|
|
 |
|
Calanoida ( Order ) |
|
|
|
Lucicutiidae ( Family ) |
|
|
|
Lucicutia ( Genus ) |
|
|
| |
Lucicutia ovalis (Giesbrecht, 1889) (F,M) | |
| | | | | | | Syn.: | Isochaeta ovalis Giesbrecht,1889; 1892 (p.367, 773, Descr.F, figs.F); Giesbrecht & Schmeil, 1898 (p.112, Rem. F); Farran, 1926 (p.278, Rem.); Lysholm & al., 1945 (p.35); Sewell, 1948 (p.384, 569); C.B. Wilson, 1950 (p.241); Brodsky, 1950 (1967) (p.340, figs.F, Rem.); Tanaka, 1963 (p.53, figs.F,M, Rem.); Furuhashi, 1966 a (p.295, vertical distribution vs mixing Oyashio/Kuroshio region, Table 8, 9); Mazza, 1966 (p.71); Soenen, 1974 (p.120); Kovalev & Shmeleva, 1982 (p.84); Shih & Young, 1995 (p.70);
Lucicutia frigida Wolfenden, 1911 (p.320, figs.F); Hardy & Gunther, 1935 (1936) (p.180, Rem.); Sewell, 1948 (p.384, 570, 575); Brodsky, 1950 (1967) (p.340, figs.F, Rem.); Vervoort, 1957 (p.126, figs.F,M, Rem.); Minoda, 1971 (p.38, Rem.); 1972 (p.326); Morioka, 1972 a (p.314); Hopkins & Torres, 1988 (tab.1); Voronina & Kolosova, 1999 (p.71); Yamaguchi & al., 2002 (p.1007, tab.1);
Lucicutia ovaliformis Brodsky, 1950 (1967) (p.334, figs.F,M); Minoda, 1958 (p.253, Table 1, 2, abundance); Heotner, 1965 (p.11,65, figs.F,M, Rem.: variability): Gordon & al., 1985 (p.89, Table 2, fish diet); Yamaguchi al., 2002 (p.1007, tab.1); Homma & Yamaguchi, 2010 (p.965, Table 2); Homma & al., 2011 (p.29, Table 2, 3, 4, 5, abundance, feeding pattern: suspension feeders); Takenaka & al., 2012 (p.1669, fig.2, 3, Table 1, bioluminescence); Takenaka & al., 2013 (p.201, DNA sequences vs luciferase);
no Lucicutia ovalis Wolfenden, 1906 (p.28); 1911 (p.319); Farran, 1929 (p.263, fig.M); Chen & Zhang, 1965 (p.84,figs.F,M);
No Lucicutia ovalis Grice, 1962 (p.222, figs.F) | | | | Ref.: | | | Farran, 1926 (p.277); Grice, 1963 a (p.497, Rem.); Heptner, 1965 ( p.1175: Rem.); Grice & Hulsemann, 1965 (part., p.224); Hülsemann, 1966 (p.732, figs.F,M, Rem.); Vaupel-Klein, 1970 (p.4, 37); Bradford, 1971 b (p.24, figs.F,M); Heptner, 1971 (p.140, Rem. F,M); Björnberg & al., 1981 (p.648, figs.F,M); Gardner & Szabo, 1982 (p.360, figs.F,M); Razouls, 1994 (p.155, figs.F,M); Chihara & Murano, 1997 (p.829, tab.5, Pl.124: F,M); Bradford-Grieve & al., 1999 (p.883, 945, figs.F,M); Bradford-Grieve,1999 b (p.107, figs.F,M, Rem., figs.177, 191); Barthélémy, 1999 a (p.8); Boxshall & Halsey, 2004 (p.132: F; p.134: M); Conway & al., 2003 (p.90, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.344, figs.F,M, Rem.) | 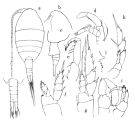 issued from : O. Tanaka in Publs Seto Mar. Biol. Lab., 1963, XI (1). [p.54, Fig.174]. As .
Female: a, habitus (dorsal); b, forehead (left lateral side); c, last thoracic segment and urosome (left lateral side); d, A2; e, Mxp; f, P1; g, P2; h, P5.
