|
|
 |
|
Calanoida ( Order ) |
|
|
|
Calanoidea ( Superfamily ) |
|
|
|
Paracalanidae ( Family ) |
|
|
|
Mecynocera ( Genus ) |
|
|
| |
Mecynocera clausi Thompson, 1888 (F,M) | |
| | | | | | | Syn.: | Leptocalanus filicornis Giesbrecht,1888;
Dolichocera tenuis : Bernard, 1958 a (p.195, figs.F,M)
no Calocalanus tenuis Farran, 1926;
Dolichocerea tenuis Bernard, 1963 b (p.160) | | | | Ref.: | | | I.C. Thompson, 1888 b (p.150, figs.F,M); Giesbrecht, 1892 (p.160, 773, figs.F); T. Scott, 1894 b (p.80, figs.F,M); Giesbrecht & Schmeil, 1898 (p.23, Rem. F); Wheeler, 1901 (p.167, fig.F, Rem.F); Thompson & Scott, 1903 (p.233, 242); Esterly, 1905 (p.137, fig.F); A. Scott, 1909 (p.25); Wolfenden, 1911 (p.202); Sewell, 1914 a (p.204); Pesta, 1920 (p.499); Früchtl, 1924 b (p.34); Sars, 1925 (p.23); Farran, 1926 (p.233); 1929 (p.208, 221); Rose, 1929 (p.15); Sewell, 1929 (p.60); Wilson, 1932 a (p.36, fig.F); Rose, 1933 a (p.72, figs.F); Dakin & Colefax, 1933 (p.204); Tanaka, 1935 (p.152, figs.F); Farran, 1936 a (p.79); Mori, 1937 (1964) (p.28, figs.F, juv.M); Dakin & Colefax, 1940 (p.96, figs.F); Farran & Vervoort, 1951 b (n°34, p.3, figs.F); Carvalho, 1952 a (p.142, figs.F); Tanaka, 1956 (p.272); Vervoort, 1957 (p.35, Rem.); Chiba & al., 1957 (p.306); 1957 a (p.11); Marques, 1959 (p.207); Brodsky, 1962 c (p.114, figs.F, M); Vervoort, 1963 b (p.116), Rem.; Paiva, 1963 (p.23, figs.F); Legaré, 1964 (p.33, figs.F); Chen & Zhang, 1965 (p.38, figs.F,M); Owre & Foyo, 1967 (p.40, figs.F); Park, 1968 (p.540); Vidal, 1968 (p.24, figs.F,M); Vilela, 1968 (p.12, Rem.M juv.); Ramirez, 1969 (p.51, fig.F, Rem.); Andronov, 1970 (p.980, Descr.M, figs.F,M, juv., Rem.); Corral Estrada, 1970 (p.135, Descr.M, figs.F,M, Rem); Corral, 1972 (p.37, 39, figs.F,M); Bradford, 1972 (p.34, figs.F); Razouls, 1972 (p.94, Annexe: p.26, figs.F,M); Marques, 1974 (p.14); Greenwood, 1977 (p.50); Dawson & Knatz, 1980 (p.4, figs.F); Björnberg & al., 1981 (p.624, figs.F,M); Marques, 1982 (p.758); Brodsky & al., 1983 (p.217, figs.F,M, Rem.); Nishida, 1989 (p.173, table 1, 2, 3: dorsal hump); Bradford-Grieve, 1994 (p.71, figs.F,M, fig.99); Chihara & Murano, 1997 (p.833, Pl.125: F,M); Bradford-Grieve & al., 1999 (p.878, 911, figs.F,M); Barthélémy, 1999 a (p.10, Fig.19); Conway & al., 2003 (p.152, figs.F,M, Rem.); Boxshall & Halsey, 2004 (p.137: figs.F,M); Avancini & al., 2006 (p.63, Pl. 32, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.908, figs.F,M, Rem.); Bradford-Grieve, 2008 (p.59, figs.M, Rem.: F, M); Andronov, 2014 (p.36, Fig.: A2) | 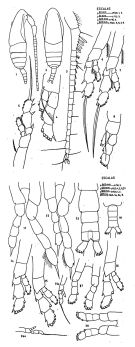 issued from : Corral Estrada J. in Tesis Doctoral, ser.A-n°129, sec. Biologicas; Publ. Fac .Ciencias, Madrid, 1970, p. 135. [Lamina 38, 39]. Male (from Canarias Is.): 1, 2, habitus (dorsal, lateral); 3, A1; 4, P1; 5, P2; 6, P3, 7, P4; 11, P5; 12, P5 (2nd form); 13, Urosome (dorsal). Nota: Head and 1st thoracic segment separate, 4th and 5th separate. Rostum with 2 filaments. Head and 1st thoracic segment with a stronf depresion. The urosomal segments and furca in the proportional lengths 13:30:14:14:13: 16 = 100. A1 24 segmented ( (1+2 fused, 3-4-5 fused, 6 to 24 free), exceeds the caudal rami by the last 3 segments. Female: 8, P1; 14, P2; 15, 15a, P3; 16, 16a, P4; 17, 19, P5; 18, Urosome (juvenile form).
