|
|
 |
Fiche d'espèce de Copépode |
|
|
Calanoida ( Ordre ) |
|
|
|
Metridinidae ( Famille ) |
|
|
|
Pleuromamma ( Genre ) |
|
|
| |
Pleuromamma xiphias (Giesbrecht, 1889) (F,M) | |
| | | | | | | Syn.: | Pleuromma xiphias Giesbrecht,1889;1892 (p.347, 357, 774, figs.F,M) | | | | Ref.: | | | Giesbrecht & Schmeil, 1898 (p.109, Rem. F,M); Thompson & Scott, 1903 (p.235, 249); Esterly, 1905 (p.176, figs.F); Farran, 1908 b (p.61); A. Scott, 1909 (p.124, Rem.); Wolfenden, 1911 (p.289); Sars, 1925 (p.202, figs.F,M); Farran, 1926 (p.273); 1929 (p.209, 260); Rose, 1929 (p.30); Farran, 1929 (p.260); Sciacchitano, 1930 (p.26, figs.F,M); Steuer, 1932 a (p.5, 70, figs.F,M); Sewell, 1932 (p.269, fig.M); Wilson, 1932 a (p.figs.F); Rose, 1933 a (p.181, figs.F,M); Farran, 1936 a (p.111); Mori, 1937 (1964) (p.70, figs.F,M); Jespersen, 1940 (p.47); Lysholm & al., 1945 (p.32); Sewell, 1947 (p.169, fig.44, figs.M, Rem.); Farran, 1948 f (n°17, p.3, figs.F); Davis, 1949 (p.51, F,M); Brodsky, 1950 (1967) (p.307, figs.F,M); Marques, 1953 (p.110, fig.M); Vervoort, 1957 (p.124, Rem.); Chiba & al., 1957 (p.308); 1957 a (p.12); Grice, 1962 (p.215, figs.F,M, Rem.); Tanaka, 1963 (p.23, Rem.F,M); Giron-Reguer, 1963 (p.31, fig.F); Paiva, 1963 (p.53); Chen & Zhang, 1965 (p.69, figs.F); Vervoort, 1965 (p.105, Rem.); Owre & Foyo, 1967 (p.74, figs.F,M); Ramirez, 1969 (p.69, figs.F,M, Rem.); Shih & al., 1971 (p.42); Bradford, 1972 (p.46, figs.M); Arcos, 1975 (p.20, figs.M); Dawson & Knatz, 1980 (p.5, figs.F,M); Björnberg & al., 1981 (p.640, figs.F,M); Gardner & Szabo, 1982 (p.344, figs.F, M); Zheng & al., 1982 (p.46, figs.M); Ferrari, 1984 a (p.168, 170, fig.M); 1985 (p.2 & suiv., figs.F,M, juv., Rem.); Baessa de Aguiar, 1986 (1989) (p.60, Rem.); Huys & Boxshall, 1991 (p.65, figs.); J.S. Park & Mauchline, 1994 (p.108 & suiv., fig.F); Weatherby & al., 1994 (p.670 & suiv.); J.S. Park, 1995 (p.212, figs.); Lenz & al., 1996 (p.355, figs. A1); Chihara & Murano, 1997 (p.839, tab.7, Pl.130: F,M); Cuoc & al., 1997 (p.656, figs.F); Ferrari & Benforado, 1998 a (p.209, figs. juv., F,M); Bradford-Grieve & al., 1999 (p.884, 948, figs.F,M); Bradford-Grieve,1999 b (p.124, figs.F,M, Rem., figs.179, 192); Conway & al., 2003 (p.96, figs.F,M, Rem.); G. Harding, 2004 (p.29, figs.F,M); Boxshall & Halsey, 2004 (p.147: fig.F); Mulyadi, 2004 (p.66, figs.F,M, Rem.); Ferrari & Dahms, 2007 (p.63, 68, 72, 87, Rem., Table V, X); Vives & Shmeleva, 2007 (p.384, figs.F,M, Rem.); Goetze, 2011 (p.580, genetic structure); Takenaka & al., 2013 (p.201, DNA sequences vs luciferase) | 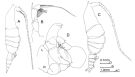 isued from : J.M. Bradford-Grieve in The Marine Fauna of New Zealand: Pelagic Calanoid Copepoda. National Institute of Water and Atmospheric Research (NIWA). NIWA Biodiversity Memoir, 111, 1999. [p.126, Fig.86]. Female (41°53'S, 174°55'E): A, habitus (right lateral side); B, P5. Male: C, habitus (right lateral side); D, P5. Female characteristics: Resembles P. abdominalis but differs in outline of anterior head which continues forwards into a pointed process which is bent slightly downwards. Male characteristics: Forehead as in female except it is directed forwards not downwards
|
 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.46, Fig.14]. Female (from N Cape Verde Is.): A, habitus (lateral right side); B, urosome (ventral); C, genital segment (lateral right side); D, caudal rami (dorsal); E, caudal ramus (ventral). Bars: A = x; B-E = Y. Nota: Caudal rami with dorsally row of hairs and 2 pores, ventrally 1 large pore near insertion of lateral seta; 4 distal pores, 4 apical, 1 dorsal, 1 lateral setae
|
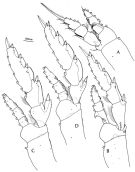 issued from : F.D. Ferrari in Smiths. Contr. Zool., 1985, 420. [p.47, Fig.15]. Female: A, P1; B, P2; C, P3; D, P4.
|
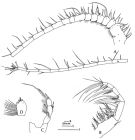 issued from : F.D. Ferrari in Smiths. Contr. Zool., 1985, 420. [p.48, Fig.16]. Female: A, A1; B, Mxp; C, P5; D, terminal segment of P5. Bars: A, B on scale R; C on scale S; D on scale T.
|
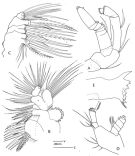 issued from : F.D. Ferrari in Smiths. Contr. Zool., 1985, 420. [p.49, Fig.17]. Female: A, A2; B, Mx1; C, Mx2; D, Md (mandibular palp); E, Md (mandibular blade). Nota: Mx2 bears 6 inner lobes (although Mx2 of calanoids is generally considered to have no more than 5 inner lobes plus an endopod), but the last lobe may be obscured by lobe 5 and the developed endopod.
|
 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.50, Fig.18]. Male: A, habitus (lateral left side); B, urosome (dorsal); C, idem (ventral); D, rostrum (lateral); E, P5 (anterior); F, idem (posterior). A on scale x; B, C on scale Y; D-F on scale Z.
|
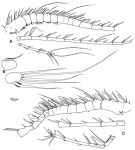 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.51, Fig.19]. Male: A, left A1; B, caudal rami (ventral); C, right A1. Nota: Caudal rami asymmetrical in structure and lateral spine.
