|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Metridinidae ( Famille ) |
|
|
|
Pleuromamma ( Genre ) |
|
|
| |
Pleuromamma robusta (F. Dahl, 1893) (F,M) | |
| | | | | | | Syn.: | Pleuromma robustum F. Dahl, 1893;
no Pleuromamma robusta antarctica Steuer, 1931 (p.8, fig.F); 1932 a (p.24, figs.F,M); 1933 (p.13, figs.F); Séret, 1979 (p.139, figs.F,M); Razouls, 1994 (p.146, fig.F, Rem.);
Pleuromamma robusta typica Steuer, 1931 (p.7); 1932 a (p.24, figs.F,M); 1933 (p.12, figs.F,M);
Pleuromamma robusta robusta : Bainbridge, 1972 (p.61, Appendix Table III: occurrence);
? Pleuromamma robusta : Mazzocchi & al. 1995 (p.57, figs.F,M, Rem.); Pleuromamma sp. Razouls, 1994 (p.146, fig.F, Rem.). | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.110); Sars, 1902 (1903) (p.115, figs.F,M); Farran, 1908 b (p.61); Pesta, 1920 (p.528); Farran, 1929 (p.260); Lysholm & Nordgaard, 1921 (p.24); Sars, 1925 (p.205); Farran, 1926 (p.272); 1929 (p.209, 260); Rose, 1929 (p.30); Steuer, 1931 (p.7); 1932 a (p.20, 24, 75); Wilson, 1932 a (p.126, figs.F,M); Steuer, 1933 (p.11, figs.F,M); Rose, 1933 a (p.183, figs.F,M); Jespersen, 1934 (p.102, Rem.); Mori, 1937 (1964) (p.70, figs.F); Jespersen, 1940 (p.47, fig.5); Wilson, 1942 a (p.202, fig.F); Lysholm & al., 1945 (p.32); Farran, 1948 f (n°17, p.3, figs.F,M); Marques, 1953 (p.115, figs.F,M); Vervoort, 1957 (p.125, Rem.); Marques, 1959 (p.214); Chen & Zhang, 1965 (p.69, figs.F,M); Vervoort, 1965 (p.107, Rem. as P. robusta robusta); Fleminger, 1967 a (p.XII, Rem.); Ramirez, 1969 (p.73, figs.F,M, Rem.); Shih & al., 1971 (p.42, 205); Kos, 1972 (Vol.I, figs. F, M, Rem.; Bradford, 1972 (p.44, figs.F,M); Shih & Laubitz, 1978 (p.49); Björnberg & al., 1981 (p.642, figs.F,M); Gardner & Szabo, 1982 (p.340, figs.F,M); Zheng & al., 1982 (p.50, figs.F,M); Ferrari, 1984 a (p.168, 170); Roe, 1984 (p.358); Kim & al., 1993 (p.270); J.S. Park & Mauchline, 1994 (p.408 & suiv., fig.F); J.S. Park, 1995 (p.212, figs.); Ferrari & Saltzman, 1998 (p.215, figs.F,M, Rem.); Bradford-Grieve & al., 1999 (p.884, 949, figs.F,M); Bradford-Grieve,1999 b (p.124, figs.F,M, Rem., figs.179, 192); Conway & al., 2003 (p.95, figs.F,M, Rem.); G. Harding, 2004 (p.30, figs.F,M); Mulyadi, 2004 (p.64, figs.F,M, Rem.); Vives & Shmeleva, 2007 (p.382, figs.F,M, Rem.) | 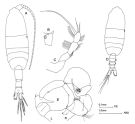 isued from : Bradford-Grieve J.M. in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.125, Fig.85]. Female (41°45'S, 174°55'E): A, habitus (dorsal); B, genital somite (left lateral side); C, P5. Male: D, habitus (dorsal); E, P5 (L = left leg; R = right leg). Female characteristics: Genital swelling on anterior part of genital segment. - A1 denticles small. - Endopod segment 1 of P2 with a notch and hooks on both sides. - P5 4-segmented with 3 apical setae. Male characteristics: Urosome almost symmetrical. - Geniculate A1 with proximal segments with degenerate teeth, segment 17 without a tooth, segment 18 has a rasp with small, oblique teeth. - Endopod segment 1 of P2 with a notch and hooks on both sides. Nota: The Southwest Pacific specimens have the inner extensions on male P5 left exopod segment 1 and right exopod segment 2 longer than those figured by Steuer (1932).