|
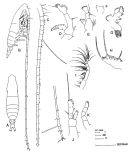 issued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). New Zealand Oceanographic Institute Memoir, 102, 1994. [p.72, Fig.38]. Female: A, habitus (dorsal); B, idem (lateral right side); C, A1; D, A2; E, Md (mandibular palp); F, Md (distal part of mandibular blade); G, Mx1; H, Mx2; I, Mxp; J, P1; K, P5. Nota: No males were detected in the Southwest Pacific samples examined.
|
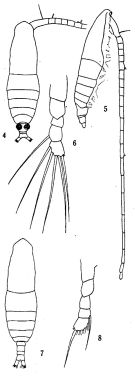 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.8, 4-8]. Female (from E China Sea): 4, habitus (dorsal); 5, idem (lateral right side); 6, right P5 (posterior). Male: 7, habitus (dorsal); ; 8, P5 (posterior). Nota: The figure 8 is doutful in this species.
|
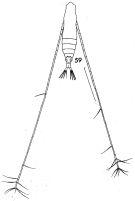 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.56, Lam. IX, fig.59 ]. Female (from off Mar del Plata): 59, habitus (dorsal). Scale bar in mm: 0.6. Nota: Left A1 longer than the right (proportion: 49:43)
|
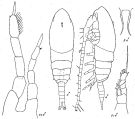 issued from : K.A. Brodsky, N.V. Vyshkvartzeva, M.S. Kos & E.L. Markhaseva in Opred Fauna SSSR, 1983, 135. [p.218, Fig.102. Male after Andronov, 1970 and 1973 (redrawn)].
|
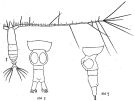 issued from : K.A. Brodsky, N.V. Vyshkvartzeva, M.S. Kos & E.L. Markhaseva in Opred Fauna SSSR, 1983, 135. [p.217, Fig.101]. Female from Androbnov, 1970. Nota: A1-24-segmented.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.11, Figs.1-3]. Female: 1, habitus (dorsal); 2, P1; 3, P5.
|
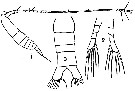 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.23, Figs.1-3]. Male (immature): 1, habitus (dlateral right side); 2, P5; 3, urosome (dorsal). Nota: Such male was described as adult by C.B. Wilson (1932).
|
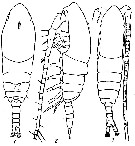 issued from : V.N. Andronov in Zool. Zh., 1970, 49 (7). [p.981, Fig.1]. Male (from Namibia, comm. pers.): 1, 2, habitus (dorsal and lateral, respectively). Nota: A1 20-segmented Male copepodite V: 3, habitus (dorsal).