|
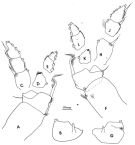 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.52, Fig.20]. Male: P1: left basipodal segments 1 and 2 (anterior); B, left basipodal segment 2 (posterior); C, left exopodal segments 1-3 (anterior); D, left endopodal segment 1 (anterior); E, left endopodal segments 2-3 (anterior); F, right basipodal segments 1-2 (anterior); G, right basipodal segment 2 (posterior); H, right endopodal segment 1 (anterior); J, right exopodal segments 2-3 (anterior); K, right endopodal segment 1 (anterior); L, right endopodal segments 2-3 (anterior).
|
 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.53, Fig.21]. Male: P2: A, left basipodal segments 1-2 (anterior); B, left exopodal segment 1 (anterior); C, left exopodal segment 2 (anterior); D, left exopodal segment 3 (anterior); E, idem (lateral); F, left endopodal segment 1 (anterior); G, idem (lateral); H, left endopodal segment 1 (posterior); J, left exopodal segment 2 (anterior); K, left endopodal segment 3 (anterior); L, right basipodal segments 1-2 (anterior); M, right exopodal segment 1 (anterior); N, right exopodal segment 2 (anterior); P, right exopodal segment 3 (anterior); Q, idem (lateral); R, right endopodal segment 1 (anterior); S, idem (lateral); T, idem (posterior); U, right endopodal segment 2 (anterior); V, right endopodal segment 3 (anterior).
|
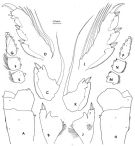 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.54, Fig.22]. Male: P3: A, left basipod segments 1-2; B, left exopodal segment 1; C, left exopodal segment 2; D, left exopodal segment 3; E, left endopodal segment 1; F, left endopodal segment 2; G, left exopodal segment 3; H, right basipodal segments 1-2; J, right exopodal segment 1; K, right exopodal segment 2; L, right exopodal segment 3; M, right endopodal segment 1; N, right endopodal segment 2; P, right endopodal segment 3.
|
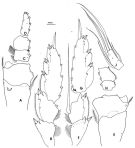 issued from : F.D. Ferrari in Smiths; Contr. Zool., 1985, 420. [p.55, Fig.23]. Male: P4: A, left basipodal segmets 1-2; B, left exopodal segments1-3; C, left endopodal segments 1-2; D, left endopodal segment 3; E, right basipodal segments 1-2; F, right exopodal segments 1-2; G, right exopodal segment 3; H, right endopodal segments 1-2; J, right endopodal segment 3.
|
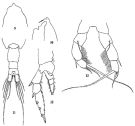 issued from: Q.-c Chen & S.-z. Zhang in Studia Marina Sinica, 1965, 7. [Pl.22, 9-13]. Female (from E China Sea): 9, forehead (dorsal); 10, idem (lateral); 11, urosome (dorsal); 12, left P2 (posterior); 13, P5 (posterior).
|
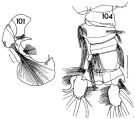 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.70, Lam. XIII, figs.101, 104]. Male (from off Mar del Plata): 101, left P5; 104, urosome (dorsal). Scale bars in mm: 0.05 (101); 0.2 (104).
|
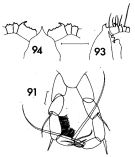 issued from : F.C. Ramirez in Contr. Inst. Biol. mar., Buenos Aires, 1969, 98. [p.64, Lam. XII, figs.91, 93, 94 ]. Female (from off Mar del Plata): 91, P5; 93, forehead (lateral); 94, idem (dorsal). Scale bars in mm: 0.05 (91); 0.5 (93, 94).
|
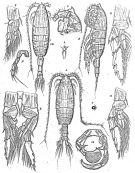 Issued from : G.O. Sars in Résult. Camp. Scient. Prince Albert I, 69, pls.1-127 (1924). [Pl.LV, figs.1-12]. Female: 1, habitus (dorsal); 2, idem (lateral left side); 3, forehead (lateral); 4, rostrum; 5, P5. Male: 6, habitus (dorsal); 7, luminous organ on lateral left side of the posterior margin of the head; 8, P1; 9, P2; 10, P3; 11, P4; 12, P5.
|
 issued from : R.B.S. Sewell in The John Murray Expedition, 1933-34, Scientific Reports, VIII (1), 1947. [p.170, Fig.45]. Male (from Arabian Sea): A, habitus (dorsal); B, urosome (lateral left side) of forma minor; C, idem of forma major; D, A1 (hinge joint) of forma minor; E, idem of forma major; F, right P2 of forma major; G, idem of forma minor; H, exopod of P3 of forma major; I, P5 of forma major; J, P5 of forma minor.
|
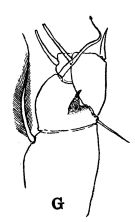 issued from : R.B.S. Sewell in The John Murray Expedition, 1933-34, Scientific Reports, VIII (1), 1947. [p.166, Fig.44, G]. Process on the posterior aspect of the 2nd basal segment of P1. Remarks: The shape of the hook-like or spine-like process shows some variation in different genera, but it is undoubtedly homologous throughout the whole series. The function of this organ is unknown.
|
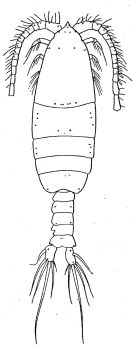 Issued from : R.B.S. Sewell in Mem. Indian Mus., 1932, X (continued). [p.270, Fig.91]. Male (from Sri lanka): habitus, dorsal (arrangement of the cutaneous pores).
|
 issued from : F. Giron-Reguer in Thèse Fac. Sc. Paris, 1963 [Fig.10]. Female (from Alboran Sea): forehead (dorsal).
|
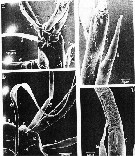
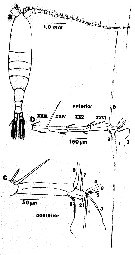 issued from : T.N. Weatherby, K.K. Wong & P.H. Lenz in J. Crustacean Biol., 1994, 14 (4). [p.672, Fig.1]. Line drawing of P. xiphias and right A1. a, dorsal view of female showing relative length and normal position of antenna as seen in live animals; b, dorsal view of distal segment (XXIII to XXVII-XXVIII) and setae of first antenna of male, showing normal position of setae as seen in live animals. Segment nomenclature follows Huys & Boxshall (1991). Numbering of setae used throughout this text: c, highter magnification drawing of (b) showing two most distal segments. The morphological results indicate the presence of 9 identifiable setae on the distal tip of A1; these include 5 dually innervated and 2 singly innervated mechanoreceptive setae. The setae of the distal tip as a single sensory element reflect in physiological and behavorial specialization. Seta number 1 is the only long posteriorly pointing seta. Seta number 2 clearly has an associated function, being closely apposed to number 1 and, secondarily shortened (indicating that its original length has become superfluous). Seta number 3 is unlike any other seta in its remarkable flattened whiplike shape, the function of which is still obscure. Seta 4 is the only chemosensilla within 0.200 mm of the tip, and we have as yet no information as to its chemical sensitivity. Seta 5 is the only pure mechanosensory seta of a length which seems to be a part of the sequence of declining lengths running out the antennular shaft and is one of only three singly innervated setae (numbers 6 and 7 being the other two). Seta number 6 appears vestigial and is probably completely within the boundary layer for most hydrodynamic stimuli. Seta 7 appears to be only bimodal mechano-chemosensory seta on the tip. Seta number 8 is the second longest seta, the longest anteriorly pointing one on the antenna and the only one which appears to be controlled by a discrete muscle. Seta number 9 is the only \\\\\\\"feathered\\\\\\\" seta on the tip, sharing the feathered attribute with only two other setae in the distal region (segments XXV and XXIV), which may suit them well for monitoring direction of water flow.
