

|
|

Famille
Centropagidae Giesbrecht, 1892
Ref.:
Giesbrecht, 1892 (p.58); Giesbrecht & Schmeil, 1898 (p.52); Sars, 1902 (1903) (p.73); Esterly, 1905 (p.170); van Breemen, 1908 a (p.91); Gurney, 1931 a (p.85); Rose, 1933 a (p.183); Mori, 1937 (1964) (part., p.8, 55, Genera Key); Brodsky, 1950 (1967) (p.314); Gonzalez & Bowman, 1965 (p.247); Bayly & Arnott, 1969 (Rev., p.190, 194, Genera Key); Lee, 1972 (p.1, spematophore, coupling device); Andronov, 1974 a (p.1005); Kiefer, 1978 (p.55); Bowman & Abele, 1982 (p.9); Razouls, 1982 (p.430); Dussart & Defaye, 1983 (p.11); Brodsky & al., 1983 (p.141, 146); Sazhina, 1985 (p.115, N); Dussart, 1986 (p.63); Mauchline, 1988 (p.712: pores cuticulaires); Zheng Zhong & al., 1984 (1989) (p.242, Rem.); Razouls, 1993 (p.307); Maly, 1996 (p.727); Madhupratap & al., 1996 (p.863, Table 5: %/copepods); Chihara & Murano, 1997 (p.765); Barthélémy, 1999 a (p.23); Bradford-Grieve & al., 1999 (p.884, 901, 904, 950); Bradford-Grieve,1999 b (p.131, Def.); Ohtsuka & Huys, 2001 (p.461); Boxshall & Halsey, 2004 (p.12, 13; 49; 86: Def.; p.88: Genera Key); Vives & Shmeleva, 2007 (p.464, Genera Key); Blanco-Bercial & al., 2011 (p.103, Table 1, Fig.2, 3, 4, molecular biology, phylogeny) ; Laakmann & al., 2019 (p.330, fig. 2, 3, phylogenetic relationships)
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm
Bradford-Grieve J.M., (2002 onwards). Key to calanoid copepod families. Version 1 : 2 oct 2002. http://www.crustacea.net/crustace/calanoida/index.htm
Rem.:
Cette famille comprend deux genres marins : Centropages, Isias, les autres genres sont saumâtres et dulçaquicoles (cf. in Dussart & Defaye, 1983, p.11 & suiv.; Adamowicz & al., 2007, p.279 & suiv.): Boeckella De Guerne & Richard,1889; Rev. Bayly,1964; Calamoecia Brady,1906; Gippslandia Bayly & Arnott,1969 (cf. in Bradford-Grieve,1999 b (p.140, Déf., Rem.); Gladioferens Henry,1919 (cf. in Dakin & Colefax, 1940, p.89; Bradford-Grieve,1999 b (p.141, Def., Rem.); Hemiboeckella Sars,1912; Karukinka Menu-Marque,2002; Limnocalanus Sars,1863; [Metaboeckella Ekman,1905]; Neoboeckella Bayly,1992; Osphranticum Forbes,1882; Parabroteas Mrazek,1901; [Pseudoboeckella Mrazek,1901 = Boeckella]; Parathalassius Dussart,1986 (cf. in Bradford-Grieve,1999 b (p.147, Déf. Rem.); Sinocalanus Burckhardt, 1913; Cf. in Chihara & Murano, 1997 (p.767, Pl.87); [ Pseudolovenula Marukawa,1921 est une forme juvénile de Megacalanus (Vervoort,1946, p.56)].
Dussart (1986, p.63) note que Bayly & Arnott (1969) ont redéfini la famille des Centropagidae de la façon suivante:
- Urosome de 3 à 4 segments chez la femelle.
- Urosome de 4 à 5 segments chez le mâle.
- A1 de 24 à 25 segments chez la femelle, géniculée à droite chez le mâle.
A2 avec au moins 5 segments à l'exopodite.
- P1 à P4 à exopodite triarticulé; P2 à P4 endopodite au moins biarticulé.