|
 issued from : V.N. Andronov in Zool. Zh., 1970, 49 (7). [p.982, Fig.2]. Female: 1, 2, urosome (dorsal and lateral, respectively). Female copepodite V: 3, 4, urosome (dorsal and lateral, respectively). Male: 9, P1; 10, P5; 13, rostrum. Male copepodite V: 11, P5. Copepodite IV: 12, P5. 5, 6, urosome (dorsal and lateral, respectively).
|
 issued from : V.N. Andronov in Zool. Zh., 1973, 52 (11). [p.1719]: P5 Male: a, left leg; b, right leg.
|
 issued from : J.M. Bradford-Grieve in Zootaxa, 2008, 1852. (p.61, Fig.1]. Male (from 20°26'S, 5°55'E): A, habitus (lateral); B, P1 (anterior); C, P2 (anterior); D, P5 (anterior). Scale bars 0.1 mm.
|
 issued from : J.M. Bradford-Grieve in Zootaxa, 2008, 1852. (p.62, Fig.2]. male: A, left A2; B, left Md; C, left Mx1; D, right Mx2; E, right Mxp. Scale bar 0.1 mm. Nota: Md with degenerate gnathobase; palp well developed with endopod 2-segmented, exopod with 6 setae. Mx1reduced with degenerate inner lobes, endopod with 2 small setae, exopod with 8 setae, praecoxa epipodite with 6 large setae. Mx2 reduced to a small, knob-like remnant. Mxp sexually dimorphic; inner setae of syncoxa, basis and endopod segments absent or very reduced; 1 terminal and 2 outer endopod setae ellarged, plumose, 2 outer setae directed towards base of limb (as is typical of Paracalanidae, Calanidae and Eucalanidae).
|
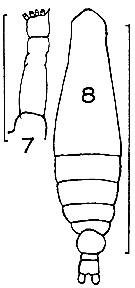 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.35, Fig.5 (7-8)]. Female (from Kaikoura, New Zealand): 7, P5; 8, habitus (dorsal). Scale bazrs: 1 mm (8); 0.1 mm (7).
|
 issued from : C. Razouls in Th. Doc. Etat Fac. Sc. Paris VI, 1972, Annexe. [Fig.28]. Female (from Banyuls, G. of Lion): A, habitus (dorsal). Male: B, urosome (dorsal); C, P5.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19. Atlas von 54 Tafeln. [Taf.11 , Fig.43-45 ]. Female: 43, P5 (anterior view); 44, P2 (postrior view); 45, Mxp (posterior view).
|
 issued from : R.-M. Barthélémy in These Doct. Univ. Provence (Aix-Marseille I), 1999. [Fig.19, D]. Female (from unknown localization): external ventral view genital double-somite. go = genital operculum. Scale bar: 0.020 mm.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf. 35, Figs.21, 22]. Female: 21, urosome (ventral); 22, forehead (ventral).
|
 issued from : P.E. Lapernat & C. Razouls in Vie Milieu, 2002, 52 (1). [p.21, Pl. II, fig.4]. Masticatory edge of Md gnathobase female (from off Malta, Mediterranean Sea).
|
 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.36, Fig. 9, 14]. Mecynocera clausi after Bradford-Grieve, 1994. 14, exopod of A2.
|
 Issued from : V.N. Andronov in Russian Acad. Sci. P.P. Shirshov Inst. Oceanol. Atlantic Branch, Kaliningrad, 2014. [p.89, Fig.23: 2]. Mecynocera clausi after Bradford-Grieve, 2008. Mxp male.