Nota: The urosome segments and furca are in the proportional lengths as 36 : 9 : 7 : 16 : 32 = 100.
Male: i, P5. Nota: The urosome segments and furca are in the proportional lengths as 17 : 13 : 12 : 8 : 13 : 37 = 100.
- Cephalosome without lateral protrusions.
- Caudal rami 1.4 times longer than wide; 2nd caudal seta about as long as urosome.
- Right A1 extends to posterior border of anal segment.
- P1 endopod 3-segmented.
- P5 right basipod 1 smooth; proximal part of inner margin of both basipode 2 with a pointed projection; right endopod 3-segmented.
|
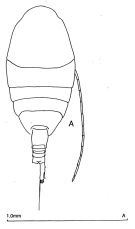 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.107, Fig.71]. Female (from 28°52'S, 178°05'W): A, habitus (dorsal). Nota: head without lateral protrusions. - Anal segment equal in length to urosome segment 3. -Caudal rami about 4 times longer than wide. - Genital segment, in lateral view, with a ventral protrusion on posterior half of segment, in dorsal view symmetrical. - A1 extends to posterior border of anal segment. - P1 endopod 3-segmented. - P5 endopod 2-segmented.
|
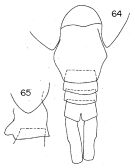 Issued from : K. Hülsemann in Bull. Mar. Sc., 1966, 16 (4). [p.718, Figs.64-65]. Female: 64, urosome (dorsal); 65, genital segment (lateral left side).
|
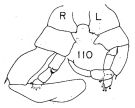 Issued from : K. Hülsemann in Bull. Mar. Sc., 1966, 16 (4). [p.726, Fig.110]. Male: 110, P5.
|
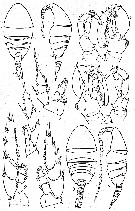 issued from : M.V. Heptner in Zool. Zh., 1972, 44 (8). [p.1169, Fig.1]. Female: 1-2, habitus (dorsal and lateral, respectively); 3, P1; 4, P2; 5, P4; 6, P5. Male: 7-8, P5; 9-10, habitus (dorsal and lateral, respectively).
|
 issued from : M.V. Heptner in Zool. Zh., 1972, 44 (8). [p.1172, Fig.2]. Female (variability): 1-5, genital segment (lateral); 6-9, idem (dorsal); 10, last thoracic segment and urosome (lateral); 11-21, endopod of P5; 22-25, endopod of P2.
|
 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Report-Ser. B, Vol. III, 1957 [Fig.111]. As Lucicutia frigida. Female (from 65°08'S, 80°58'E): a, postrior part cephalothorax and urosome (lateral); b, idem (dorsal); c, habitus (dorsal); d, right Mx1; e, right P1 (anterior).
|
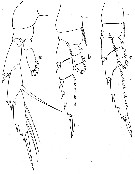 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Report-Ser. B, Vol. III, 1957 [Fig.112]. As Lucicutia frigida. Female: a-c, P1 to P4 (left legs, posterior).
|
 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Report-Ser. B, Vol. III, 1957 [Fig.113]. As Lucicutia frigida. Male: a, posterior part cephalothorax and urosome (lateral; b, idem (dorsal); c, P5; d, distal part of left A1.
|
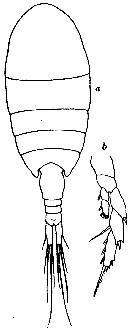 issued from : R.N. Wolfenden in Die Marinen Copepoden der Deutschen Südpolar-Expedition 1901-1903, 1911. [p.320, Fig.62]. As Lucicutia frigida. Female: a, habitus (dorsal); b, P5.
|
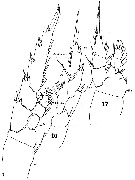 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. - Fauna Flora Golf. Neapel, 1892. Atlas von 54 Tafeln. [Taf. 29 , Figs.15, 16, 17 ]. As Isochäta ovalis. Female: 15, P3 (anterior view); 16, P5 (posterior view); 17, P1 (anterior view).
|
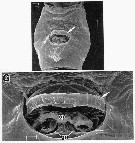 issued from : R.-M. Barthélémy in These Doc. Univ. Provence (Aix-Marseille I), 1999. [Fig.17, F, G]. Female: F, external ventral view genital double-somite; G, genital area. gn = gonoporal slits; sp = residual seminal products; opercular pad arrowed.