- P5 biramées chez la femelle, à exopodite triarticulé, l'article median prolongé en épine ou en appendice spiniforme à angle distal interne.
- P5 mâle dissymétriques.
Les genres qui composent cette famille sont:
- Centropages dont les exopodites 1 et 2 de P1 sont munis d'une épine externe.
- Isias dont les endopodites de P5 sont uniarticulés et souvent peu apparents chez le mâle.
- Limnocalanus et Sinocalanus caractérisés notamment par la P5 du mâle aux exopodites biarticulés et très dissymétriques.
- Osphranticum dont la P5 droite du mâle a un exopodite inaeticulé orné de 2 soies terminales tandis que la P5 gauche est biarticulée à l'exopodite qui porte 3 soies terminales.
- Boeckella et Hemiboeckella aux endopodites des P1 à P4 triarticulés chez la femelle.
- Calamoecia aux endopodites des P2 à P4 biarticulés, tandis que l'endopodite des P1 est uniarticulé.
- Gladioferens A1 droite des mâles à 18 articles, géniculée entre les articles 15 et 16 et à l'abdomen de 4 segments chez la femelle;
- Gippslandia dont la P5 du mêle est primitive et dissymetrique avec une P5 droite à exopodite biarticulé et endopodite uniarticulé.
- Pseudoboeckella se distingue du genre Boeckella par l'endopodite de la P5 droite du mâle qui est triarticulé et possède des soies.
- Metaboeckella est supprimé par Bayly (1964) et considère la seule espèce de ce genre (M. dilatata) comme une espèce du genre Boeckella. au contraire de Kiefer (1978) qui maitien son statut de genre.
- Parathalassius est un nouveau genre (Dussart, 1986) qui se rapproche des genres Pseudoboeckella et Hemiboeckella mais dont le caractère primitif des P5 semble en faire le genre le plus primitif de tous les Centropagidae connus.
After Madhupratap & al. (1996), la famille des Centropagidae représente de 0,1 à 1,9 % des copepodes selon la saison dans la couche de mélange des eaux océaniques de la région ouest de l'Inde (Mer Arabe), en usant un filet type Multiple Plankton Closing Net à 200 µm de vide de maille (mesh aperture).
Dussart (1986, p.63) note que Bayly & Arnott (1969) ont redéfini la famille des Centropagidae de la façon suivante:
- Urosome de 3 à 4 segments chez la femelle.
- Urosome de 4 à 5 segments chez le mâle.
- A1 de 24 à 25 segments chez la femelle, géniculée à droite chez le mâle.
A2 avec au moins 5 segments à l'exopodite.
- P1 à P4 à exopodite triarticulé; P2 à P4 endopodite au moins biarticulé.
- P5 biramées chez la femelle, à exopodite triarticulé, l'article median prolongé en épine ou en appendice spiniforme à angle distal interne.
- P5 mâle dissymétriques.
Les genres qui composent cette famille sont:
- Centropages dont les exopodites 1 et 2 de P1 sont munis d'une épine externe.
- Isias dont les endopodites de P5 sont uniarticulés et souvent peu apparents chez le mâle.
- Limnocalanus et Sinocalanus caractérisés notamment par la P5 du mâle aux exopodites biarticulés et très dissymétriques.
- Osphranticum dont la P5 droite du mâle a un exopodite inaeticulé orné de 2 soies terminales tandis que la P5 gauche est biarticulée à l'exopodite qui porte 3 soies terminales.
- Boeckella et Hemiboeckella aux endopodites des P1 à P4 triarticulés chez la femelle.
- Calamoecia aux endopodites des P2 à P4 biarticulés, tandis que l'endopodite des P1 est uniarticulé.
- Gladioferens A1 droite des mâles à 18 articles, géniculée entre les articles 15 et 16 et à l'abdomen de 4 segments chez la femelle;
- Gippslandia dont la P5 du mêle est primitive et dissymetrique avec une P5 droite à exopodite biarticulé et endopodite uniarticulé.