| | | | | Compl. Ref.: | | | Pearson, 1906 (p.8); Wilson, 1932 (p.25); 1942 a (p.193); Massuti Alzamora, 1942 (p.87); Lysholm & al., 1945 (p.10); Oliveira, 1945 (p.191); Sewell, 1948 (p.407, 412, 414, 422, 433, 436, 442, 450, 453, 459, 468, 469, 478, 481, 489, 491, 493); C.B. Wilson, 1950 (p.262); Krishnaswamy, 1953 (p.113); Yamazi, 1958 (p.147, Rem.); Deevey, 1960 (p.5, Table II, annual abundance) ; Fagetti, 1962 (p.11); Grice & Hart, 1962 (p.287, table 3, table 4: abundance); Gaudy, 1962 (p.93, 99, Rem.: p.102); Ganapati & Shanthakumari, 1962 (p.7, 15); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Duran, 1963 (p.13); Shmeleva, 1963 (p.141); V.N. Greze, 1963 a (tabl.2); Gaudy, 1963 (p.20, Rem.); Björnberg, 1963 (p.24, Rem.); Unterüberbacher, 1964 (p.17); De Decker, 1964 (p.16, 22, 30); De Decker & Mombeck, 1964 (p.13); Grice & Hulsemann, 1965 (p.223); Shmeleva, 1965 b (p.1350, lengths-volume -weight relation); Pavlova, 1966 (p.43); Neto & Paiva, 1966 (p.22, Table III); Furuhashi, 1966 a (p.295, vertical distribution vs mixing Oyashio/Kuroshio region); Mazza, 1966 (p.69); 1967 (p.327, 329, fig.65); Ehrhardt, 1967 (p.737, 887, fig.11, geographic distribution), Rem.); Fleminger, 1967 a (tabl.1); Grice & Hulsemann, 1967 (p.14); De Decker, 1968 (p.45); Vinogradov, 1968 (1970) (p.78); Séguin, 1968 (p.488); Delalo, 1968 (p.137); Berdugo & Kimor, 1968 (p.447); Evans, 1968 (p.13); Dowidar & El-Maghraby, 1970 (p.268); Park, 1970 (p.474); Deevey, 1971 (p.224); Gamulin, 1971 (p.381, tab.2); Paulmier, 1971 (p.168); Binet & al., 1972 (p.71); Roe, 1972 (p.277, tabl.1, tabl.2); Ibanez & Seguin, 1972 (p.81, annual cycle, multivarite analysis); Apostolopoulou, 1972 (p.327, 334); Heinrich, 1973 (p.95, fig.3); Björnberg, 1973 (p.309, 387); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Vives & al., 1975 (p.35, tab.II, III); Peterson & Miller, 1976 (p.14, Table 1, 3, abundance vs interannual variations); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Tranter, 1977 (p.596); Carter, 1977 (1978) (p.35); Peterson & Miller, 1977 (p.717, Table 1, seasonal occurrence); Comaschi Scaramuzza, 1978 (p.17); Dessier, 1979 (p.134, 201, 204); Porumb, 1980 (p.168); Star & Mullin, 1981 (p.1322, abundance); Brenning, 1982 (p.7, Rem.); Rudyakov, 1982 (p.208, Table 2); Vaissière & Séguin, 1980 (p.23, tab. 2); Kovalev & Shmeleva, 1982 (p.82); Vives, 1982 (p.290); Turner & Dagg, 1983 (p.16); Dessier, 1983 (p.89, Tableau 1, Rem., %); Binet, 1984 (tab.3); Tremblay & Anderson, 1984 (p.4, Rem.); De Decker, 1984 (p.315, 354: chart); Scotto di Carlo & al., 1984 (1042); Guangshan & Honglin, 1984 (p.118, tab.); Regner, 1985 (p.11, Rem.: p.26); Brenning, 1985 a (p.28, fig.9, Table 2); Jansa, 1985 (p.108, Tabl.I, , II, III, IV); Binet, 1985 (p.85, tab.3); Moraitou-Apostolopoulou, 1985 (p.303, occurrence/abundance in E Mediterranean Sea, Rem.: p.315); Brinton & al., 1986 (p.228, Table 1); Madhupratap & Haridas, 1986 (p.105, tab.1); Lozano Soldevilla & al., 1988 (p.57); Dessier, 1988 (tabl.1); Cervantes-Duarte Hernandez-Trujillo, 1989 (tab.3); Pancucci-Papadopoulou & al., 1990 (p.199); Suarez & al., 1990 (tab.2); Madhupratap & Haridas, 1990 (p.305, fig.6: vertical distribution