|
 issued from : T.N. Weatherby, K.K. Wong & P.H. Lenz in J. Crustacean Biol., 1994, 14 (4). [p.673, Fig.2]. Segment XXVII-XXVIII female: right A1: a, dorsal view; insertion of setae numbers 1 and 2 into single socket is shown. b, ventral view: seta number 3 and its insertion into antennal shaft are visible (arrow)), as is ventral pit pore (P). c, close-up of cuticular depression in seta number 1 (arrow), which appears to accomodate seta number 2 (dorsal view); beyond setae numbers 1 and 2 textured loop of seta number 3 is visible. d, higher magnification of setae numbers 1 and 2; note difference in cuticular texture between basal and distal regions of both setae. r = ridge of setal socket; S = fold separating posterior and anterior setae; P = pore.
|
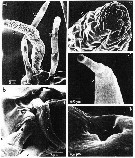 issued from : T.N. Weatherby, K.K. Wong & P.H. Lenz in J. Crustacean Biol., 1994, 14 (4). [p.675, Fig.3]. Segment XXVII-XXVIII of A1 male. a, dorsal view of 4 anterior setae (left , non-geniculated A1). b, end-on view of right antennal tip showing ventral surface (at top); shown are ventral pit pore, ridge of socket of setae numbers 1 and 2, and common base shared by setae numbers 4 and 5. c, apical invagination on seta number 4 (left A1). d, apical pore on seta number 7 (left A1). e, higher magnification of ventral pit pore. P = pit pore; r = raised ridge of setal socket; S = fold separating posterior and anterior setae.
|
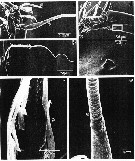 issued from : T.N. Weatherby, K.K. Wong & P.H. Lenz in J. Crustacean Biol., 1994, 14 (4). [p.677, Fig.4]. Segment XXVI of A1. a, dorsal view of seta number 8 (male, left, non-geniculated A1). b, dorsal view of seta number 9 (male, right A1, geniculated). c, higher magnification of seta number 9; note insertion of setules and laterally flattened pseudoannulated appearance of setal shaft. d, top: ventral view of distal tip (female, right A1) showing position of pore on XXVI. Bottom: magnified view of pore. e, higher magnification of seta number 8, showing transition from smooth basal to pseudoannulated region.
|
 issued from : T.N. Weatherby, K.K. Wong & P.H. Lenz in J. Crustacean Biol., 1994, 14 (4). [p.678, Fig.5]. Basic structure of mechanoreceptors in distal tip of A1. a, dorsal view of distal subsegments with schematic drawing of longitudinal section of sensory and scolopale cell structures for seta number 8. Bipolar sensory cell bodies (scb) are located in segment XXVI, extending their proximal dendites (pd) distally. A peridentritic accessory cell containing scolopale material (sc) forms tube, enclosing distal dendrite(s) (dd) in liquor-filled cavity. Scolopale is elaborated from ciliary (basal body, bb) region of dendrite nearly to point at which seta segments antennal shaft. Bands of microtubules (mtb) appear at intervals, anchoring scolopale cell to cuticle. Only one distal dendrite shown for clarity. Dashed lines labeled 6a and 6b correspond to antennal cross sections shown in Fig.6. b-d, cross sections through distal dendrites of mechanoreceptive sensory structures. Location of these cross sections shown by corresponding letter on left of antennular shaft in a. magnifications of micrographs and expanded views as shown in (d). b, pair of distal dendrites (dd) containing arrays of microtubules having both electron-dense and electron-lucent centers. Pair of dendrites loosely wrapped by thin processes of 2 or 3 accessory cells. Expanded view shows magnification of microtubules ; c, distal dendrites within scolopale tube (sc) show proliferation of microtubules with electron-dense cores. Microtubules arise in rows from 9 electon-dense crescents in each dendrite. Expanded view shows magnification of crescents and microtubules. d, ciliary region of sensory structure, showing pair of dendrites running through liquor cavity (lc), enclosed by thin scolopale tube. Left dendrite sectioned at level of basal body ; other just distal to its basal body, showing 9 + 0 circlet of microtubules with electron-dense centers in distal dendrite (see expanded view). Desmosomes formed between scolopale cell and attachment cell (*) mark sites of attachment of bands of microtubules which extend ti cuticle. Nota: The receptor organs are characterized by well)developed scolopale tubes and large numbers of microtubules within the dendrites. The scolopale, which acts as a supporting structure, forms a complete tube around the liquor cavity and dendrites.
|
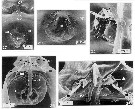 issued from : C. Cuoc, D. Defaye, M. Brunet, R. Notonier & J. Mazza in Mar. Biol., 1997, 129. [p.657, Figs.27, 29, 32, 34, 36]. Scanning electron micrographs of genital area of females (from N Atlantic). 27, outer genital area of inseminated female; note pedunculate structure (arrow) on spermatophoral plug ( sp); 29, epicuticular membrane (star) masking the copulatory pore ( ) of virgin female; 32, inner dorsal view of left egg-laying duct (d3); 34, inner dorsal view of genital area; note position of seminal (d1) and blind () ducts; 36, inner dorsal view of parts of genital area; note single shell duct (d2; bd: blind duct.
m: muscle.
|
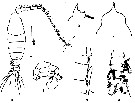 issued from : Z. Zheng, S. Li, S.J. Li & B. Chen in Marine planktonic copepods in Chinese waters. Shanghai Sc. Techn. Press, 1982 [p.46, Fig.26]. Male: a, habitus (dorsal); b, forehead (lateral); c, idem (dorsal view); d, urosome (ventral); e, geniculate segments of left A1; f, P5. Scale bar in mm.