- Pseudoboeckella se distingue du genre Boeckella par l'endopodite de la P5 droite du mâle qui est triarticulé et possède des soies.
- Metaboeckella est supprimé par Bayly (1964) et considère la seule espèce de ce genre (M. dilatata) comme une espèce du genre Boeckella. au contraire de Kiefer (1978) qui maitien son statut de genre.
- Parathalassius est un nouveau genre (Dussart, 1986) qui se rapproche des genres Pseudoboeckella et Hemiboeckella mais dont le caractère primitif des P5 semble en faire le genre le plus primitif de tous les Centropagidae connus.
After Madhupratap & al. (1996), la famille des Centropagidae représente de 0,1 à 1,9 % des copepodes selon la saison dans la couche de mélange des eaux océaniques de la région ouest de l'Inde (Mer Arabe), en usant un filet type Multiple Plankton Closing Net à 200 µm de vide de maille (mesh aperture).
Figures
Planche 1
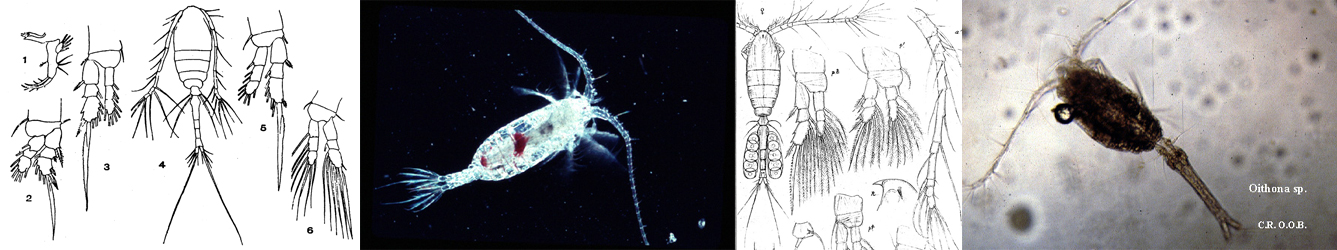
Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.86].
Armature formula of swimming legs P1 to P4, and P5 female (f).
Nota:: Inner seta on basis of P1 located medial to endopod or situated on anterior surface of basis and passing across face of 1st endopodal segment. Setation of both rami reduced in some species and genera; can be variable within species. Rarely P2, 3 and 4 showing asymmetry, as in asymmetrically enlarged coxal setae in P4 of female Gladioferens antarcticus Bayly.
- Female P5 typically with 3-segmented rami; sometimes reduced or modified. Inner margin of 2nd exopodal segment producede into conspicuous spinous process. Endopodal segmentation and setation reduced in some genera.
- Male P5 asymmetrical, coxa and basis distinct; legs typically biramous and both rami primitively 3-segmented, as in Parathalassius. Rami often modified by loss or fusion of segments. Endopod of both legs primitively unmodified, with setation 0-1; 0-1; 2,2,2 but often reduced: sometimes 1-segmented or vestigial, as in Isias. Exopods on both sides often modified by fusion of 2nd and 3rd exopodal segments. Right exopod chelate in some genera, with claw-like 3rd segment opposing spinous process on inner margin of 2nd segment.
- Eggs typically retained in mass on ventral side of urosome or released into water.
Issued from : G.A. Boxshall & S.H. Halsey in An Introduction to Copepod Diversity. The Ray Society, 2004, No 166, Part. I. [p.86].
Armature formula of swimming legs P1 to P4, and P5 female (f).
Nota:: Inner seta on basis of P1 located medial to endopod or situated on anterior surface of basis and passing across face of 1st endopodal segment. Setation of both rami reduced in some species and genera; can be variable within species. Rarely P2, 3 and 4 showing asymmetry, as in asymmetrically enlarged coxal setae in P4 of female Gladioferens antarcticus Bayly.