night/day; fig.7: cluster, Rem: p.318); Echelman & Fishelson, 1990 a (tab.2); Yoo, 1991 (tab.1); Suarez & Gasca, 1991 (tab.2); McKinnon, 1991 (p.471); Suarez, 1992 (App.1); Seguin & al., 1993 (p.23, 26: Rem.); Kimmerer, 1993 (tab.2); Hays & al., 1994 (tab.1); Kouwenberg, 1994 (tab.1); Heinrich, 1995 (tab.1); Shih & Young, 1995 (p.69); Kotani & al., 1996 (tab.2); Errhif & al., 1997 (p.422); Park & Choi, 1997 (Appendix); Hure & Krsinic, 1998 (p.28, 100); Noda & al., 1998 (p.55, Table 3, occurrence); Gilabert & Moreno, 1998 (tab.1, 2); Alvarez-Cadena & al., 1998 (tab.3,4); Suarez-Morales & Gasca, 1998 a (p108); Mauchline, 1998 (tab.30, 58); Lapernat, 1999 (p.14, 55); Lopes & al., 1999 (p.215, tab.1); Dolganova & al., 1999 (p.13, tab.1); Neumann-Leitao & al., 1999 (p.153, tab.2); Onishchik, 1999 (p.76, fig.2); Lopez-Salgado & al., 2000 (tab.1); Razouls & al., 2000 (p.343, Appendix); Pakhomov & al., 2000 (p.1663, Table 2, transect Cape Town-SANAE antarctic base); Moraitou-Apostolopoulou & al., 2000 (tab.I, fig.8); Lapernat & Razouls, 2001 (p.123, tab.1); Hidalgo Escribano, 2001 (p.159, tab.2); Hidalgo & Escribano, 2001 (p.158, fig.5); Holmes, 2001 (p.40); Sameoto & al., 2002 (p.12); Zerouali & Melhaoui, 2002 (p.91, Tableau I); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, figs.5, 6); Labat & al., 2002 (p.741, tab.3); Paffenhöfer & Mazzocchi, 2003 (p.1139, figs.7, 8); Vukanic, 2003 (139, tab.1); Shimode & Shirayama, 2004 (tab.2); Hsiao & al., 2004 (p.326, tab.1); Chang & Fang, 2004 (p.456, tab.1); Daly Yahia & al., 2004 (p.366, fig.4); Lan & al., 2004 (p.332, tab.1); Fernandez de Puelles & al., 2004 (p.654, fig.7); CPR, 2004 (p.56, fig.162); Lo & al., 2004 (p.89, tab.1); Vukanic & Vukanic, 2004 (p.9, tab. 2, 3); Obuid Allah & al., 2005 (p.123, occurrence % vs metal contamination); Shimode & al., 2005 (p.113 + poster); Berasategui & al., 2005 (p.485, tab.1); Berasategui & al., 2006 (p.485: fig.2); Zuo & al., 2006 (p.163: tab.1); Isari & al., 2006 (p.241, tab.II); Hwang & al., 2006 (p.943, tabl. I); Dias & Araujo, 2006 (p.54, Rem., chart); Lavaniegos & Jiménez-Pérez, 2006 (p.147, tab.2, 3, Rem.); Zervoudaki & al., 2006 (p.149, Table I); Koppelmann & Weikert, 2007 (p.266: tab.3); Hwang & al., 2007 (p.24); Dur & al., 2007 (p.197, Table IV); Cornils & al., 2007 (p.278, Table 2); Fernandez de Puelles & al., 2007 (p.348, fig.7); Valdés & al., 2007 (p.103: tab.1); Khelifi-Touhami & al., 2007 (p.327, Table 1); McKinnon & al., 2008 (p.844: Tab.I, p.848: Tab. IV); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.6); Morales-Ramirez & Suarez-Morales, 2008 (p.520); Fernandes, 2008 (p.465, Tabl.2); Muelbert & al., 2008 (p.1662, Table 1, 3); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Tseng L.