|
 issued from : G.O. Sars in An Account of the Crustacea of Norway. Vol. IV. Copepoda Calanoida. Published by the Bergen Museum, 1903. [Pl.LXXVIII]. Female & Male. Nota: lum.org = luminous organ; Sens.ap = aesthetasc on A1 male
|
 issued from : F.D. Ferrari & J. Saltzman in Plankton Biol. Ecol., 1998, 45 (2). [p.216, Fig.7]. Female (from eastern tropical Pacific): A, genital complex (ventral); B, idem (lateral right side); C, dark organ on right side of thoracic segment 2; D, A1 segments 1-2 ( arrows to segmental attenuations); E, P5. Male: F, rostral area (lateral); G, urosome (ventral); H, urosomal segments 2-3 (dorsal); I, anal segment and caudal rami ( dorsal) ( arrow to set of spinules to left of anal flap); J, left A1 segments 13-14 ( arrowhead to tip of distal rigid seta, arrow to axial angle of segment attenuation); K, P5 (anterior) ( open arrow connects left coxa and basis; arthropodial membranes and poorly sclerotized areas stippled, curved arrow to angled proximal face of attenuation of left exopod 3); straight arrow to twisted attenuation of right exopod 2); L, P5 (posterior) (arthrodial membranes and poorly sclerotized areas stippled).
Nota: Thoracic, abdominal somites and ramal segments are numbered according to their appearance during development as interpreted from data of Hulsemann (1991).
|
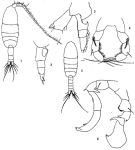 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.23, 1-6]. Female (from E China Sea): 1, habitus (dorsal); 2, urosome (lateral right side); 3, endopod of left P2 (posterior); 4, P5 (posterior). Male: 5, habitus (dorsal); 6, P5 (posterior).
|
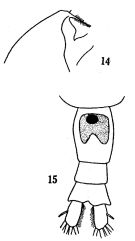 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.22, 14-15]. Female: 14, forehead (lateral); 15, urosome (ventral).
|
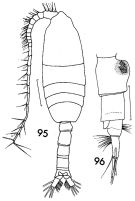 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.70, Lam. XIII, figs.95, 96]. Female (from off Mar del Plata): 96, urosome (lateral left side). Male: 95, habitus (dorsal). Scale bars in mm: 0.5 (95); 0.4 (96).
|
 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.50, Fig.28-1]. a, habitus (dorsal); b-c, urosomelateral and ventral, respectively); d, forehead (lateral); e, proximal segments of A1; f, P1; g, basipodite segments 1 and 2, exopodal segment 1 and endopod of P1; h, P2; i, basipodal segment 2, exopodal segments 1 and 2, endopod of P2; j, P3; k, P4; l, P5. Scale bar in mm.
|
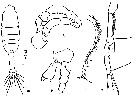 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.51, Fig.28-2]. Male: m, habitus (dorsal); n, right A1; o, grasping segments of right A1; p, basipodal segment 2, exopodal segment 1 and endopod of P2; q, P5. Scale bar in mm.
|
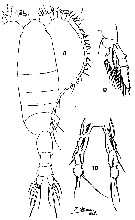 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (2nd edit., 1964). [Pl.35, Figs.8-10]. Female: 8, habitus (dorsal); 9, P2: 10, P5.
|
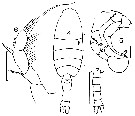 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.45, Fig.11, (4-7]. Female (from Kaikoura, New Zealand): 4, habitus (dorsal); 6, P5. Male: 5, P5; 7, urosome (dorsal). Scale bars: 1 mm (4, 7); 0.1 mm (5, 6). Nota: Most specimens were of the scanty haired forma antarctica (Steuer, 1932) but some forma Typica were present.
|
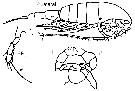 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.30]. Female & Male. L = left leg; R = right leg.