|
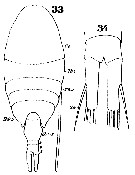 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.38, Figs.33, 34]. As Isochäta ovalis. Female: 33, habitus (dorsal); 34, caudal rami (ventral).
|
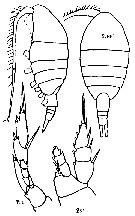 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.334, Fig.232]. As Lucicutia ovaliformis. Female (from NW Pacific): habitus (dorsal and lateral left side); S1, P1; S5, P5. Nota: Genital segment long and short 2nd and 3rd abdominal segments: Genital segment as long as remainder of abdomen excluding caudal rami; ventral process situated very distally, with the apex directed obliquely downward and backward. Caudal rami slightly shorter than combined length of 2nd, 3rd and 4th abdominal segments; three and 2/3 times longer than wide, . A1 slightly shorter than body. P1-with endopod 3-segmented; 3rd endopodite segment with sharp elongate inner distal corner. 2nd exopodite segment of P5 with greatly produced outer distal corner; larger bristle sityated at uppermost corner and directed obliquely upward
|
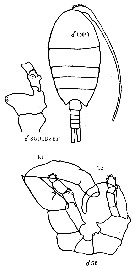 Issued from : K.A. Brodskii in Calanoida of the Far Eastern Seas and Polar Basin of the USSR. Opred. Fauna SSSR, 1950, 35 (Israel Program for Scientific Translations, Jerusalem, 1967) [p.334, Fig.232]. As Lucicutia ovaliformis. Male: habitus (dorsal); S5, P5 (Le = left leg; Ri = right leg) S5 Ri B2 En, basipodal segments and endopodal segments 1 and 2 of right leg of P5. Nota: Posterior thoracc corners produced slightly backwards, right corner larger than the left and turned slightly aside. P5 characteristic in structure, exopodite 2-segmented, much more massive on right right than on left leg; 1st basipodite with protuberances, protuberance of left leg strewn with blunt spinules, 2nd basipodite bearing simple protuberances; 2nd segment of right endopodite, and sometimes also outer margin of 2nd basipodite of same leg, bearing protuberances of various forms, strewn with blunt spinules.
|
 Lucicutia ovalis Lucicutia ovalis female: 1 - Characters following not combined : Prosome about 3 times longer than urosome. Cephalosome with slightly projecting and rounded anterior corners and well developed lateral spinous projections; anal somite about as long as wide; caudal rami 11.7 times longer than wide and bowed outwards at base, leaving elliptical space between rami proximally. 2 - P1 with 3-segmented endopod. 3 - Cephalosome without spinous projections . 4 - Genital double-somite symmetrical (dorsal view). 5 - Anal somite much shorter than caudal ramus. 6 - P5 with 2-segmented endopod. 7 - Caudal rami about 4 times longer than wide. Genital double-somite with marked ventral protrusion (lateral view).
|
 Lucicutia ovalis Lucicutia ovalis male: 1 - P1 with 3-segmented endopod. 2 - Cephalosome without lateral projections. 3 - Right A1 reaching at most 2 segments beyond tip of caudal rami. 4 - Caudal rami at most 7 times longer than wide. 5 - Inner margin of basis of right P5 smooth. 6 - Proximal part of inner margin of basis of both P5 with pointed process; endopod of right P5 3-segmented.
|
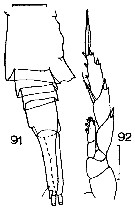 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.24, Figs.91, 92]. Female (from Ross Sea): 91, urosome (lateral, right side); 92, P5. Scale bar: 100 µm.
|
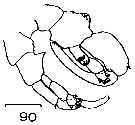 Issued from : J.M. Bradford in N.Z. Oceanogr. Inst., 1971, 206, Part 8, No 59. [p.24, Fig.90]. Male: 90, P5.
|
 Issued from : C. Razouls in Ann. Inst. océanogr., Paris, 1994, 70 (1). [p.155]. Caractéristiques morphologiques de Lucicutia ovalis femelle et mâle adultes. Terminologie et abbréviations: voir à Calanus propinquus.