-C. & al., 2008 (p.153, Table 2, occurrence vs geographic distribution); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Pagano, 2009 (p.116); Miyashita & al., 2009 (p.815, Tabl.II); Tseng & al., 2009 (p.327, fig.5, feeding); Lidvanov & al., 2010 (p.356, Table 3); Hernandez-Trujillo & al., 2010 (p.913, Table 2); Cornils & al., 2010 (p.2076, Table 3); Schnack-Schiel & al., 2010 (p.2064, Table 2: E Atlantic subtropical/tropical); Hidalgo & al., 2010 (p.2089, Table 2); Dias & al., 2010 (p.230, Table 1, fig.7 b); Mazzocchi & Di Capua, 2010 (p.426); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Nowaczyk & al;, 2011 (p.2159, Table 2); Hsiao S.H. & al., 2011 (p.475, Appendix I); Pepin & al., 2011 (p.273, Table 2, seasonal abundance); Selifonova, 2011 a (p.77, Table 1, alien species in Black Sea); Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Mazzocchi & al., 2011 (p.1163, fig.6, long-term time-series 1984-2006); Isari & al., 2011 (p.51, Table 2, abundance vs distribution); Mazzocchi & al., 2012 (p.135, annual abundance 1984-2006); Shiganova & al., 2012 (p.61, Table 4); Uysal & Shmeleva, 2012 (p.909, Table I); Salah S. & al., 2012 (p.155, Tableau 1); DiBacco & al., 2012 (p.483, Table S1, ballast water transport); Zizah & al., 2012 (p.79, Tableau I, Rem.: p.86); Miloslavic & al., 2012 (p.165, Table 2, transect distribution); Aubry & al., 2012 (p.125, fig.8 b, interannual variation); Dorgham & al., 2012 (p.473, Table 4: abundance vs season); Hidalgo & al., 2012 (p.134, Table 2) ; Gubanova & al., 2013 (in press, p.4, Table 2); Belmonte & al., 2013 (p.222, Table 2, abundance vs stations); Cornils & Blanco-Bercial, 2013 (p.861, Table 1, molecular analysis, figs.3, 4, 5); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Hirai & al., 2013 (p.1, Table I, molecular marker); Terbiyik Kurt & Polat, 2013 (p.1163, Table 2, seasonal distribution); Siokou & al., 2013 (p.1313, fig.4, 8, biomass, vertical distribution); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Mazzocchi & al., 2014 (p.64, Table 3, 4, 5, spatial & seasonal composition %); Zaafa & al., 2014 (p.67, Table I, occurrence); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Zakaria & al., 2016 (p.1, Table 1); Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); Ben Ltaief & al., 2017 (p.1, Table III, Summer relative abundance); El Arraj & al., 2017 (p.272, table 2); Marques-Rojas & Zoppi de Roa, 2017 (p.495, Table 1); Benedetti & al., 2018 (p.1, Fig.2: ecological functional group); Belmonte, 2018 (p.273, Table I: Italian zones); Chaouadi & Hafferssas, 2018 (p.913, Table II: occurrence); Hure M. & al., 2018 (p.1, Table 1: abundance, % composition, Table 2: correlations); Palomares-Garcia & al., 2018 (p.178, Table 1: occurrence); Bode & al., 2018 (p.66, fig.5, vertical distribution %, Rem). Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions). | | | | NZ: | 22 | | |
|
Distribution map of Mecynocera clausi by geographical zones
|
| | | | | | | | | | | | | | | 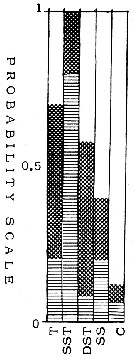 issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.25, Fig.11]. issued from : T.S.K. Björnberg in Bol. Inst. Oceanogr., Sao Paulo, 1963, XIII (1). [p.25, Fig.11].
Probability of occurrence of Mecynocera clausi in different environments.
T: Tropical waters (above 36.00 p.1000 salinity and above 20°C temperature); SST: Surface subtropical waters (salinity around 36 p.1000 and temperature 18°C or less); DST: Deeper shelf waters (salinity between 34 and 36, temperature under 20°C p.1000); SS: Surface shelf waters (same salinity and temperature above 20°C);
C: coastal waters with low salinity and variable temperature.