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.36, Figs.1-2, 5]. Female: 1, habitus (dorsal); 2, P5; 5, forehead (lateral).
|
 issued from : T. Mori in The pelagic Copepoda from the neighbouring waters of Japan, 1937 (1964). [Pl.36, Figs.3-4]. Male: 3, habitus (dorsal); 4, P5 (posterior).
|
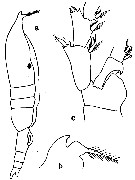 issued from : C.O. Esterly in Univ. Calif. Publs. Zool., 1905, 2 (4). [p.176, Fig.34]. Female from San Dego Region): a, habitus (lateral; b, forehead (lateral); c, 2nd basal segment and proximal segments of rami of P2.
|
 issued from : J.M. Bradford in Mem. N. Z. Oceonogr. Inst., 1972, 54. [p.45, Fig.11, (8-9)]. Male (from Kaikoura, New Zealand): 8, habitus (dorsal); 9, P5. Scale bars: 1 mm (8); 0.1 mm (9).
|
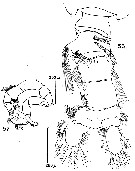 issued from : D.F.R. Arcos in Gayana, Zool., 1975, 32. [Lam.VI, Figs.56-57]. Male (from Bahia de Concepcion, Chile): 56, urosome (dorsal); 57, P5.
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.214, Pl.20, Figs.18-21]. Female (from equatorial Pacific): 18-19, habitus (dorsal and lateral, respectively); 20, posterior part of thorax and urosome (dorsal); 21, same (lateral, left side).
|
 issued from : G.D. Grice in Fish. Bull. Fish and Wildl. Ser., 1962, 61. [p.216, Pl.21, Figs.1-5]. Female (from equatorial Pacific): 1, proximal part of A1; 2, P5. Male: 3, habitus (lateral, right side); 4, urosome (dorsal); 5, P5.
|
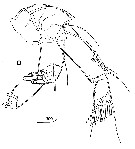 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.65, Fig.2.2.12, D]. From Sargasso Sea: D, A2.
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.71, Fig.2.2.18, A-C]. A, Mx1; B, same (exopodal setae); C, same (endopod).
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.72, Fig.2.2.19, A]. A, armature of maxillulary arthrite.
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.74, Fig.2.2.21, A-D]. A, Mx2 (complete limb); B, basis and endopod with 1st endopodal segment shaded (anterior view); C, basis and endopod (posterior view); D, endopod with segments disarticulated).
|
 issued from : R. Huys & G.A. Boxshall in Copepod Evolution. The Ray Society, 1991, 159. [p.76, Fig.2.2.23, A]. A, Mxp.
|
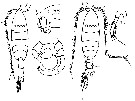 issued from : G. Harding in Key to the adullt pelagic calanoid copepods found over the continental shelf of the Canadian Atlantic coast. Bedford Inst. Oceanogr., Dartmouth, Nova Scotia, 2004. [p.29]. Female & Male. Nota: Forehead produced anteriorly into sharp point in both sexes (arrowed) and A1 with strong curved spine on segment 1 and straight spine on segment 2..
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.33, Fig.45]. Female: 45, forehead (lateral).
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.32, Fig.14]. Male: 14, right A1 (distal segments 14 to 25). Ae = aesthetasc; Spr = plumose seta; Sdi = distal seta.
|
 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.33, Fig.42]. Male: 42, forehead (lateral).
|
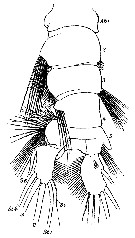 Issued from : W. Giesbrecht in Systematik und Faunistik der Pelagischen Copepoden des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. Fauna Flora Golf. Neapel, 1892, 19 , Atlas von 54 Tafeln. [Taf.33, Fig.50]. Male: 50, urosome (dorsal).
|
 issued from : Mulyadi in Published by Res. Center Biol., Indonesia Inst. Sci. Bogor, 2004. [p.66, Fig.37]. Female (from Flores Sea): a, forehead (dorsal); b, urosome (dorsal); ; c, P5. Male: dhabitus (dorsal); e, P5.
|
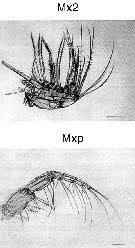 Issued from : M. Sano, K. Maki, Y. Nishibe, T. Nagata & S. Nishida in Progr. Oceanogr., 2013, 110. [p.19, Figs. 8 d, 9, d]. Second maxilla (Mx2) and maxilliped (Mxp) of Pleuromamma xiphias from Sagami Bay (Japan) in April 2009. Scale bars: 200 µm. Compare appendage structure and setal armement with Euchirella rostrata, Undeuchaeta major, Chirundina streetsii, Scaphocalanus echinatus, Scottocalanus helenae, Scottocalanus securifrons, Spinocalanus magnus and calanus sinicus for interpreted particles catching.
|
 Issued from : P.H. Lenz, T.M. Weatherby, W. Weber & K.K. Wong inZooplankton sensory ecology and p)hysiology. Edit. P.H. Lenz, D.K. Hartline, J.E. Purcell & D.L. Macmillan, 1996 . [p.357, Fig. 1A]. Drawing of segments 6 to 10 (ancestral segments VIII to XIII) showing distribution of setae and pattern of methylene blus staining of neurone (dense stippling). Azocarmine-staining aesthetasc-like setae indicated by light stippling. Right A1 of adult female Pleuromamma xiphias.
|
 Issued from : P.H. Lenz, T.M. Weatherby, W. Weber & K.K. Wong inZooplankton sensory ecology and p)hysiology. Edit. P.H. Lenz, D.K. Hartline, J.E. Purcell & D.L. Macmillan, 1996 . [p.357, Fig. 1B]. Scanning electron micrograph of apical pore of a tapered seta. Proximal seta located on segment 1 (ancestral segment III) on the left A1 in an adult male P. xiphias.
|
 Issued from : P.H. Lenz, T.M. Weatherby, W. Weber & K.K. Wong inZooplankton sensory ecology and p)hysiology. Edit. P.H. Lenz, D.K. Hartline, J.E. Purcell & D.L. Macmillan, 1996 . [p.360, Table 1]. Distribution of identified sensory setae on A1 of P. xiphias female. Nota: Table 1 shows a segmental account of all setal types on A1 females: of the 80 identified setae on A1, 60% are mixed-modality, possessing a distal pore and innervated by both mechano- and putative chemosensory neurons. Aesthetasc-like chemosensory setae account for 29% of the total. Pure mechanosensory setae showing no evidence of chemosensory dendrites within the setae and lacking a distal pore, account for 11% of the setae. The latter are primarily found distally and they include hree posteriorly-pointing feathered setae.
|
 issued from : J. Yen, P.H. Lenz, D.V. Gassie & D.K. Hartline in J. Plankton Res., 1992, 14 (4). [p.502, Fig. 4]. Neural activity from the A1 of Pleuromamma xiphias from Kaneohe Bay, Hawaii. Left side: spontaneous background activity. Right side: responses to a variety of mechanical stimuli (bending seta). All recordings are 40 ms long. Compare with Labidocera madurae, Euchirella curticauda, Undinula vulgaris, Acartia fossae.
