 µm; F = 100 µm.
|
 Issued from : A. Lewis in Crustaceana, 2014, 87 (10). [p.1216, Fig.9]. Paraeuchaeta elongata (Esterly, 1913): A, enlarged view of labrum and paragnath; B, confocal: ventral view from a posterior angle to show the position of the supralabral lobe relative to the labrum. The ridge just posterior to the setae is an artefact where two image frames were joined. See: http://booksandjournals.brillonline.com/content/journals/15685403. Nota: Relative to the body axis, the average angle of the labral-paragnath complex is + 14°; the complex faces down and forward. The anterior margin of the labrum proper is located a very short distance behind the first antenna, about 3% of the total body length. In lateral view, the labrum is narrow and clamshell-shaped hood. In ventral view, the bilobed lip is small; each of the tiny lobes bears a clump of setules. The labrum is contiguous with an anterior labral lobe that has a projecting anterior end with a cluster of long, naked setules (Park, 1995, called this structure the supralabrum). Both the labrum and the sub conical paragnaths are heavily sclerotized. The inner surface and part of the posterior surface of each paragnath is covered by setules. The base of each paragnath is connected to a heavily scleritized transverse bar that forms of the paragnath plate, and contains a cluster of small, spine like projections of teeth. The average length of the complex (not including the supralabral structure) is almost 12% of the total body length. With the base of the supralaral structure (not the projecting portion), the entire complex is 15% of the total body length. The length of the labrum base is slightly less than 40% of the length of the labrum-paragnath complex (excluding the supralabral lobe). The heigh of the labrum (including lip) is 1.2 times the length; the height of each paragnath is 62% of the length.
|
 Issued from : A. Lewis in Crustaceana, 2014, 87 (10). [p.1220, Fig.13, D]. Paraeuchaeta elongata (Esterly, 1913): D, gnathobase. See: http://booksandjournals.brillonline.com/content/journals/15685403.
| | | | | Compl. Ref.: | | | Shih & Marhue, 1991 (tab.2, 3); Dolganova & al., 1999 (p.13, tab.1); Logerwell & Ohman, 1999 (p.428, tab.1); Lenz & al., 2000 (p.338); Yamaguchi & Ikeda, 2002 a (p.1); Yamaguchi & al., 2002 (p.1007, tab.1); Yamaguchi & al., 2004 (p.480, tab.2, 3); Lavaniegos & Jiménez-Pérez, 2006 (tab.2, 4, Rem.); Ikeda & al., 2006 (p.1791, Table 2); Galbraith, 2009 (pers. comm.); Park & Ferrari, 2009 (p.143, Table 8, biogeography); Chiba & al., 2009 (p.1846, Table 1, occurrence vs temperature change); Homma & Yamaguchi, 2010 (p.965, Table 2); Homma & al., 2011 (p.29, Table 2, 3, 5, abundance); Batten & Walne, 2011 (p.1643, Table I, abundance vs temperature interannual variability); Abe & al., 2012 (p.100, temporal & vertical changes, gut content); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Ohashi & al., 2013 (p.44, Table 1, Rem.); Chiba S. & al., 2015 (p.968, Table 1: length vs climate); Ohtsuka & Nishida, 2017 (p.565, Table 22.1, Rem.) | | | | NZ: | 5 | | |
|
Distribution map of Paraeuchaeta elongata by geographical zones
|
| | | | | | 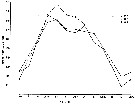 issued from : A.G. Lewis & A. Ramnarine inJ. Fish. Res. Bd. Canada, 1969, 26. [p.1355, Fig.3]. As Euchaeta japonica. issued from : A.G. Lewis & A. Ramnarine inJ. Fish. Res. Bd. Canada, 1969, 26. [p.1355, Fig.3]. As Euchaeta japonica.
Average numbers of eggs per egg cluster from January 1966 through August 1968 from Strait of Georgia (between Vancouver Island and the mainland.
Each point represents the average of 32 egg clusters.
Nota: The period of maximum egg production occurs during spring, at a time when large numbers of developmental stages of Calanus plumchrus are disponible as a food source. This increase in food to the female however, does not provide any demonstrable increase in the overall survival of the eggs. This suggests that the seasonal abundance of food is primarily responsible for the number of eggs produced and not their viability. |
 issued from : C.H. Greene & M.R. Landry in Ecology, 1985, 66 (5). [p.1413, Fig.6]. issued from : C.H. Greene & M.R. Landry in Ecology, 1985, 66 (5). [p.1413, Fig.6].
The vulnerability index as a function of prey type for P. elongata feeding on the different developmental stages of Calanus pacificus (circles) and adulut Pseudocalanus sp. (star).
The line drawings of the different prey types are drawn to scale with the scanning electron micrograph of Paraeuchaeta. |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.512, Fig.1]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.512, Fig.1].