In each column no shading means no probability of finding the species in the samples from this environment; horizontal shading indicates the probability of finding the sepecies in percentages less than one in samples from this environment; cross shading indicates the probability of finding it in percentages higher than one and black shading represents the probability of finding it in the largest percentages of the total number of copepods.
For Björnberg (1963, p.24) this species was present in nearly all samples of the most saline and warm waters off Brazil; it is a surface copepod, because it did not occur in the samples collected in deep layers and its environmental optimum lies in cooler temperatures between 18 and 10°C. |
 issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 6]. Mecynocera clausi (from South Adriatic). issued from : A.A. Shmeleva in Bull. Inst. Oceanogr., Monaco, 1965, 65 (n°1351). [Table 6: 6]. Mecynocera clausi (from South Adriatic).
Dimensions, volume and Weight wet. Means for 50-60 specimens. Volume and weight calculated by geometrical method. Assumed that the specific gravity of the Copepod body is equal to 1, then the volume will correspond to the weight. |
 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.313, Fig.6].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 70- Mecynocera clausi.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
| | | | Loc: | | | Cosmopolite (tropical, subtropical). sub-Antarct. (South of South Africa, Indian, S Tasmania), South Africa (E & W), Tristan da Cunha, Namibia, off Angola, Baia Farta, Congo, G. of Guinea, Ivorian shelf, Dakar, Morocco-Mauritania, Cap Ghir, Canary Is., off Madeira, Azores, Portugal, Argentina, off Rio de La Plata (shelfbreak), Uruguay (continental shelf), Brazil, off Rio de Janeiro, Campos Basin, off Macaé, off Natal, Caribbean Colombia, Bahia de Mochima (Venezuela), Caribbean Sea, Jamaica, Yucatan, G. of Mexico, Florida, off Bermuda, Station "S", Sargasso Sea, off Bermuda: Station "S" (32°10'N, 64°30'W), Delaware Bay (outside), Chesapeake Bay, New York, Long Island, Woods Hole, off Nova Scotia E, Newfoundland, Flemish Cape, off Ireland (SW & W), English Channel, Belon estuary, Bay of Biscay, Ibero-moroccan Bay, off W Tangier, Medit. (M'Diq, Alboran Sea, Habibas Is., Sidi Fredj coast, Gulf of Annaba, El Kala shelf, Baleares, Banyuls, Marseille, Ligurian Sea, Tyrrhenian Sea, Napoli, Strait of Messina, Gulf of Taranto, NW Tunisia, G. of Gabes, Malta, Adriatic Sea, Ionian Sea, Aegean Sea, Thracian Sea, Iskenderun Bay, Lebanon Basin, W Egyptian coast, Alexandria, Black Sea), G. of Aqaba, off Sharm El-Sheikh, Safâga, Red Sea, off Madagascar S, Rodrigues Is., Seychelles, Indian, Sri Lanka, India (Lawson's Bay), S Indian (subtropical convergence), Bay of Bengal, Indonesia-Malaysia, SW Sulawesi, SW Celebes, Australia (W, North West Cape), China Seas (Yellow Sea, East China Sea, South China Sea), off SW Taiwan, Taiwan (S, E, Kaohsiung Harbor, NW, N: Mienhua Canyon), S Korea, Japan Sea, Japan (Kuchinoerabu Is., Tanabe Bay, Sagami Bay), Kuroshio zone, Bering Sea, Alaska, Pacif. (tropical), Oregon (off Newport), California, W Baja California, Bahia Magdalena, La Paz, Gulf of California, W Costa Rica, Clipperton Is., Pacif. (W equatorial), Pacif. (N Central Gyre), Australia (Great Barrier), New Caledonia, New Zealand (Kaikoura), S Pacif. (NPFZ), Galapagos-Ecuador, Peru, Chile (N-S, Mejillones Peninsula, off Santiago)