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.65, Fig.36]. Female (from Flores Sea): a, habitus (dorsal); b, urosome (lateral left side); c, urosome (ventral); d, P2. Male: e, habitus (dorsal); f, P5.
|
 Issued from : C. Razouls in Ann. Inst. océanogr., Paris, 1994, 70 (1). [p.140]. Caractéristiques morphologiques de Pleuromamma robusta antarctica femelle et mâle adultes. Terminologie et abbréviations: voir à Calanus propinquus.
|
 Issued from : C. Razouls in Ann. Inst. océanogr., Paris, 1994, 70 (1). [p.148]. Comparison of varieties robusta and antarctica.
1: anal segment and caudal rami from robusta robusta; 2: same, from robusta antarctica; 3, P5 female from Pleuromamma sp. (Kerguelen Islands).
Un individu femelle récolté à la station ''Kerfix'' (Ïles Kerguelen) présente des caractères qui ne permettent pas de l'attribuer en toute certitude à P. robusta antarctica
- La longueur totale est de 4,48 mm.
- Pr/Ur = 1,83.
- A1 comme in Steuer (1933, fig.56).
- La tête et le Th1 présentent une ligne de suture fine et complète.
- La P5 est tout à fait comparable à celle figurée in Steuer (1933, p.5, fig.13 ou p.15, fig.65), et non comme celle de P. robusta in Steuer (1933, p.12, fig.51) ou in Vervoort (1951, p.126, fig.69b).
- Les proportions de l'urosome sont en %: 32,9/28,7/20,4/F = 17,9.
- Le segment génital montre une spermathèque en position très antérieure comme chez P. robusta.
- L'opercule anal est lisse.
- Les rames caudales (Furca) ne montrent aucune trace de poils sur les bord internes; Lg/lg = 2,0.
- Par comparaison avec P. robusta antarctica la P5 montre qu'à l'apex du segment distal la soie la plus externe est la moitié de la suivante, elle-même un peu moins de la moitié de la plus longue.
Un second individu femelle examiné présente une longueur totale de 4,22 mm. Pr/Ur = 2,08; les rames caudales présentent des soies longues sur les bords internes; l'opercule anal est lisse; la P5 comme chez P. quadrungulata, mais les A1 totalement sans dents ou pointes.
|
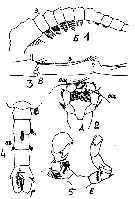 Issued from : M.C. Kos in Field guide for plankton. Zool Institute USSR Acad., Vol. I, 1972. After Steuer, 1932. Female: 1, A1 (proximal segments); 2,P5. Male: 3, A1 (grasping segments); 4, abdomen (dorsal); 5, P5/
| | | | | Ref. compl.: | | | Cleve, 1904 a (p.195); Pearson, 1906 (p.24); Hardy & Gunther, 1935 (1936) (p.167, Rem.); Jespersen, 1939 (p.68, Rem.); Sewell, 1948 (p.349, 503, 508, 514, 519, 547, 549, 557, 567); C.B. Wilson, 1950 (p.289); Baker, 1954 (p.203, 211, fig.5); Østvedt, 1955 (p.15: Table 3, p.75); ? King & Hida, 1955 (p.11); Chiba & al., 1957 (p.308); 1957 a (p.12); Yamazi, 1958 (p.150, Rem.); Conover, 1960 (p.399, Table I, respiratory rate); Fagetti, 1962 (p.28); Marshall & Orr, 1962 (tab.3); Grice & Hart, 1962 (p.287, table 3); Grice, 1963 a (p.496); Bary, 1963 a (p.1519, Table 1); 1964 (p.183, T-S diagram-occurrences); Unterüberbacher, 1964 (p.27); De Decker, 1964 (p.16, 24, 27); Grice & Hulsemann, 1965 (p.224); Mazza, 1966 (p.71); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio/Kuroshio transitional