Clearance rates of Euchaeta elongata (= Paraeuchaeta elongata) feeding on naupliar (closed symbols) and copepodid stages (open symbols).
Data from Yen (1983) (triangles), and Greene & Landry (1985) |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.514, Table 2]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.514, Table 2].
Comparison of the predatory potential of Euchaeta elongata (= Paraeuchaeta elongata) and Calanus pacificus. Feeding rate estimates are for copepod eliciting maximum clearance rates.
Euchaeta data from Yen (1983; 1985); Calanus data from Landry (1980).
Population abundance estimates are for Dabob Bay, Washington during summer months (B. Frost, pers. comm.).
F = predatory clearance rates (volume swept clear of prey) see Landry, 1978.
F = V [Ln (nc) - ln (ne] / NT.
V = container volume (liters), N = number of predators, T = duration of the experiments (days), nc and ne are respectively, the numbers of intact (living) prey recovered from control and experimental containers. See also Frost, 1972.
For each of the predatory copepods, a preferred prey was defined as the prey for which calculated clearance rate was highest, i.e., F max. I max = maximum of prey captured for copepod by day. |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.514, Table 3]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.514, Table 3].
Comparison of size of body and appendages for seven species of marine copepods. Relationships among predation rates, selection patterns, swimming speed and orientation, and morphology of prey sensory and capture appendages. |
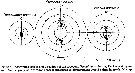 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.519, Fig. 8]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.519, Fig. 8].
Swimming speeds and directions of the copepods Neocalanus cristatus, Euchaeta elongata (= Paraeuchaeta elongata), Calanus pacificus. See also to Tortanus discaudatus, Corycaeus anglicus, Oithona spinifera (= Oithona setigera).
Swimming speeds measured as displacement over 2-s time intervals. 0° is up.
Study of the relationships among predation rates, selection patterns, swimming speed and orientation, and morphology of prey sensory and capture appendages.
The mean relative swimming speeds (S) of the predatory copepods and their preferred prey were calculated by the equation (Gerritsen, 1980).
S = [u + v)3 - [u - v)3 / 6uv.
u and v are respectively, the mean swimming speeds (mm/s) of the prey and the predator. |
 Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.522-523, Table 5]. Issued from : M.R. Landry & V.L. Fagerness in Bull. Mar. Sc., 1988, 43 (3) [p.522-523, Table 5].
Comparison of the predatory characteristics of seven planktonic marine copepods. Euchaeta elongata = Paraeuchaeta elongata.
F max is the mean, maximum clearance rate measured for the predator feeding on its preferred prey. Prey are developmental stages of common species of planktonic copepods.
F = female, M = male, C5 = stage 5. |
| | | | Loc: | | | China Seas (Yellow Sea, East China Sea), S & E Korea, Japan Sea, Japan (Izu, Onagawa, off Sanriku, Honshu: Toyama bay, off E, SE Hokkaido), Station ''H'', S Okhotsk Sea, Kuril Is., off SE Kuril Is. S Aleutian Is., Bering Sea, (S & western neritic) S Aleutian Basin, Pacif. (central subarctic), G. of Alaska, Icy Strait, Station 'P', British Columbia, Fjord system (Alice Arm & Hastings Arm), Haro Strait, Saanich Inlet, Dabob Bay, Portland Inlet, Vancouver Is., Dabob Bay, Friday Harbor, Port Susan, Puget Sound, Oregon (Yaquina, off Newport), California, W Baja California | | | | N: | 71 | | | | Lg.: | | | (3) F: 7,6-6; M: 6,4-5,9; (22) F: 6,5-6,3; M: 6,3-5,5; (23) F: 7,8-6,75; M: 7,05-6,4; (63) F: 7,21-6,28; M: 6,31-6,03; (91) F: ± 8; M: ± 8,4 ; (99) F: 7,39-6,81; M: 6,28; (144) F: 4,13; (208) F: 7,5-6; M: 6,8; (1106) F: 7,30-7,81; M: 5,92-6,51; (1174) F: 7,31-7,81; M: 5,92-6,51; (1259) F: 6,0; {F: 4,13-8,00; M: 5,50-8,40}
Chiba S. & al., 2015 (p.971, Table 1: Total length female (June-July) = 6.1 mm [optimal SST (°C) = 6.3].
(1259) F: 6, 0 (total length), 4,4 (prosome length); A1 = 4,5 mm. (total length). | | | | Rem.: | meso-bathypelagic.
For Tanaka (1958, p.357) the specimen might have been brought down into Sagami Bay by the cold intermediate water of the Oyashio current.
Park (1995, p.85) found this species along the coast of the whole of the northern Pacific ranging from 33°N off California northward to the Aleutian Islands, westward to the Kuril Islands, and southward along the Pacific coast of northern Japan down to 35°N. | | | Last update : 19/06/2023 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed July 03, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