- Female P5 typically with 3-segmented rami; sometimes reduced or modified. Inner margin of 2nd exopodal segment producede into conspicuous spinous process. Endopodal segmentation and setation reduced in some genera.
- Male P5 asymmetrical, coxa and basis distinct; legs typically biramous and both rami primitively 3-segmented, as in Parathalassius. Rami often modified by loss or fusion of segments. Endopod of both legs primitively unmodified, with setation 0-1; 0-1; 2,2,2 but often reduced: sometimes 1-segmented or vestigial, as in Isias. Exopods on both sides often modified by fusion of 2nd and 3rd exopodal segments. Right exopod chelate in some genera, with claw-like 3rd segment opposing spinous process on inner margin of 2nd segment.
- Eggs typically retained in mass on ventral side of urosome or released into water.
;)
Planche 2
Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.131]. Seta and spine formula of swimming legs P1 to P4.
- P1 - P5 female exopods and endopods 3-segmented; apical spine of exopod strongly serrate; sometimes endopod segments 1 and 2 fused.
- P5 female biramous, natatory, basipods 1 and 2 without setae, exopod segments 1 and 2 with or without 1 outer edge spine, segment 2 with a strong inner spine-like process, exopod segment 3 with or without 2 outer edge spines, 1 terminal serrate spine and 2 or 4 inner edge setae; endopod segments 1 and 2 with 0-1 and 1 inner-edge setae respectively, endopod segment 3 with 4-6 setae.
- P5 male asymmetrical, usually resembling other swimming legs, rami 2 or 3-segmented, right exopod variously transformed into a grasping organ, sometimes endopod atrophied.
Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.131]. Seta and spine formula of swimming legs P1 to P4.
- P1 - P5 female exopods and endopods 3-segmented; apical spine of exopod strongly serrate; sometimes endopod segments 1 and 2 fused.
- P5 female biramous, natatory, basipods 1 and 2 without setae, exopod segments 1 and 2 with or without 1 outer edge spine, segment 2 with a strong inner spine-like process, exopod segment 3 with or without 2 outer edge spines, 1 terminal serrate spine and 2 or 4 inner edge setae; endopod segments 1 and 2 with 0-1 and 1 inner-edge setae respectively, endopod segment 3 with 4-6 setae.
- P5 male asymmetrical, usually resembling other swimming legs, rami 2 or 3-segmented, right exopod variously transformed into a grasping organ, sometimes endopod atrophied.
Genre
Ref.:
Kröyer, 1849 (in Damkaer & Damkaer, 1979, p.43); Brady, 1878 (p.64); Giesbrecht, 1892 (p.59, 303); Giesbrecht & Schmeil, 1898 (p.53, spp.); Wheeler, 1901 (p.172); Sars, 1902 (1903) (p.74); Esterly, 1905 (p.171); van Breemen, 1908 a (p.92); A. Scott, 1909 (p.112); Sewell, 1929 (p.227); Gurney, 1931 a (p.87); Sewell, 1932 (p.227); Wilson, 1932 a (p.85, spp. Key); Rose, 1933 a (p.183, spp. Key); Mori, 1937 (1964) (p.57, spp. Key); Dakin & Colefax, 1940 (p.91, clé spp.); Farran, 1948 (n°11, p.3); Davis, 1949 (p.53); Brodsky, 1950 (1967) (p.315, spp. Key); Kasturirangan, 1963 (p.31, spp. Key); Tanaka, 1963 (p.7); Vervoort, 1964 b (p.308, Rem.: 5 species Groups); Chen & Zhang, 1965 (p.72, spp. Key); Gonzalez & Bowman, 1965 (p.247); Ramirez, 1966 (p.14); Björnberg, 1972 (p.32, Rem. N); Lee, 1972 (p.1, coupling device); Björnberg & al., 1981 (p.644, clé spp.); Razouls, 1982 (p.420); Gardner & Szabo, 1982 (p.351, Rem.); Zheng Zhong & al., 1984 (1989) (p.243, spp. Key); Mauchline, 1988 (p.711); McKinnon & Kimmerer, 1988 (p.171, 175, Rem.); Razouls, 1993 (p.307); Chihara & Murano, 1997 (p.765, clé spp.); Mauchline, 1998 (p.68: F; p.73: M; p.95: F, M); Mulyadi, 1998 (p.54, spp. Key); Bradford-Grieve & al., 1999 (p.951: spp. Key); Bradford-Grieve,1999 b (p.131, Déf., Rem.); Boxshall & Halsey, 2004 (p.90); Mulyadi, 2004 (p.123, spp. Key in Indonesian waters); Vives & Shmeleva, 2007 (p.464, 465: spp. Key); Brylinski & Courcot (2019, Rem.: p.365, coupling device);
Rem.:
type: Centropages typicus . Total: 35 espèces (dont 3 douteuses)
Remarques sur les dimensions et le sex-ratio
Le rapport des longueurs (M/F) est de 0,99 (n = 33; SD = 0,1906). Le sex-ratio (F/M) est de 1,03.