area, Table 8); Ehrhardt, 1967 (p.739, 887, figs.51-53, geographic distribution, Rem.); Matthews, 1967 (p.159, Table 1, Rem.); Fleminger, 1967 a (tabl.1); Deevey & Brooks, 1977 (tab.2); Delalo, 1968 (p.138); Vinogradov, 1968 (1970) (p.68, 268); De Decker, 1968 (p.45); Macdonald & al., 1972 (p.213, hydrostatic pressure effect); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 b (p.548, Rem.); Björnberg, 1973 (p.338, 388); Harding, 1974 (p.141, tab.2, gut contents); Corral Estrada & Pereiro Muñoz, 1974 (tab.I); Vives & al., 1975 (p.46, tab.II, III, IV, XII); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Carter, 1977 (1978) (p.36); Arashkevich, 1978 (p.118, Table: diets); Colebrook, 1978 (tab.1); Longhurst & Williams, 1979 (p.1, Table IVb, V, fig.5, 4b, 8a, vertical distribution/North Atlantic Drift); Pipe & Coombs, 1980 (p.223, figs. 1, 2, table 1, vertical distribution); Vaissière & Séguin, 1980 (p.23, tab.2); Vives, 1982 (p.293); Kovalev & Shmeleva, 1982 (p.84); Sameoto, 1984 (p.767, vertical migration); Guangshan & Honglin, 1984 (p.118, tab.); Tremblay & Anderson, 1984 (p.5); De Decker, 1984 (p.316); Hopkins, 1985 (p.197, Table 1, gut contents); Bamstedt & Tande, 1985 (p.259, Table 2: literature data respiration & excretion); Brenning, 1985 a (p.28, Table 2); 1986 (p.9, spatial distribution, T-S diagram, Rem.); Dearborn & al., 1986 (p.1, predation by benthic star); Lozano Soldevilla & al., 1988 (p.59); Ward, 1989 (tab.2); Ferrari & Dearborn, 1989 (p.1315, predator); Cervantes-Duarte & Hernandez-Trujillo, 1989 (tab.3); Pancucci-Papadopoulou & al., 1990 (p.199); Atkinson & al., 1990 (tab.1); Fransz & al., 1991 (p.9); Yoo, 1991 (tab.1); Baessa De Aguiar, 1991 (1993) (p.107); Hernandez-Trujillo, 1991 (1993) (tab.I); Morales C.E. & al., 1991 (p.455, Table I, grazing); Gowing & Wishner, 1992 (tab.1); Ayukai & Hattori, 1992 (p.163, Table 5, fecal pellet production rate); Siegel & al., 1992 (p.18, tab.3); Morales C.E. & al., 1993 (p.185); Hays & al., 1994 (tab.1); Shih & Young, 1995 (p.71); Ward & al., 1995 (p.195, Table 2); Atkinson & al., 1996 (p.195, feeding v.s. phytoplankton bloom); Atkinson & al., 1996 (p.1387, diel periodicity, feeding); Park & Choi, 1997 (Appendix); Falkenhaug & al., 1997 (p.449, spatio-temporal pattern); Hure & Krsinic, 1998 (p.61, 102); Mauchline, 1998 (tab.30, 64); Noda & al., 1998 (p.55, Table 3, occurrence); Reid & Hunt, 1998 (p.310, figs.2, 3, Rem.); Gilabert & Moreno, 1998 (tab.1, 2); Halvorsen & Tande, 1999 (p.279, tab.2, 3, Rem.: p.282); Tirelli & Mayzaud, 1999 (p.197, gut evacuation); Lapernat, 2000 (tabl.3, 4) ; Atkinson & Sinclair, 2000 (p.46, 50, 51, 54, 55, zonal distribution); Razouls & al., 2000 (p.343, Appendix); d'Elbée, 2001 (tabl. 1); Holmes, 2001 (p.20); Li & al., 2001 (p.894, tab.1); Sameoto & al., 2002 (p.13); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, figs.5, 6); Schmidt K. & al., 2003 (p.277, stable isotopes 15N, 13C); Hsiao & al., 2004 (p.326, tab.1); CPR, 2004 (p.60, fig.179); Lo & al., 2004 (p.89, tab.1); Berasategui & al., 2005 (p.313, fig.2); Zuo & al., 2006 (p.159, tab.1, abundance, fig.8: statios group); Schultes & al., 2006 (p.21, fig.1); Hop & al., 2006 (p.182, Table 4); Dur & al., 2007 (p.197, Table IV); Blachowiak-Samolyk & al., 2007 (p.2716, Table 2); Jitlang & al., 2008 (p.65, Table 1); Cabal & al., 2008 (289, Table 1); Ward & al., 2008 (p.241, Appendix II ); Morales-Ramirez & Suarez-Morales, 2008 (p.521); Lopez Ibarra, 2008 (p.1, Table 1, 2: Rem.); Fernandes, 2008 (p.465, Tabl.2); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); C.-Y. Lee & al., 2009 (p.151, Tab.2); Galbraith, 2009 (pers. comm.); Kruse & al., 2009 (p.301, gut content); Tseng & al., 2009 (p.327, fig.5, feeding); Lan Y.-C. & al., 2009 (p.1, Table 2, % vs hydrogaphic conditions); Takahashi & al., 2010 (p.317, Table 3, 4); Takahashi & al., 2010 (p.235, Table 2); Mazzocchi & Di Capua, 2010 (p.426); W.-B. Chang & al., 2010 (p.735, Table 2, abundance); Dvoretsky & Dvoretsky, 2010 (p.991, Table 2); Pepin & al., 2011 (p.273, Table 2, seasonal abundance); Kosobokova & al., 2011 (p.29, Table 2, Rem.: Nansen Basin); Hsiao S.H. & al., 2011 (p.475, Appendix I); Kâ & Hwang, 2011 (p.155, Table 3: occurrence %); Takahashi & al., 2011 (p.134, distribution); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Salah S. & al., 2012 (p.155, Tableau 1); Zizah & al., 2012 (p.79, Tableau I, Rem.: p.86); Alvarez-Fernandez & al., 2012 (p.21, Rem.: Table 1); Teuber & al., 2013 (p.1, Table 1, 2, 3, fig.5, abundance vs oxygen minimum zone, respiration rates, enzyme activity); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Tseng & al., 2013 (p.507, seasonal abundance); Bode M. & al., 2013 (p.1, Table1, 3, respiration rate & ETS activity); Schukat & al., 2013 (p.1, Table 1, 2, fig.2, respiration, ingestion); Lidvanov & al., 2013 (p.290, Table 2, % composition); Ward & al., 2014 (p.305, Table 6, 7, seasonal and abundance in the ''Discovery'' Investigations in the 1930s); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Lopez-Ibarra & al., 2014 (p.453, fig.6, 8, Table 2, biogeographical affinity); Fierro Gonzalvez, 2014 (p.1, Tab. 3, 5, occurrence, abundance); Zaafa & al., 2014 (p.67, Table I, occurrence); Zakaria & al., 2016 (p.1, Table 1); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Belmonte, 2018 (p.273, Table I: Italian zones); Bode & al., 2018 (p.840, Table 1, respiration & ingestion rates, depth); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions). | | | | NZ: | 23 | | |
|
Carte de distribution de Pleuromamma robusta par zones géographiques
|
| | | | | | | | | | | | | | | | | | 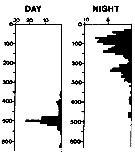 issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.23, Fig.8a]. issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.23, Fig.8a].
Day/night percentage numerical profiles at India Station (59°N, 19°W) at end of March 1975.
Depth in meter.
Sampling by LHPRs (Longhurst-Hardy Plankton Recorders). |
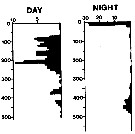 issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.24, Fig.8b]. issued from : A. Longhurst & R. Williams in J. Plankton Res., 1979, 1 (1). [p.24, Fig.8b].