| | | | | Compl. Ref.: | | | Heinrich, 1961 (Tab.4); Gaudy, 1962 (p.93, 99, Rem.: p.111) ; Grice, 1963 a (p.496); Gaudy, 1963 (p.26, Rem.); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Björnberg, 1963 (p.53, Rem.); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Grice & Hulsemann, 1967 (p.18); Vinogradov, 1968 (1970) (p.60, 78, 257, 268); Park, 1970 (p.477); Timonin, 1971 (p.281, trophic group); Roe, 1972 (p.277, tabl.1, tabl.2); Apostolopoulou, 1972 (p.328, 361); Björnberg, 1973 (p.344, 387); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Vives, 1982 (p.293); Greze & al., 1983 (p.17); Hopkins, 1985 (p.197, Table 1, gut contents); Brinton & al., 1986 (p.228, Table 1); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Wishner & Allison, 1986 (tab.2); Brenning, 1986 (p.11, Rem.); Madhupratap & Haridas, 1986 (p.105, tab.1); Atkinson & Peck, 1988 (p/.463,Table 1); Suarez & al., 1990 (tab.2); Madhupratap & Haridas, 1990 (p.305, fig.3: vertical distribution night/day; fig.7: cluster; Hattori, 1991 (tab.1, Appendix); Suarez & Gasca, 1991 (tab.2); Suarez, 1992 (App.1); Ashjian & Wishner, 1993 (p.483, abundance, species group distributions); Kotani & al., 1996 (tab.2); Hure & Krsinic, 1998 (p.102); Suarez-Morales & Gasca, 1998 a (p.110); Siokou-Frangou, 1999 (p.476); Atkinson & Sinclair, 2000 (p.46, 50, 51, 54, 55, zonal distribution); Razouls & al., 2000 (p.343, tab. 5, Appendix); Pakhomov & al., 2000 (p.1663, Table 2, transect Cape Town-SANAE antarctic base); Lopez-Salgado & al., 2000 (tab.1); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters); Hsiao & al., 2004 (p.326, tab.1); Shimode & al., 2005 (p.113 + poster); Hwang & al., 2006 (p.943, tabl. I); Koppelmann & Weikert, 2007 (p.266: tab.3); Hwang & al., 2007 (p.24); Dur & al., 2007 (p.197, Table IV); Schnack-Schiel & al., 2008 (p.1045: Tab.2); Morales-Ramirez & Suarez-Morales, 2008 (p.520); Fernandes, 2008 (p.465, Tabl.2); Galbraith, 2009 (pers. comm.); Tseng & al., 2009 (p.327, fig.5, feeding); Park & Ferrari, 2009 (p.143, Table 4, Appendix 1, biogeography); Dias & al., 2010 (p.230, Table 1); Mazzocchi & Di Capua, 2010 (p.426); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Jang M.-C & al., 2012 (p.37, abundance and seasonal distribution); Uysal & Shmeleva, 2012 (p.909, Table I); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Ward & al., 2014 (p.305, Table 6, 7, seasonal and abundance in the ''Discovery'' Investigations in the 1930s); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Mazzocchi & al., 2014 (p.64, Table 5, abundance); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones) | | | | NZ: | 18 | | |
|
Distribution map of Lucicutia ovalis by geographical zones
|
| | | | | | | | | | | | | | | 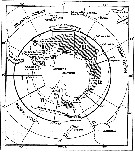 issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Reports - Ser. B, Vol. III, 1957 [Fig.114]. As Lucicutia frigida. issued from : W. Vervoort in B.A.N.Z. Antarctic Reseach Expedition, Reports - Ser. B, Vol. III, 1957 [Fig.114]. As Lucicutia frigida.
Chart showing the geographical distribution (white circle) in the seas surrounding the Antarctic continent.
Nota: In this chart the area frequented by whaling vessels has been hatched. The Antarctic circle (66°.5 S) has been drawn as a broken line. The numbers I to VI refer to the sectors into which the Antarctic seas are divided according to Mackintosh (1942) (after Vervoort, 1951). |
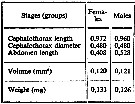 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 31 ]. Lucicutia ovalis (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 31 ]. Lucicutia ovalis (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
 Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3] Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3]
Lucicutia ovalis from Scotia Sea.