| | | | | Ref. compl.: | | | Cleve, 1904 a (p.195); Hardy & Gunther, 1935 (1936) (p.167, Rem.); Wilson, 1942 a (p.202); Oliveira, 1945 (p.191); Sewell, 1948 (p.329, 503, 514, 519, 525, 532, 534, 539, 547, 549, 558, 567); Moore, 1949 (p.54); C.B. Wilson, 1950 (p.290); King & Hida, 1955 (p.11); Yamazi, 1958 (p.150, Rem.); Wickstead, 1962 (p.546, food & feeding); Fagetti, 1962 (p.28); Giron-Reguer, 1963 (p.31); Grice, 1963 a (p.496); Ahlstrom & Thrailkill, 1963 (p.57, Table 5, abundance); Gaudy, 1963 (p.25, Rem.); Björnberg, 1963 (p.51, Rem.); Deevey, 1964 (p.589, Table 1, lengths variation); Unterüberbacher, 1964 (p.27); De Decker & Mombeck, 1964 (p.13); Grice & Hulsemann, 1965 (p.224); Pavlova, 1966 (p.44); Mullin, 1966 (p.546, Table I, III, diet); Furuhashi, 1966 a (p.295, vertical distribution in Oyashio/Kuroshio transitional area, Table 8, 9); Mazza, 1966 (p.71); 1967 (p.321, 323, fig.64); Ehrhardt, 1967 (p.740, 887, figs.13, 51-53, geographic distribution, Rem.); Fleminger, 1967 a (tabl.1); Grice & Hulsemann, 1967 (p.17); Delalo, 1968 (p.138); Séguin, 1968 (p.488); De Decker, 1968 (p.45); Vinogradov, 1968 (1970) (p.81, 83, 91, 277); Omori, 1969 (p.5, Table 1); Morris, 1970 (p.2301); Park, 1970 (p.477); Timonin, 1971 (p.281, trophic group); Gueredrat, 1971 (p.300, fig.10, Table 1, 2); Lee & al., 1971 (p.1150, 1151); Deevey, 1971 (p.224); Roe, 1972 (p.277, tabl.1, tabl.2); 1972 b (p.545); Champalbert & Gaudy, 1972 (p.159, respiration vs. temperature); Bainbridge, 1972 (p.61, Appendix Table III: occurrence); Björnberg, 1973 (p.338, 388); Harding, 1974 (p.141, tab. 2, gut contents); Vives & al., 1975 (p.46, tab.II, III, XII); Arashkevich, 1975 (p.351, digestion time); Gaudy, 1975 (p.109, Table I, respiration); Deevey & Brooks, 1977 (p.256, tab.2, Station "S"); Timonin & Voronina, 1977 (p.290); Tranter, 1977 (p.596, 603); Carter, 1977 (1978) (p.36); Omori, 1978 a (p.261, Table 1: chemical composition); Dessier, 1979 (p.89, 206); Hayward, 1980 (p.295, Table 2, vertical distribution, feeding); Vives, 1982 (p.293); Rudyakov, 1982 (p.208, Table 2); Kovalev & Shmeleva, 1982 (p.84); Dessier, 1983 (p.89, Tableau 1, Rem., %); Gaudy & Boucker, 1983 (p.37, Table 1, Rem.: metabolism); Tremblay & Anderson, 1984 (p.5); Guangshan & Honglin, 1984 (p.118, tab.); De Decker, 1984 (p.316, 361: chart); Sameoto, 1984 (p.767, vertical migration); Cummings, 1984 (p.163, Table 2); Petipa & Borichenko, 1985 (tab.1); Longhurst, 1985 (tab.2); Brenning, 1985 a (p.28, Table 2); 1986 (p.8, spatial distribution, Rem.); Wishner & Allison, 1986 (tab.2); Chen Y.-Q., 1986 (p.205, Table 1: abundance %, Table 2: vertical distribution); Saraswathy, 1986 (p.192); Madhupratap & Haridas, 1986 (p.105, tab.1); Rudyakov, 1986 (tab.1); Ambler & Miller, 1987 (tab.2, 3, 4, 5); Latz & al., 1987 (tab.IV); Lozano Soldevilla & al., 1988 (p.59); Buskey & al., 1989 (p.207, vertical distribution vs light penetration); Heinrich, 1990 (p.18); Madhupratap & Haridas, 1990 (p.305, vertical distribution night/day; fig.7: cluster); Timonin, 1990 (p.479); Suarez & al., 1990 (tab.2); Suarez & Gasca, 1991 (tab.2); Hattori, 1991 (tab.1, Appendix); Baessa De Aguiar, 1991 (1993) (p.107); Suarez, 1992 (App.1); Yen & al., 1992 (p.495, tab.1, mechanoreception); Ashjian & Wishner, 1993 (p.483, abundance, species group distributions); Seguin & al., 1993 (p.26, Rem.); Hays & al., 1994 (tab.1); Heinrich, 1995 (tab.1); Webber & Roff, 1995 (tab.1); Shih & Young, 1995 (p.71); Kotani & al., 1996 (tab.2); Lenz & al., 1996 (p.1, behaviour); Errhif & al., 1997 (p.422); Suarez-Morales & Gasca, 1997 (p.1525); Gilabert & Moreno, 1998 (tab.1, 2); Suarez-Morales, 1998 (p.345, Table 1); Suarez-Morales & Gasca, 1998 a (p.110); Mauchline, 1998 (tab.23, 33, 63); Hartline & al., 1999 (p.132, Rem.: biolux); Neumann-Leitao & al., 1999 (p.153, tab.2); Lapernat, 2000 (tabl. 3, 4); Razouls & al., 2000 (p.343, Appendix); Steinberg & al., 2000 (p.137, fig.1: biomass time series 1994-1997, figs.2, 3, 4: respiration/excretion, C flux); Haury & al., 2000 (p.69, Table 1); Lopez-Salgado & al., 2000 (tab.1); Lenz & al., 2000 (p.337); Madhupratap & al., 2001 (p. 1345, vertical distribution vs. O2, figs.4, 5: clusters, p.1353); Rebstock, 2001 (tab.2); Holmes, 2001 (p.20); Sameoto & al., 2002 (p.13); Rebstock, 2002 (p.71, Table 3, 5, 6, Fig.2, climatic variability); Beaugrand & al., 2002 (p.1692); Beaugrand & al., 2002 (p.179, figs.5, 6); Schnetzer & Steinberg, 2002 (p.89, figs.); Hsiao & al., 2004 (p.326, tab.1); Pusch & al., 2004 (251, tab.3); Gallienne & al., 2004 (p.5, tab.3); CPR, 2004 (p.60, fig.180); Lo & al., 2004 (p.89, tab.1); Berasategui & al., 2005 (p.313, fig.2); Prusova & Smith, 2005 (p.75); Zuo & al., 2006 (p.162: tab.1); Dias & Araujo, 2006 (p.57, Rem., chart); Lavaniegos & Jiménez-Pérez, 2006 (tab.2, Rem.); Ikeda & al., 2006 (p.1791, Table 2); Mackas & al., 2006 (L22S07, Table 2); Khelifi-Touhami & al., 2007 (p.327, Table 1); Hwang & al., 2007 (p.24); Dur & al., 2007 (p.197, Table IV); Neumann-Leitao & al., 2008 (p.799: Tab.II, fig.66); Morales-Ramirez & Suarez-Morales, 2008 (p.521); Fernandes, 2008 (p.465, Tabl.2); Ayon & al., 2008 (p.238, Table 4: Peruvian samples); Gaard & al., 2008 (p.59, Table 1, N Mid-Atlantic Ridge); Waggett & Buskey, 2008 (p.111, Table 1); Lan Y.C. & al., 2008 (p.61, Table 1, % vs stations); C.