Figures
Planche 0
Vervoort (1964) divides Centropages species into 5 species groups: "typicus", "furcatus", "hamatus", "orsinii", "violaceus" according to the shape of the last thoracic somite, shape of abdomen, morphology of P5 and the male A1; subsequently, Ohtsuka & al. (2003, 2005) recognized 2 other groups: "trispinosus" and "alcocki".
Prusova & al. (2019, p.526) underline that Vervoort (1964) pointed out the genus Centropages to be revised, and arranged species known at this time provisionnaly into 5 species-Groups: 'typicus, furcatus, hamatus, orsinii, violaceus', with C. ponticus referred to the 'hamatus' Group, with C. abdominalis, C. alcocki, C. hamatus, C. kroyeri, C. tenuiremis, C. trispinosus. Subsequently, Ohtsuka & al. (2003, 2005) recognized 2 other Groups: 'trispinosus' and 'alcocki', thereby removing 2 corresponding species from the Vervoort's 'hamatus'-Group. Thus, to date the 'hamatus' Group includes 5 species.
Vervoort (1964) divides Centropages species into 5 species groups: "typicus", "furcatus", "hamatus", "orsinii", "violaceus" according to the shape of the last thoracic somite, shape of abdomen, morphology of P5 and the male A1; subsequently, Ohtsuka & al. (2003, 2005) recognized 2 other groups: "trispinosus" and "alcocki".
Prusova & al. (2019, p.526) underline that Vervoort (1964) pointed out the genus Centropages to be revised, and arranged species known at this time provisionnaly into 5 species-Groups: 'typicus, furcatus, hamatus, orsinii, violaceus', with C. ponticus referred to the 'hamatus' Group, with C. abdominalis, C. alcocki, C. hamatus, C. kroyeri, C. tenuiremis, C. trispinosus. Subsequently, Ohtsuka & al. (2003, 2005) recognized 2 other Groups: 'trispinosus' and 'alcocki', thereby removing 2 corresponding species from the Vervoort's 'hamatus'-Group. Thus, to date the 'hamatus' Group includes 5 species.
Genre
Ref.:
Bayly & Arnott, 1969 (p.193)
Rem.:
type: Gippslandia estuarina. Total: 1 sp.
Remarques sur les dimensions et le sex-ratio
In brackish water in the Gippsland lakes of Victoria (SE Australia).
Figures
;)
Planche 1
Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.141, Fig.99]. Gippslandia estuarina from Bayly & Arnott (1969).
Female: A, habitus (lateral view); B, P5.
Male: C, habitus (lateral view); D, P5.
Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999. [p.141, Fig.99]. Gippslandia estuarina from Bayly & Arnott (1969).
Female: A, habitus (lateral view); B, P5.
Male: C, habitus (lateral view); D, P5.