Day/night percentage numerical profiles at India Station (59°N, 19°W) in early May 1975.
Depth in meter. |
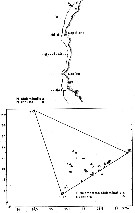 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.8, Figs.5, 6]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.8, Figs.5, 6].
Spatial distribution and T-S Diagram for Pleuromamma robusta and P. abdominalis from 8° S - 26° N; 16°- 20° W.
SO: Southern Surface Water (S °/oo: 34,50; T°C: 29,0); ND: Northern Water of the Surface Layer (S °/oo: 37,5; T°C: 21,0); SD: Southern Deep Water of the surface layer (S °/oo: 35,33; T°C: 13,4). See commentary in Temora stylifera and Brenning (1985 a, p.6). |
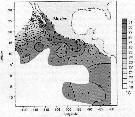
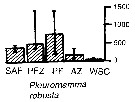 Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3] Issued from : A. Atkinson & J.D. Sinclair in Polar Biol., 2000, 23. [p.50, Fig.3]
Pleuromamma robusta from Scotia Sea.
Median and interquartile ranges of copepods (nos /m2) in the five water zones; from north to south these are SAF Subantractic Front area, PFZ Polar frontal Zone, PF Polar Front area, AZ Antarctic Zone, WSC Weddell-Scotia Confluence area/ East Wind Drift.
Numbers on the plots are upper interquartiles where these could not be scaled. |
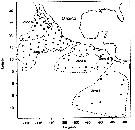
 issued from A. de C. Baker in 'Discovery' Rep., 1954, 27. [p.215, Fig.5]. issued from A. de C. Baker in 'Discovery' Rep., 1954, 27. [p.215, Fig.5].
Occurrence of Pleuromamma robusta in all longitudes around Antarctic zone of the Southern Ocean.
The percentage frequency of occurrence in samples taken within every 20° of longitude.
Nota: During the 'Discovery' investigations some thousands of plankton samples have been taken from stations spread over the whole of the Southern Ocean at all seasons of the year, the majority south of the Antarctic Convergence. An arbitrary selection of samples has been made from hauls between the surface and a depth of 250 m, which means that they have been taken from within the limits of the Antarctic surface water. |
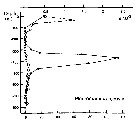 Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.203, Fig.4, B]. Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.203, Fig.4, B].
Abundance (10/m3) and biomass (g dry mass/m3) profiles at the oceanic station from the shelf break in water 4000 m deep off Bird Island, South Georgia (53°04'S, 39°51'W) in January 1990.
Values on the horizontal axes at the base of each profile represent abundance and the one above biomass. Solid line = midday haul; hatched line = midnight haul. |
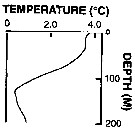 Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.198, Fig.1, B (modified C.R.)]. Issued from : P. Ward, A. Atkinson, A.W.A. Murray, A.G. Wood, R. Williams & S. Poulet in Polar Biol., 1995, 15. [p.198, Fig.1, B (modified C.R.)].
Profile temperature-depth at the oceanic stations from the shelf break in water 4000 m deep off Bird Island, South Georgia, in January 1990. |
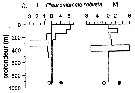 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.9 h]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.9 h].
Verical distribution of Pleuromamma robusta at an eutrophic site (off Mauritanian coast: 20°32 'N, 18°36' W) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
 Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.459, Fig.8]. Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.459, Fig.8].
Abundance recorded in the geographic zones with the highest canonical coefficient on the first two axes of the discriminant analysis.
See chart of geographic zones (Fig.1, p.454) |
 Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.454, Fig.1]. Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.454, Fig.1].
Study area and samplingb stations in the Eastern Tropical Pacific Ocean.
The six geographic zones identified in the analysis: Western coast of the Baja California peninsula (a), Mexicvo's coastal region (b), Mexsico's oceanic region (c), central coastal region (d), central oceanic region (e), equatorial oceanic region (f).