-Y. Lee & al., 2009 (p.151, Tab.2); Galbraith, 2009 (pers. comm.); Park & Ferrari, 2009 (p.143, Table 5, Appendix 1, biogeography); Williamson & McGowan, 2010 (p.273, Table III, Pacific central gyres: N & S); Dias & al., 2010 (p.230, Table 1); Mazzocchi & Di Capua, 2010 (p.426); Medellin-Mora & Navas S., 2010 (p.265, Tab. 2); Hirst & al., 2010 (p.2193, sex ratio); Hsiao S.H. & al., 2011 (p.475, Appendix I), Tutasi & al., 2011 (p.791, Table 2, abundance distribution vs La Niña event); Tseng L.-C. & al., 2011 (p.47, Table 2, occurrences vs mesh sizes); Andersen N.G. & al., 2011 (p.71, Fig.3: abundance); Pillai H.U.K. & al., 2011 (p.239, Table 3, vertical distribution); Hidalgo & al., 2012 (p.134, Table 2); Takenaka & al., 2012 (p.1669, fig.2, 3, Table 1, bioluminescence); Teuber & al., 2013 (p.28, Table 1, respiration rates); Teuber & al., 2013 (p.1, Table 1, 2, 3, fig.5, abundance vs oxygen minimum zone, respiration rates, enzyme activity); in CalCOFI regional list (MDO, Nov. 2013; M. Ohman, comm. pers.); Kobari & al., 2013 (p.78, Table 2); Bode M. & al., 2013 (p.1, Table 1, 3, respiration rate & ETS activity); Hirai & al., 2013 (p.1, Table I, molecular marker); Peijnenburg & Goetze, 2013 (p.2765, genetic data); Schukat & al., 2013 (p.1, Table 1, 2, fig.2, respiration, ingestion); Sano & al., 2013 (p.11, Table 2, 5, A1, figs.8, 9, feeding habits, vertical distribution, Rem.: p.23); Lidvanov & al., 2013 (p.290, Table 2, % composition); Hwang & al., 2014 (p.43, Appendix A: seasonal abundance); Zaafa & al., 2014 (p.67, Table I : as sciphias, lapsus calami, occurrence); Bonecker & a., 2014 (p.445, Table II: frequency, horizontal & vertical distributions); Fierro Gonzalvez, 2014 (p.1, Tab. 3, 5, occurrence, abundance); Bode & al., 2015 (p.268, Table 1, 2, figs.3, 4, chemical components, trophic level, geographic zone); Dias & al., 2015 (p.483, Table 2, abundance, biomass, production); Zakaria & al., 2016 (p.1, Table 1) ; Benedetti & al., 2016 (p.159, Table I, fig.1, functional characters); El Arraj & al., 2017 (p.272, table 2, spatial distribution); Belmonte, 2018 (p.273, Table I: Italian zones); Bode & al., 2018 (p.840, Table 1, respiration & ingestion rates, depth); Acha & al., 2020 (p.1, Table 3: occurrence % vs ecoregions, Table 5: indicator ecoregions); Hirai & al., 2020 (p.1, Fig. 5: cluster analysis (OTU), spatial distribution). | | | | NZ: | 24 | | |
|
Carte de distribution de Pleuromamma xiphias par zones géographiques
|
| | | | | | | | | | | | | | | | | |  issued from : M. Saraswathy in Mahasagar-Bull. Nat. Inst. Oceanogr., 1986, 19 (3). [p.190, Fig.2B]. issued from : M. Saraswathy in Mahasagar-Bull. Nat. Inst. Oceanogr., 1986, 19 (3). [p.190, Fig.2B].
Distribution of P. xiphias in the Indian Ocean (upper water column of 200 m).
Triangle: South West Moonsoon season: April 16 to October 15 (white triangle = negative stations; black = animals collected). Circle: North East Moonsoon season: October 16 to April 15 (white circle = negative stations; black = numbers of ianimals collected).
Nota: Though there appears to be a slight clustering of positive stations in the equatorial region, the species was randomly distributed in the entire area |
 issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.74, Table 31]. issued from : H.B. Owre & M. Foyo in Fauna Caribaea, 1, Crustacea, 1: Copepoda. Copepods of the Florida Current. 1967. [p.74, Table 31].
Vertical distribution of Pleuromamma xiphias at the ''40-Mile station'' in the Florida Current (± 25°35'N, 79°27'W).
SL 53: 18 V 1958; SL 55: 21 VII 1958. A: during midday; B;: during midnight. |
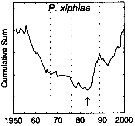 issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 v]. issued from : G.A. Rebstock in Global change Biology, 2002, 8. [p.77, Fig.2 v].
Climatic regime shifts and decadal-scale variability in calanoid copepod populations off southern California (31°-35°N, 117°-122°W.
Cumulative sums of nonseasonal anomalies from the long-term means of copepod abundance from years 1950 to 2000.
A negative slope indicates a period of below-average anomalies; a positive slope indicates a period of above-average anomalies. Abrupt changes in slope indicate step changes. Step changes are marked with arrow (downward -pointing for decreases).
The October 1966 cruise (prior to the increase in sampling depth), March 1976 cruise (prior to the 1976-77 climatic regime shift), and October 1988 cruise (prior to the hypothesized 1989 climatic regime shift) are marked with vertical lines. |
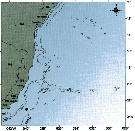 issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.57]. issued from : C. de O. Dias & A.V. Araujo in Atlas Zoopl. reg. central da Zona Econ. exclus. brasileira, S.L. Costa Bonecker (Edit), 2006, Série Livros 21. [p.57].