Median and interquartile ranges of copepods (nos /m2) in the five water zones; from north to south these are SAF Subantractic Front area, PFZ Polar frontal Zone, PF Polar Front area, AZ Antarctic Zone, WSC Weddell-Scotia Confluence area/ East Wind Drift.
Numbers on the plots are upper interquartiles where these could not be scaled. |
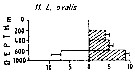 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.310, Fig.3]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.310, Fig.3].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 11- Lucicutia ovalis.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
| | | | Loc: | | | Antarct. (SW Atlant., Weddell Sea, Indian, SW Pacif., Ross Sea), South Georgia, sub-Antarct. (Indian), G. of Guinea, off Morocco-Mauritania, Canary Is., Portugal, Bay of Biscay, Azores, Brazil, off Rio de Janeiro, off Macaé, Caribbean Sea, Caribbean Colombia, G. of Mexico, off Bermuda (Station "S"), Sargasso Sea, off Cape Hatteras, Medit. (Alboran Sea, Marseille (very rare), Ligurian Sea, Tyrrhenian Sea, Strait of Sicily, Adriatic Sea, Aegean Sea, Johnston Bank, W Egyptian coast, N Lebanon Basin), Arabian Sea, Madagascar, Indian, Bay of Bengal, Andaman Sea, China Seas (East China Sea, South China Sea), off SW Taiwan, Taiwan (S, E, NW), S Korea, Korea Strait, Japan, off Sanriku, off SE Japan, Hokkaido, Station Knot, Kuril-Kamchatka, S Aleutian Basin, Bering Sea, S Aleutian Is., Station "P", off British Columbia, W Baja California, Gulf of California, W Costa Rica, Pacif. (equatorial), Galapagos-Ecuador, Chile, New Zealand (N North Island, off NE, NW); off W Kermadec Is. | | | | N: | 107 | | | | Lg.: | | | (10) F: 1,6-1,5; (21) F: 1,8-1,5; M: 1,5-1,2; (22) F: 1,7-1,6; M: 1,5-1,4; (23) F: 2-1,34; M: 1,72-1,22; (25) F: 1,76-1,58; M: 1,54-1,44; (26) F: 1,69; M: 1,55; (38) F: 1,44-1,34; M: 1,32-1,22; (47) F: 1,5; (102) F: 1,55-1,4; M: 1,85-1,3; (199) M: 1,29; (208) F: 1,9-1,8; M: 1,55; (237) F: 1,4; (920) F: 1,31; (991) F:1,50-1,80; M: 1,20-1,50; (1108) F: 1,78; {F: 1,31-2,00; M: 1,20-1,85}
| | | | Rem.: | epi, meso & bathypelagic.
Sampling depth (Antarct., sub-Antarct.) : 500-1000-2000 m. Sargasso Sea: 500-2000 m (Deevey & Brooks, 1977, Station "S"); 660-1340 m at Station S 2 in S Bösö (E middle Japan).
Male: The 3rd segment of the endopod of P2 has 6 setae in Tanaka's specimen, according to Giesbrecht 4 setae and Farran's specimen from the Bay of Biscay 5 setae.
After Björnberg (1963, p.53) this species is registered for the first time off Brazil.
After Bradford-Grieve (1999, p.107) A1 on the figured Southwest Pacific female extends as far as the posterior border of the caudal rami when held straight posteriorly.
Observed off Macaé (S Brazil) by Dias & al. (2010) in Intermediate Antarctic water.
Timonin (1971, p.282) considers the trophic interrelations in the equatorial and tropical Indian Ocean, and divides the plankters into 6 trophic groups from the litterature and the results of studies of mouth-parts structure and intestine content. This species is a coarse-filter feeder herbivorous.
All authors are not in accordance concerning the synonymy between L. ovalis (Giesbrecht, 1889) and L. ovaliformis Brodsky, 1950.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 4 corresponding to small filter feeding herbivorous and mixed feeding omnivorous (mostly broadcasters).
See in DVP Conway & al., 2003 (version 1) | | | Last update : 16/11/2020 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2026. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed June 22, 2026] © copyright 2005-2026 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