Ninety-six zooplankton samples were collected with a Bongo net (333 Mm mesh size) towed obliquely from 200 m. An average of 438 m3 of water was filtered by tow. Sampling began in July near the Baja California and continued toward equator until December 2003.
The sampling area comprises a large area in the Eastern tropical Pacific Ocean (ETPO) located between the subtropical gyres of the North and South pacific in tropical Surface Water (TSW); this water is characterized by surface temperature > 25°C and Salinity < 34. Sampling locations encompassed parts of the eastern Pacific warm pool, the North Equatorial Countercurrent, the North Equatorial Current, and the southern terminus of the California Current. The area is also influenced by the California Current (cold waters) and the Costa Rican Coastal Current (warm waters. In this sense, physical and chemical characteristics of the sea influence the community structure dynamics of pelagic copepods. |
 Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.455, Fig.2]. Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.455, Fig.2].
Seawater temperature at a depth of 5 m in the study area. The dots represent the sampling stations.
See Fig.1 for the geographic zones. |
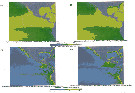 Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.456, Fig.3]. Issued from : G.A. Lopez-Ibarra, S. Hernandez-Trijillo, A. Bode & M.J. Zetina-Rejon in Cah. Biol. Mar., 2014, 55. [p.456, Fig.3].
Satellite images for the average sea surface temperature (SST) and Chlorophyll-a concentration (Chl a) in the Eastern Tropical Pacific Ocean. A: SST Summer/2003. B: SST Autumn/2003. C: Chl a Summer/2003. D: Chl a Autumn/2003.
See Fig.1 for the geographic zones. |
 Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1). Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1).
Cf. explanations of these measures in Calanoides natalis from the same authors.
Compare with Pleuromamma abdominalis and P. xiphias. |
 Issued from : M. Bode, A. Schukat, W. Hagen & H. Auel in J. Exp. Mar. Biol. Ecol., 2013, 444. [p.3, Table 1]. Issued from : M. Bode, A. Schukat, W. Hagen & H. Auel in J. Exp. Mar. Biol. Ecol., 2013, 444. [p.3, Table 1].
Dry mass, individual respiration rate and ETS activity from the northern Benguela Current upwelling system along transects at 23°S and 26°40'S, off Walvisbay and Lüderitz (Namibia).
Relationship between individual ETS activities and individual respiration rates; mean ETS activities with standard deviations are plotted again mean respiration rates (Cf. in Calanoides natalis. The letter besides each point identifies the associated species in Table 1 (h, for this species) in the same authors.
For Conover & Corner (1968) the respiration rate at 3-5°C were 0.93 µL O2 h-1 ind-1 (DM: 362µg), in the North Atlantic; For Corner (1960) at 8°C: 2.26-2.80 (283 µg) in Western North Atlantic. |
 Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.48, Fig. 22 and p.51, Fig.24] Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.48, Fig. 22 and p.51, Fig.24]
Spatial distribution of the isotopic values from the dominant species for localities (see map for the different zones: a to f). A: 13C; B: 15N.
Nota: Sampling in Julio to December 2003, with a Bongo net (333 µm mesh aperture) from obliquely drawing from 200 m depth to surface.
Cf. Map of the 6 zones sampled, legend and remarks by the same author in Subeucalanus subcrassus. |
 Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.37, Table 3]. Issued from : G.A. Lopez Ibarra in Tesis, Inst. Politec. Nac., CICIMAR, 2008. [p.37, Table 3].
Summary of abundance and steady isotopics 15N and 13C in Subeucalanus subcrassus.
C = carnivore; T = tropical biogeographical affinity; zones (a-f) = Cf. fig.1 in the same author.
Compare with Subeucalanus subcrassus and other species (Tabla 3).
Note the differences with pleuromamma abdominalis and P. gracilis; P; robusta seems in this region more strictly carnivorous.