Chart of occurrence in Brazilian waters (sampling between 22°-23° S).
Nota: sampling only 1 specimen. |
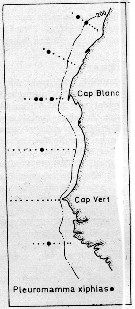 issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.8, Fig.5]. issued from : U. Brenning in Wiss. Z. Wilhelm-Pieck-Univ. Rostock - 35. Jahrgang 1986. Mat.-nat. wiss. Reihe, 5. [p.8, Fig.5].
Spatial distribution for Pleuromamma xiphias from 8° S - 26° N; 16°- 20° W. |
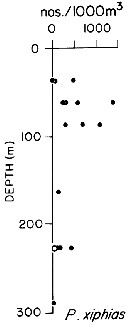 issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.299, Fig.2]. issued from : T.L. Hayward in Mar. Biol., 1980, 58. [p.299, Fig.2].
Plots of day (open circle) and night (filled circle) depth distributions of Pleuromamma xiphias in the North Pacific central gyre (main sampling location: 25°N, 155°W with drogue), September 1968. |
 Issued from : E. Goetze in Integr. Comp. Biol., 2011, 51 (4). [p.587, Fig.3]. Issued from : E. Goetze in Integr. Comp. Biol., 2011, 51 (4). [p.587, Fig.3].
Frequencies of nonsingleton mtCOI haplotypes in population samples of P. xiphias.
Haplotypes are labeled by color, as sampling in the North Atlantic, Pacific and Indian Ocean.
Singleton haplotypes are pooled and labeled in green, black and red for Clades 2, 3 and 4 respectively. White indicates the frequency of H11. Circles indicate population structure as infered from popûlation graph, principal coordinate, and Barrier (see text).
Nota: The species showed high levels of genetic differentiation among samples throughout its global biogeographic range, both within and among ocean basins. Limited dispersal among distinct pelagic provinces seems to have played a major role in population differentiation in this species, with strong genetic breaks observed across known oceanographic fronts or current systems in all three ocean basins.
The Indo-West Pacific (IWP) holds a highly distinct genetic population of this species that was sampled in both the western and eastern Indian Oceans. This suggests that the IWP does not act as a strong barrier to gene flow between basins, despite the relatively shallow water depth (<200 m) and vertically extensive (>400 m) diel migration. A pattern of isolation by distance was observed in the Indian Ocean with genetic differentiation among samples down to spatial scales of ± 800 km, indicating that realized dispersal in P. xiphias occurs over much smaller spatial scales than in previously reported oceanic holoplankton. Given its highly regionalized population genetic structure, P. xiphias may have some capacity to adapt to local oceanographic conditions, and it should not be assumed that populations of this species in distinct pelagic biomes will respond in the same way to shared physical or climatic forcing. |
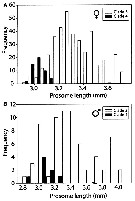 Issued from : E. Goetze in Integr. Comp. Biol., 2011, 51 (4). [p.588, Fig.4]. Issued from : E. Goetze in Integr. Comp. Biol., 2011, 51 (4). [p.588, Fig.4].
Distribution of prosome lengths (mm) for adult females (A) and males (B) of P. xiphias from genetic clades 3 and 4.
Mean body size in clade 4 is signuficantly smaller in both cases (t-test, P <0.001).
Nota: The author interprets the differences in prosome length among mt clades as being ptrimarily environmental in origin, and does not consider these results to be sufficiently strong evidence for distinct species-level status between clades 3 and 4 animals. Comparisons of animals from clades 3 and 4 collected in the same plankton tow, and which presumably experienced the same sea-water temperature during development, show a similar, but non significant, trend toward smaller body size in clade-4 animals (females only). |
 Issued from : M. Sano, K. Maki, Y. Nishibe, T. Nagata & S. Nishida in Progr. Oceanogr., 2013, 110. [p.20, Fig.10]. Issued from : M. Sano, K. Maki, Y. Nishibe, T. Nagata & S. Nishida in Progr. Oceanogr., 2013, 110. [p.20, Fig.10].
Vertical distribution of eight copepod species across 0-1000 m in Sagami bay in 21-27 April 2009.
White and black boxes denote daytime and nighttime distributions. Scaphocalanus echinatus is taken from Kuriyama & Nishida (2006).
Plankton and water samples collected from a fixed station (35°00'N, 139°20'E) using MTD horizontal closing nets (0.33 mm mesh aperture) at 15 depths (0, 50, 95, 100, 150, 195, 200, 295, 300, 395, 400, 500, 600, 800, 1000 m) during both day and night. |
 Issued from : M. Sano, K. Maki, Y. Nishibe, T. Nagata & S. Nishida in Progr. Oceanogr., 2013, 110. [p.23, Table 8]. Issued from : M. Sano, K. Maki, Y. Nishibe, T. Nagata & S. Nishida in Progr. Oceanogr., 2013, 110. [p.23, Table 8].
Previous records on food habits of Pleuromamma xiphias.
After Sano & al. (2013, p.14, Table 2) atomic C:N (mean ± SD) = 7.1 ±0.4, n = 10, in Sagami Bay (21-27 April 2009). |
 Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.13]. Issued from : P.-E. Lapernat in DEA Océanogr. Biol., Univ. P. & M. Curie, Paris VI. July 5, 2000. [Fig.13].
Verical distribution of Pleuromamma xiphias at eutrophic (Mauritania coast: 20°32' N, 18°36' W) (1 from night and 2 from day) and oligotrophic site (off NW Cape Verde Islands: 21° N, 31° W) (3 from night and 4 from day) in females (F) and males (M) (ind. per m3) in the day (white circle) and night (black circle).
Nota: Sampling in the water column 0-1000 m, one during the day and another during the night with BIONESS multiple-net: 0-75; 75-150; 150-250; 250-350; 350-450; 450-550; 550-700; 700-850; 850-965 m. In May-June 1992. |
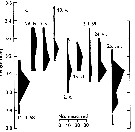 Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.593, Fig. 1, C] Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.593, Fig. 1, C]
Histogram showing the length distribution, plotted as numbers measured of females of Pleuromamma xiphias from 1958-1959, at Station "S' (32°8' N, 64°42' W), 15 miles south-east of Brermuda.
Samples by plankton net, oblique hauls from 500 m to the surface. |
 Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.592, Table 1] Issued from : G. Deevey in J. mar. biol. Ass. U.K., 1964, 44. [p.592, Table 1]
Mean cephalothorax lengths and numbers measured of females and the mean surface temperature and Chlorophyll a of the month preceding each measurement at Station 'S' (SE off Bermuda). |
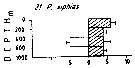 Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4]. Issued from : M. Madhupratap & P. Haridas in J. Plankton Res., 12 (2). [p.311, Fig.4].