Data from Cornils & Blanco-Bercial (2013): sp.1: 23°14'S:; 08°14'E. sp.2: 05°07'S; 119°21'E. | | | | N: | 302 | | | | Lg.: | | | (14) F: 1-0,95; (38) F: 1,2; (45) F: 1,15-1; M: 0,9-0,75; (47) F: 1-0,92; (54) F: 1,02; (72) F: 1,15-1,05; (73) F: 1,03-0,99; (91) F: ± 1,0; (104) F: 1,25; (116) F: 1,2; (125) F: 0,99-0,94; (131) F: 1,21-0,9; M: 1,12-1,04; (142) F: 1-0,9; (180) F: 1-0,93; M: 1,05-0,94; (187) F: 1,08-1,01; M: 1,08-1,01; (199) F: 1,06-0,91; (202) F: 0,92-1,1; M: 0,94- 1,05; (237) F: 1,0-1,1; (254) F: ± 1,2; (290) F: 0,95-1; M: 0,95; (327) F: 1,06-0,94; M ?juv.: 1,02-0,97; (332) F: 1; (335) F: 1,07-0,99; M: 0,99; (340) F: 1,05; (404) M: 1,05-0,94; (432) F: 1,25-0,98; (786) F: 1,29-1,09; (795) F: 1; M: 1; (920) F: 0,99; (991) F: 0,92-1,21; M: 0,94-1,12; (1112) F: 0,96-1,064; {F: 0,90-1,29; M: 0,75-1,12} | | | | Rem.: | Oceanic. Epipelagic, (2000 m: off Malta). Sargasso Sea: 0-500 m.
Sampling depth (sub-Antarct.) : 0-600 m.
The male would be described correctly for the first time by Corral Estrada (1970) and Andronov (1970, 1973). The drawings of male by Chen & Zhang (1965, Pl.8, figs.7-8) are very doubtful.
For Andronov (1970, p.985) a comparison of the 5th copepodite stage of males M. clausi with males and females Dolichocera tenuis (= Calocalanus tenuis) suggests that these two species are synonyms. An advisability of the genus Calocalanus division into three genera and their isolation as an independant family is doubtful. It is suggested to include into the family Paracalanidae the genera Calocalanus and Mecynocera. In 1973, the author erected the monotypic family Mecynoceridae based on the presence of the dorsal cephalic hump on the cephalothorax of the male, the reduced setation of both rami of legs P2 to P4, and the location of the male genital aperture on the right side.
For Bradford-Grieve (2008, pp.59-63) this species is transferred to the Paracalnidae on the basis of the form of the exopod of the male A2 and Mxp; details of the setation of the mouthparts and swimming legs and segmentation of the female urosome.
After Lapernat & Razouls (2002, p.19) the Itoh's index value = 746.2 (number of teeth: 11) on the right Md, and 799.6 on the left Md (number of teeth: 10).
After Ehrhardt (1967, p.888, 893) the species is found in the southwest Tyrrhenian Sea in water salinity between 37.21-38.23 p.1000 (mainly between 37.77- 37.91) and for the Oxygen between 3.44-6.00 cm3/l (mainly between 4.84-5.55).
After Moraitou-Apostolopoulou (1985 p.315), this species which is abundant throught the Mediterranean Sea, is found in restricted numbers in the north basin of the Aegean Sea.
After Benedetti & al. (2018, p.1, Fig.2) this species belonging to the functional group 7 corresponding to small filter feeding herbivorous.
See in DVP Conway & al., 2003 (version 1) | | | Last update : 25/10/2022 | |
|
|
 Any use of this site for a publication will be mentioned with the following reference : Any use of this site for a publication will be mentioned with the following reference :
Razouls C., Desreumaux N., Kouwenberg J. and de Bovée F., 2005-2025. - Biodiversity of Marine Planktonic Copepods (morphology, geographical distribution and biological data). Sorbonne University, CNRS. Available at http://copepodes.obs-banyuls.fr/en [Accessed August 23, 2025] © copyright 2005-2025 Sorbonne University, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