From Schmidt & al. (2003), In Antarctica (Lazarev Sea): 15N = 4.5 and 13C = -24.5. |
| | | | Loc: | | | Cosmopolite: Antarct. (Peninsula, Drake Passage, Scotia Sea, Weddell Sea, SW & SE Atlant., Indian, SW Pacif.), sub-Antarct. (SW Atlant., off South Georgia, Kerguelen Is., SW & SE Pacif.), Indian (rare), Bay of Bengal, Madagascar, Natal, South Africa (E & W), Namibia, Angola, G. of Guinea, off Lagos, Brazil-Argentina (offshore, shelf), off S Cape Verde Is., off Morocco-Mauritania, Cap Ghir (Morocco), Canary Is., off Madeira, off W Cabo Finisterre, Bay of Biscay, Newfoundland, off SW Ireland, Porcupine Bank, Norway (Raunefjorden), Barents Sea, Ibero-moroccan Bay, off W Tangier, Medit. (Alboran Sea, Castellon, NW Basin, Ligurian Sea, Tyrrhenian Sea, Strait of Messina, S Adriatic Sea, Ionian Sea, W Egyptian coast, E Basin), Red Sea, G. of Aden, Argentina, off Portugal, Caribbean, Sargasso Sea, off Bermuda: Station S (32°10N, 64°30W), off SE Cape Cod, G. of Maine, Bay of Fundy, off SE Nova Scotia, off Newfoundland, Ungava Bay, S Strait of Davis, Fram Strait, SE Greenland, Iceland (S & W), Faroe Is., Ireland, North Sea, Norwegian Sea, Malangen fjord, Barents Sea, Wyville Thomson Ridge (60°N), Nordvestbanken, Nansen Basin, E Beaufort Sea, Bering Sea, Aleutian Is., Indonesia-Malaysia, Flores Sea, China Seas (Yellow Sea, East China Sea, South China Sea), Taiwan Strait, Taiwan (S, SW, NE Mienhua Canyon), Kuroshio Current, S Korea, japan (Kuchinoerabu Is., Tanabe Bay), off S Shikoku Is., ? Pacif. (equatorial central), Oregon, California, W Baja California, off W Mexico, W Costa Rica, Pacif. (W equatorial), New Zealand (Kaikoura, NE & SW South Island ), Peru, Chile (N & S), ? Straits of Magellan (Pacific area) | | | | N: | 211 | | | | Lg.: | | | (35) F: 4,7-4,2; M: 3,24; (38) F: 3,8-3,5; M: 3,4-3; (45) F: 4,5-4; M: 3,5-3; (54) M: 2,46; (65) F: 4,3; M: 3,5; (75) F: 3,87-3,42; M: 3,19-2,78; (91) F: 4-3; (116) F: 3,69-2,9; M: 3,3; 3; (199) F: 3,5-2,89; M: 3,34-2,89; (246) F: 4,64-3,91; M: 3,48-3,19; (290) F: 2,15-2,95; M: 1,75-2; (432) F: 4,32-4,1; (449) F: 4,3-3; M: 4-3; (772) F: 4,1-2,7; M: 3,8-2,8; (909) F: 3-4,5; M: 3,1-3,9; (991) F: 3-4,7; M: 3-4; (1023) F: 2,3-2,47; M: 2,1-3,06; (1089) F: 3,5-4; M: 3-3,4; (1122) F: 3,6; M: 3,3; (1230) F: 3,0-4,9; M: 2,6-4,0; {F: 2,15-4,90; M: 2,10-4,00}
The mean female size is 3.661 mm (n = 33; SD = 0.7251), and the mean male size is 3.116 mm (n = 30; SD = 0.5350). The size ratio (male : female) is 0.88 (n = 16; SD = 0.1039). | | | | Rem.: | méso-bathypélagique.
Sampling depth (Antarct., sub-Antarct.) : 200-1000 m. 0-197 m at Station S 2 in S Bösö (E middle Japan).
Les dimensions données (290) par Chen & Zhang (1965) sont exceptionnellement minimales.
D'après C. Séret (1979, p.141) l'espèce effectue des migrations verticales importantes entre la surface et 1500 m. Elle est absente de la surface le jour et se concentre essentiellement vers 600 m. La nuit, son effectif est abondant en surface.
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 09 juin 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