Vertical distribution of calanoid copepod (mean +1 SE), abundance No/100 m3. 21- Pleuromamma xiphias.
Night: shaded, day: unshaded.
Samples collected from 6 stations located off Cochin (India), SE Arabian Sea, November 1983, with a Multiple Closing Plankton Net (mesh aperture 300 µm), in vertical hauls at 4 depth intervalls (0-200, 200-400, 400-600, 600-1000 m). |
 Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1). Issued from : M. Bode, R. Koppelmann, L. Teuber, W. Hagen & H. Auel inGlobal Biogeochemical Cycles, 2018, 32. [p.844, Table 1).
Cf. explanations of these measures in Calanoides natalis from the same authors.
Compare with Pleuromamma abdominalis and P. robusta. |
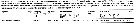 Issued from : M. Bode, A. Schukat, W. Hagen & H. Auel in J. Exp. Mar. Biol. Ecol., 2013, 444. [p.3, Table 1]. Issued from : M. Bode, A. Schukat, W. Hagen & H. Auel in J. Exp. Mar. Biol. Ecol., 2013, 444. [p.3, Table 1].
Dry mass, individual respiration rate and ETS activity from the northern Benguela Current upwelling system along transects at 23°S and 26°40'S, off Walvisbay and Lüderitz (Namibia).
Relationship between individual ETS activities and individual respiration rates; mean ETS activities with standard deviations are plotted again mean respiration rates (Cf. in Calanoides natalis Fig.3). The letter besides each point identifies the associated species in Table 1 (k, for this species) in the same authors.
For Ikeda & al. (2007) the female respiration rate at 3°C was 0.47 µl O2 h ind-1 (DM: 563 µg) in the Western subarctic zone; and 0.55 (DM: 387 µg) at 12°C in unknown region. |
| | | | Loc: | | | Cosmopolite (tropical & temperate): Antarct. (SW Atlant.), South Georgia, sub-Antarct. (Indian, Pacif. SE), South Africa (E & W), Namibia, Angola, G. of Guinea, Argentina (offshore, Brazil (off Rio de Janeiro, off Cabo de Sao Tomé, off Macaé, off Natal), Caribbean Colombia, Caribbean Sea, Jamaica, Yucatan, G. of Mexico, Sargasso Sea, off Bermuda (Station S: 32°10N, 64°30W), off Cape Hatteras, off E Cape Cod, off SE Nova Scotia, S Iceland, Faroe Is., off SW Ireland, North Sea, Great Meteor Seamount, E Azores, Morocco-Mauritania, Canary Is., off W Tangier, Medit. (Alboran Sea, El Kala shelf, Balearic Sea, Ligurian Sea, Tyrrhenian Sea, Strait of Messina, NW Tunisia, Adriatic Sea, W Egyptian coast, E Basin), G. of Aden, Arabian Sea, Indian, off Madagascar S, off SE La Réunion Is., Rodrigues Is.- Seychelles, Natal, Mascarene Basin, S Indian (subtropical convergence), equatorial Indian Ocean, Bay of Bengal, Andaman Sea (Batten Island), W Australia, Indonesia-Malaysia, Flores Sea, China Seas (East China Sea, South China Sea), off SW Taiwan, Taiwan (S, E, N: Mienhua Canyon, NW), Japan, Sagami Bay, Tanabe Bay, Suruga Bay, off Sanriku, S Hokkaido, Kuroshio zone, Pacif. (equatorial central), Hawaii, off NE Hawaii, off British Columbia, California, W Baja California, off W Mexico, W Costa Rica, Pacif. (W equatorial), Pacific (central gyres: N & S), New Zealand (Kaikoura, North Island, ...), Kermadec Is, Tasman Sea, S Tasmania, Pacif. (SE tropical), off W Guatemala, Galapagos-Ecuador, Peru, N Chile off Santiago, Bering Sea | | | | N: | 256 | | | | Lg.: | | | (1) F: 5; (16) F: 4,9-4,2; M: 4,35; (22) F: 5,87-3,5; M: 6,42-4; (26) F: 5,75-4,31; M: 4,45-3,94; (34) F: 4,4-4,08; (35) F: 4,7-4,25; (38) F: 5-4,7; M: 4,7; (45) F: 4,5-4; M: 4,3-4; (47) F: 4,5-4,4; M: 4,3-4; (59) F: 5,8-3,8; M: 6,4-4,1; (73) F: 5,3-5,14; M: 5,82-5,2; (75) M: 5,1; (101) F: 4,25-3,99; M: 4,28-3,99; (116) M: 4,9; (142) F: 4,5-4,1; (199) F: 4,96-4,18; M: 4,96-4,26; (237) F: 3,25-5,0; M: 3,5; (254) F: ± 4,75; M: ± 4,5; (290) F: 4-4,05; (432) F: 4,3-4; (909) F: 4,5-5,1; M: 4,5-5,1; (982) F: 4,56-5,13; M: 4,64-5,59; (991) F: 3,5-5,87; M: 4-6,42; (1023) M: 4,62; (1122) F: 4,05; M: 4,2; {F: 3,25-5,87; M: 3,50-6,42}
The mean female size is 4.530 mm (n = 41; SD = 0.6423), and the mean male size is 4.685 mm (n = 30; SD = 0.7786). The size ratio (male : female) is 1.004 (n = 15; SD = 0.0776). | | | | Rem.: | épi (rare), méso-bathypélagique.
Sampling depth (Antarct., sub-Antarct.) : 0-50-750 m. Les différences de taille observées au NW de lIndien (Sewell, 1947) résulteraient des positions bathymétriques des populations, donc de lorigine des masses deau (eau intermédiaire tropicale et eau intermédiaire sud polaire).
Pleuromamma xiphias f. major
Ref.: Sewell, 1947 (p.169, Rem.)
Loc.: G. dAden, Mer Arabe, Maldives
Lg.: (11) F: 5,379-4,536; M: 5,9-5,017
Pleuromamma xiphias f. minor
Ref.: Sewell, 1947 (p.171)
Loc.: G. dAden, Mer Arabe, Maldives
Lg.: (11) F: 4,52-4,133; M: 4,759-4,235
Voir aussi les remarques en anglais | | | Dernière mise à jour : 28/10/2022 | |
|
|
 Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante : Toute utilisation de ce site pour une publication sera mentionnée avec la référence suivante :
Razouls C., Desreumaux N., Kouwenberg J. et de Bovée F., 2005-2026. - Biodiversité des Copépodes planctoniques marins (morphologie, répartition géographique et données biologiques). Sorbonne Université, CNRS. Disponible sur http://copepodes.obs-banyuls.fr [Accédé le 25 mars 2026] © copyright 2005-2026 Sorbonne Université, CNRS
|
|
|
|




;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)

;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
;)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