Genre
Ref.:
Brady, 1878 (p.62); Giesbrecht, 1892 (p.60, 323); Giesbrecht & Schmeil, 1898 (p.62); Sars, 1902 (1903) (p.78); van Breemen, 1908 a (p.96); Sewell, 1932 (p.232); Rose, 1933 a (p.189); Pillai P., 1975 a (p.325, species Key); Razouls, 1982 (p.431); Dussart & Defaye, 1983 (p.12); Razouls, 1993 (p.307); Mauchline, 1998 (p.70: F; p.73: M); Bradford-Grieve,1999 b (p.145, Def.); Boxshall & Halsey, 2004 (p.88); Vives & Shmeleva, 2007 (p.482)
Rem.:
Isias clavipes Boeck, 1865. Total: 4 spp.
Remarques sur les dimensions et le sex-ratio
La moyenne des longueurs des femelles est de 1,413 mm (n= 3; S= 0,142; Cv= 0,10) et de 1,373 mm pour les mâles (n= 3; S= 0,172; Cv= 0,125). Le rapport des longueurs (M/F) est de 0,972 ou 97,2 % (n= 3; S= 0,070; Cv= 0,072)
Figures
;)
Planche 1
Issued from : M. Rose in Faune de France, N° 26, 1933. [p.189, Fig.220]. Isia clavipes. after G.O. Sars.
Issued from : M. Rose in Faune de France, N° 26, 1933. [p.189, Fig.220]. Isia clavipes. after G.O. Sars.
Genre
Ref.:
Dussart, 1986 (p.67, Def.); Bradford-Grieve, 1999b (p.147, Def.)
Rem.:
Type: Parathalassius fagesi Dussart, 1986. Total: 1 sp.
Figures
;)
Planche 1
Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999/ [p.147]. After Dussart, 1986.
Female (from mouth of La Foa River, New-Caledonia): A, habitus (dorsal); B, exopod of P5.
Male: C, habitus (dorsal); D, P5.
Issued from : J.M. Bradford-Grieve in NIWA Biodiversity Memoir 111, 1999/ [p.147]. After Dussart, 1986.
Female (from mouth of La Foa River, New-Caledonia): A, habitus (dorsal); B, exopod of P5.
Male: C, habitus (dorsal); D, P5.
Genre
Ref.:
Orsi & al., 1983 (p.358)
Rem.:
Dulçaquicole and in estuaries.
Total: 3 spp.
Total: 3 spp.
Remarques sur les dimensions et le sex-ratio
The mean female size is 1.565 mm (n= 6; SD = 0.2827), and the mean male size is 1.458 mm (n = 6; SD = 0.3122). The size ratio (male: female) is 0.937 (n = 3; SD = 0.066). The sex ratio (female:male) is 1.
Informations complémentaires pour la famille des Centropagidae ( retour aux informations principales )
)Remarques sur les tailles des adultes :
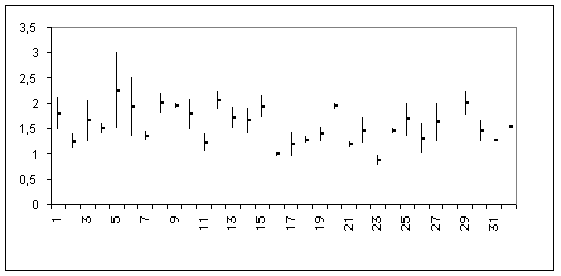
Les valeurs minimales et maximales des longueurs des femelles adultes sont représentées sur la figure 1 pour Centropages (1 à 29) et Isias (30 à 32).
Les espèces des genres Centropages et Isias se répartissent en divers groupes (Tableau I) dont les tailles extrèmes vont de 0,66 mm à 3 mm.
Tableau I: Centropages et Isias : Longueur totale et localisation (E: épipélagique; L: littoral; B: bathypélagique; N: néritique, oc: océanique)
Deux espèces présentent de fortes variations, comprises entre 1,3 et 3 mm, et correspondent à des formes cosmopolites d'eaux froides sub-antarctiques et tempérées froides, ou sub-tropicales plus ou moins profondes.
Onze espèces sont comprises entre 1,5 et 2,1 mm (soit 37,93 p. cent) et sont généralement cosmopolites tropicales, sub-tropicales et / ou dans des eaux plus froides.
Quinze espèces comprises entre ± 1 et 1,5 mm habitent pour la plupart des eaux tropicales eu tempérées chaudes. Une espèce ( C. ponticus ), du moins si elle ne correspond pas à une variété de C. kröyeri , montre les dimensions les plus faibles chez les femelles et les mâles (≤ 1 mm)
Les valeurs minimales et maximales des longueurs des femelles adultes sont représentées sur la figure 1 pour Centropages (1 à 29) et Isias (30 à 32).
figure 1
Les espèces des genres Centropages et Isias se répartissent en divers groupes (Tableau I) dont les tailles extrèmes vont de 0,66 mm à 3 mm.
Tableau I: Centropages et Isias : Longueur totale et localisation (E: épipélagique; L: littoral; B: bathypélagique; N: néritique, oc: océanique)
ESPÈCES |
LG (mm) F, M |
Localisation |
Prof |
C. brachiatus |
1,3 à 2,5-3 |
Cosmop. Eaux froides, sub-Ant, sub-Trop |
E |
C. bradyi |
- |
Cosmop. sub-Ant, Tempéré froid |
E - B |
C. abdominalis |
1,5 à 2,10 |
Tempéré chaud, sub-Arct, Arct ± saumâtre |
L |
C. aucklandicus |
- |
Tempéré froid, sub-Antarct ? |
L |
C. calaninus |
- |
Cosmop. Tropic, Tempéré froid |
E |
C. caribbeanensis |
- |
(sub)- Tropic |
|
C. chierchiae |
- |
sub- Tropic |
E |
C. elegans |
- |
Tropic |
|
C. gracilis |
- |
Tropic |
E |
C. longicornis |
- |
Tropic, sub-Tropic |
E |
C. tenuiremis |
- |
Tropic, eaux froides Pacif NW |
N |
C. typicus |
- |
Tempéré froid, sub-Tropic |
E, N, B |
C. violaceus |
- |
Cosmop Tropic & sub-Tropic |
E, oc |
C. alcocki |
± 1 à ± 1,5 |
Tropic, ± saumâtre |
L |
C. australiensis |
- |
Tempéré froid |
N |
C. brevifurcus |
- |
Tropic ± saumâtre |
|
C. dorsispinatus |
- |
Tropic |
L |
C. halinus |
- |
sub-Tropic, hyperhalin |
L |
C. hamatus |
- |
Tempéré froid |
|
C. karachiensis |
- |
Tropic |
L - E |
C. kröyeri |
- |
Tropic, Tempéré chaud |
E |
C. natalensis |
- |
sub-Tropic |
|
C. orsinii |
- |
Tropic |
E |
C. sinensis |
- |
Tropic |
|
C. trispinosus |
± 1 à ± 1,5 |
Tropic |
L |
I. clavipes |
- |
sub-Tropic, Tempéré froid |
N |
I. tropica |
- |
Tropic ± saumâtre |
L |
I. uncipes |
- |
Tropic, euryhalin |
L |
C. ponticus ± 1 |
± 1 |
Tempéré chaud |
L |
Deux espèces présentent de fortes variations, comprises entre 1,3 et 3 mm, et correspondent à des formes cosmopolites d'eaux froides sub-antarctiques et tempérées froides, ou sub-tropicales plus ou moins profondes.
Onze espèces sont comprises entre 1,5 et 2,1 mm (soit 37,93 p. cent) et sont généralement cosmopolites tropicales, sub-tropicales et / ou dans des eaux plus froides.
Quinze espèces comprises entre ± 1 et 1,5 mm habitent pour la plupart des eaux tropicales eu tempérées chaudes. Une espèce ( C. ponticus ), du moins si elle ne correspond pas à une variété de C. kröyeri , montre les dimensions les plus faibles chez les femelles et les mâles (≤ 1 mm)